Introduction
Coronary heart disease is a dynamic process in which
there are interactions between endothelial dysfunction and
inflammatory responses (1). Vascular
endothelial cells (VECs), which are pinacocytes located on the
surface of blood vessel lumen, are able to precept hemodynamic and
hematogenous signal changes, and synthesize and secrete various
vaso-active substances (2). VECs
maintain the stability of the internal environment of blood vessels
by releasing vessel dilators and constriction factors (3). If there is an imbalance in these
factors, vasculature may form atheromatous plaques under the
actions of inflammatory factors and cardiovascular risk factors
(4). Inflammatory mediators are the
factors that initiate the occurrence and progression of coronary
heart disease (5).
Janus kinase signal transducer and activator of
transcriptions (JAK/STAT) is a member of the intracellular
transduction signal pathway family (6). As an important signal pathway in this
family, JAK2/STAT3 serves an essential role in coronary heart
disease (7).
As a key factor in signal transduction process,
peroxisome proliferator-activated receptor (PPAR)-γ regulates the
expression of transcription proteins during glycometabolism and
lipogenesis (8). PPAR-γ is able to
regulate lipid metabolism, adipocyte differentiation and insulin
sensitivity (9). A previous study
demonstrated that PPAR-γ is an important cytokine that participates
in glycometabolism and lipid metabolism. It was also found that
PPAR-γ is able to improve lipid levels and regulate xanthoma cell
formations (10), and therefore may
be able to ameliorate atherosclerosis. It has also been
demonstrated that PPAR-γ is able to inhibit atherosclerosis
(10). PPAR-γ is expressed in all
vascular cells and serves an essential role in cell proliferation,
migration, differentiation, the activation of macrophages and
endothelial inflammation (11).
Recent pharmacodynamic studies have demonstrated
that hydroxy safflower yellow A (HSYA) exhibits anti-platelet
aggregation, anti-thrombogenesis, lipid-decreasing,
anti-atherosclerosis, anti-infarction, inhibiting myocardial
apoptosis and anti-oxidative stress functions, and is able to
increase coronary blood-flow volumes, improve micro-circulation and
hemorheology, and promote resistance to anoxia (12,13).
HSYA also has protective actions against acute ischemic myocardium
(12,13). The underlying mechanisms of HSYA are
associated with the regulation of nitric oxide (NO),
6-ketone-prostaglandin F1a, thromboxane B2 and angiotensin II to
increase blood-supply and oxygen supply of myocardium and reduce
myocardial cell injury and apoptosis (13). The aim of the present study was to
investigate the effect of HSYA on coronary heart disease through
assessing the levels of B-cell lymphoma 2 (Bcl-2)/Bcl-2-like
protein 4 (Bax) and PPAR-γ.
Materials and methods
Animals and coronary artery stenosis
model
The present study was approved by the Animal Welfare
and Ethics Committee of Fifth People's Hospital of Chongqing
(Chongqing, China). Male Bama miniature swine (age, 10±2 months;
weight, 23±2 kg, n=30) were obtained from the Animal Center of the
Chongqing Medical University (Chongqing, China) and housed in a
temperature controlled room (22–24°C), 55–60% humidity with a 12-h
dark/cycle and free access to food and water.
Swine were anesthetized with intramuscular ketamine
(35 mg/kg; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and
diazepam (1 mg/kg, Sigma-Aldrich; Merck KGaA) prior to surgery. A
2-cm main operation incision was made at the fourth rib line in the
left axillary level, and a thoracoscope was inserted at the left
sternal 2 level. A 1-cm auxiliary operation incision was made at
the left side of the clavicle midline between the fourth rib line.
Two vessel forceps were subsequently inserted into each hole to
push the lung lobes. Pericardium was separated and cut 2 cm to
acquire the heart. The forward diagonal and ventricular branches of
the left anterior descending (LAD) artery were acquired. The LAD
artery was gently separated into the proximal diagonal branch and
the left main branch, 1 cm apart. The LAD artery was placed on the
operating table at 37°C and the blood vessels were ligated with
silk suture. In sham group, swine were only anesthetized with
intramuscular ketamine (35 mg/kg; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) and diazepam (1 mg/kg, Sigma-Aldrich; Merck
KGaA) prior to surgery.
Experimental animals
A total of 30 Bama miniature swine were randomly
divided into four groups as follows: Sham group (sham; n=6),
coronary heart disease model group (model; n=8), low dose HSYA
group (HSYA-L; n=8) and high dose HSYA group (HSYA-H; n=8). Swine
in the sham and model groups were lavaged with normal saline. Swine
in the HSYA-L and HSYA-H groups were lavaged with 20 or 40 mg/kg of
HSYA, respectively, for 6 h. HSYA was purchased from Shanxi Huahui
Kai Tak Pharmaceutical Co., Ltd. (Shanxi, China) and its chemical
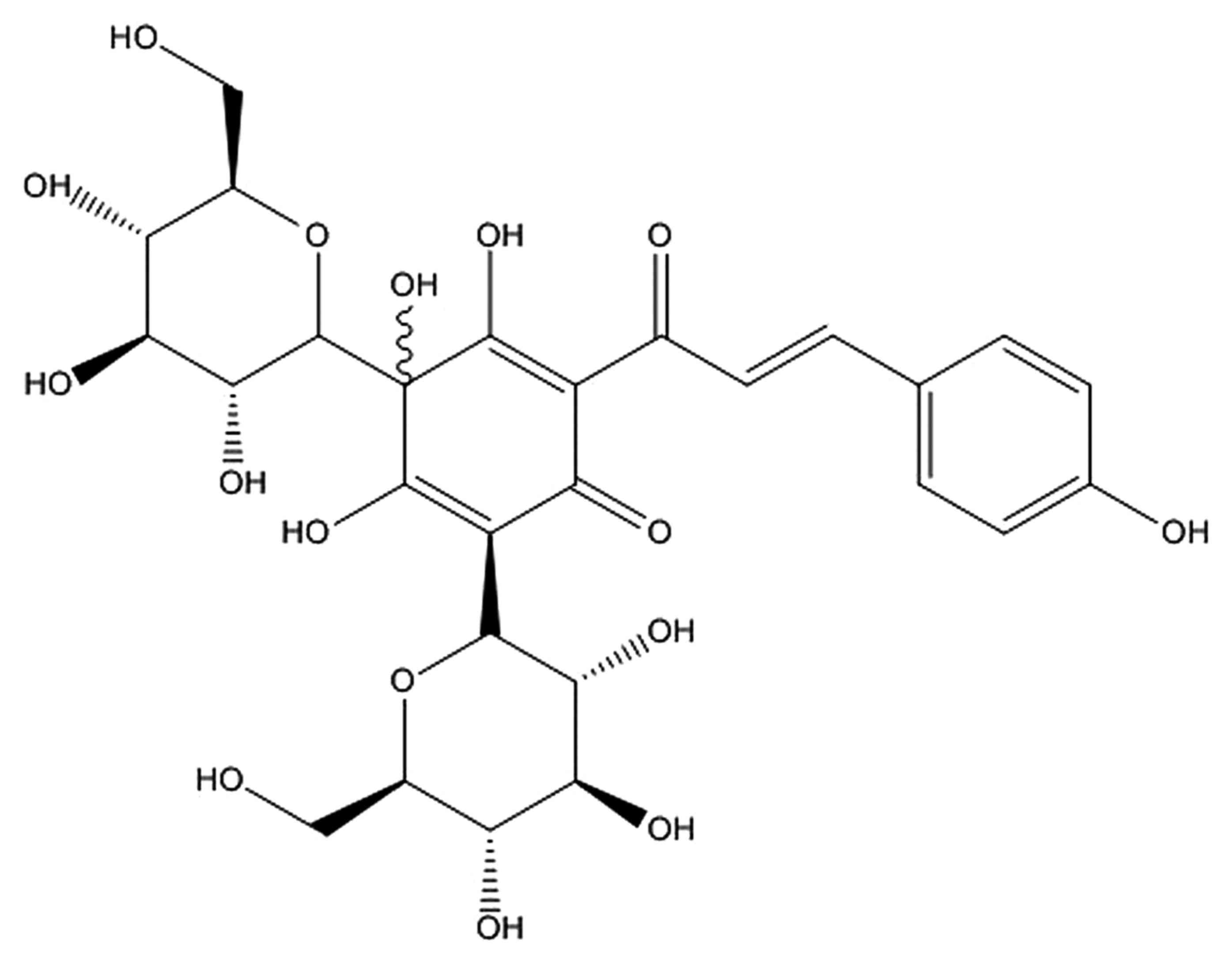
structure is shown in Fig. 1.
Left ventricular ejection fraction
(LVEF), left ventricular systolic internal diameter (LVIDs), blood
urea nitrogen (BUN) plasma creatinine (Scr) and urine levels
Ultrasound (iE33 xMatrix; Philips Healthcare,
Andover, MA, USA) was performed to survey and calculate LVEF and
LVIDs level. The levels of BUN (C013-2), Scr (C011-2) and urine
(C035-2) were measured using ELISA kits (Nanjing Jiancheng Chemical
Industrial Co., Ltd., Nanjing, China).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
After treatment with HSYA, swines were anesthetized
with intramuscular ketamine (35 mg/kg; Sigma-Aldrich; Merck KGaA)
and diazepam (1 mg/kg; Sigma-Aldrich; Merck KGaA), and sacrificed
by intravenous injection with 10% KCl immediately followed blood
emptying. Frozen heart tissue samples was acquired and washed with
PBS. Total RNA was extracted from the frozen heart tissue samples
using TRIzol (Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the manufacturer's protocol. cDNA synthesis was
performed using 2 µg total RNA and Moloney-murine leukemia virus
reverse transcriptase (Invitrogen; Thermo Fisher Scientific, Inc.).
A total of 1 µl cDNA was used for PCR with SYBR Premix ExTaq
(Invitrogen; Thermo Fisher Scientific, Inc.) on an iCycler iQ
Real-time Detection System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The primer sequences for cycloxygenase-2 (COX-2)
and inducible nitric oxide synthase (iNOS) are listed in Table I. PCR amplification was performed as
follows: Initial denaturation at 95°C for 3 min, followed by 40
cycles with denaturation at 95°C for 15 sec and annealing at 60°C
for 30 sec. β-actin was used as a reference gene. Analysis of
relative gene expression data using the 2−ΔΔCq method
(14).
 | Table I.Primers used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used for reverse
transcription-quantitative polymerase chain reaction.
| Primer | Forward sequence | Reverse sequence |
|---|
| COX-2 |
5′-TCGCTGTGCCTGATGATTCC-3′ |
5′-CTTATGATCTCGTTTCCGTC-3′ |
| iNOS |
5′-CCAAGCCCTCACCTACTTCC-3′ |
5′-CTCTGAGGGCTGACACAAGG-3′ |
| β-actin |
5′-CACGAAACTACCTTCAACTCC-3′ |
5′-CATACTCCTGCTTGCTGATC-3′ |
ELISA
After treatment with HSYA, peripheral blood was
collected and serum was collected after centrifugation at 2,000 × g
for 10 min at 4°C. Serum was used to measure IL-1β (H002), IL-6
(H007), IL-10 (H009) and TNF-α (H052) levels using ELISA Kits
(Nanjing institute of biological engineering, Nanjing, China)
according to the manufacturer's instructions.
Western blotting
Heart tissue samples were lysed in ice-cold lysis
buffer (radioimmunoprecipitation assay buffer; Beyotime Institute
of Biotechnology) and protease inhibitor mixture
(phenylmethanesulfonyl fluoride; Beyotime Institute of
Biotechnology) for 30 min. The resulting supernatant was collected
and boiled following 15 min centrifugation at 12,000 × g at 4°C.
Protein concentrations of the extracts were measured by
bicinchoninic acid assay (Nanjing Jiancheng Chemical Industrial
Co., Ltd.). A total of 50 µg whole protein was separated by 8–12%
SDS-PAGE and subsequently transferred onto nitrocellulose
membranes. Membranes were blocked with 5% non-fat milk in TBS and
Tween-20 (TBST) for 1 h at 37°C, and incubated with the following
antibodies: Anti-Bax (sc-6236, 1:3,000; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA), anti-Bcl-2 (sc-783, 1:2,000; Santa Cruz
Biotechnology, Inc.), anti-phosphorylated (p)-JAK2 (3776, 1:4,000;
Cell Signaling Technology, Inc., Danvers, MA, USA), anti-p-STAT3
(9145, 1:2,000; Cell Signaling Technology, Inc.), anti-PPAR-γ
(sc-9000, 1:500; Santa Cruz Biotechnology, Inc.) and β-actin
(sc-7210, 1:1,000) at 4°C overnight, followed by washing with TBST
three times for 10 min. Membranes were subsequently probed with the
appropriate secondary antibodies (sc-2004; 1:5,000; Santa Cruz
Biotechnology) at room temperature for 2 h and washed with TBST.
Protein was and visualized by a BeyoECL Plus (Beyotime Institute of
Biotechnology) and calculated by Image J Image analysis software
3.0 (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
Data were presented as the mean ± standard error of
the mean using SPSS 21.0 (IBM Corp., Armonk, NY, USA). Groups were
evaluated using the Kruskal-Wallis test for nonparametric
independent samples by one-way analysis of variance. P<0.05 was
considered to indicate a statistically significant difference.
Results
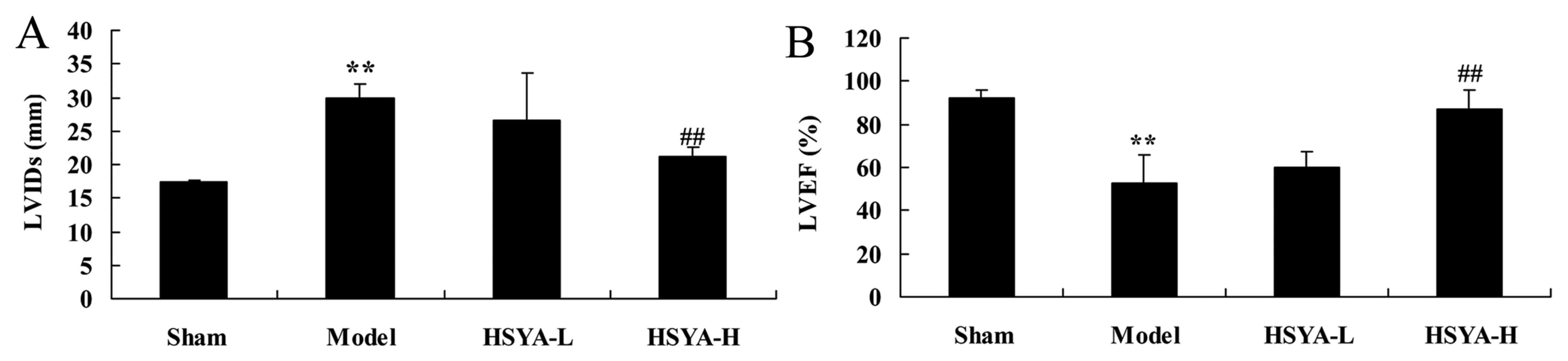
Effect of HSYA on LVIDs and LVEF in
the coronary heart disease model
Compared with the swine in the sham group, the LVIDs
level of the model group was significantly increased (P<0.01;
Fig. 2A). The LVEF were
significantly lower in the model group compared with the shame
group (P<0.01; Fig. 2B).
Treatment with a high dose of HSYA was able to significantly
reverse this effect (P<0.01; Fig 2A
and B).
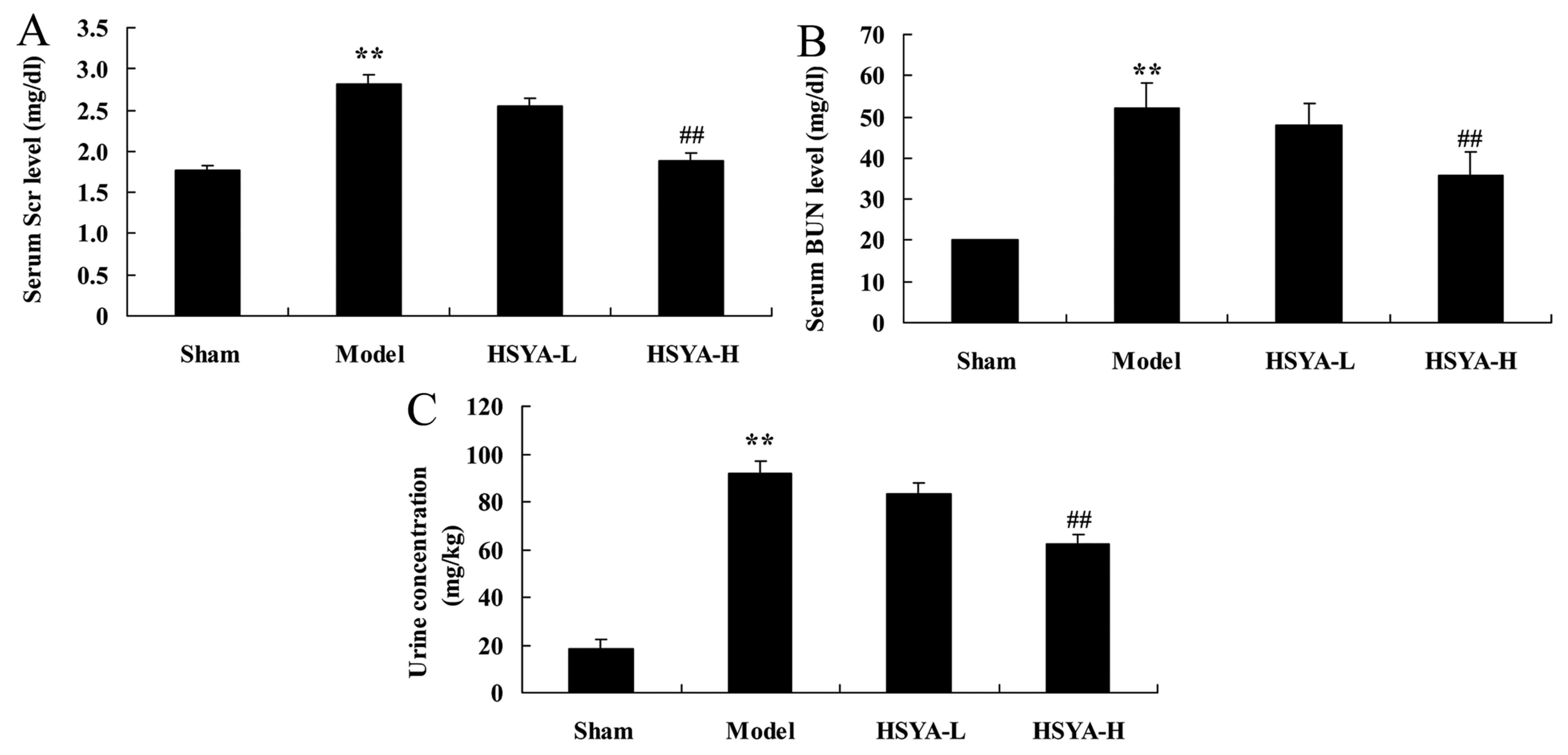
Effect of HSYA on biochemical
composition in the coronary heart disease model
Following 6 h of treatment, a significant increase
was observed in levels of Scr (P<0.01; Fig. 3A) BUN (P<0.01; Fig. 3B), and urine concentration
(P<0.01; Fig. 3C) in the model
group compared with the sham group. Treatment with high-does HSYA,
however, significantly ameliorated these effects (all P<0.01;
Fig. 3).
Effect of HSYA on the levels of
interleukin (IL-1β, IL-6, IL-10 and tumor necrosis factor (TNF)-α
in the coronary heart disease model
mRNA Levels of IL-1β, IL-6 and TNF-α were
significantly higher in the model group compared with the sham
group (all P<0.01; Fig. 4A-C),
and IL-10 expression was significantly downregulated in the model
group compared with the sham group (P<0.01; Fig. 4D). Treatment with high-dose HSYA
significantly reversed the increase in IL-1β, IL-6 and TNF-α
levels, and the decrease in IL-10 level in the model group
(P<0.01; Fig. 4).
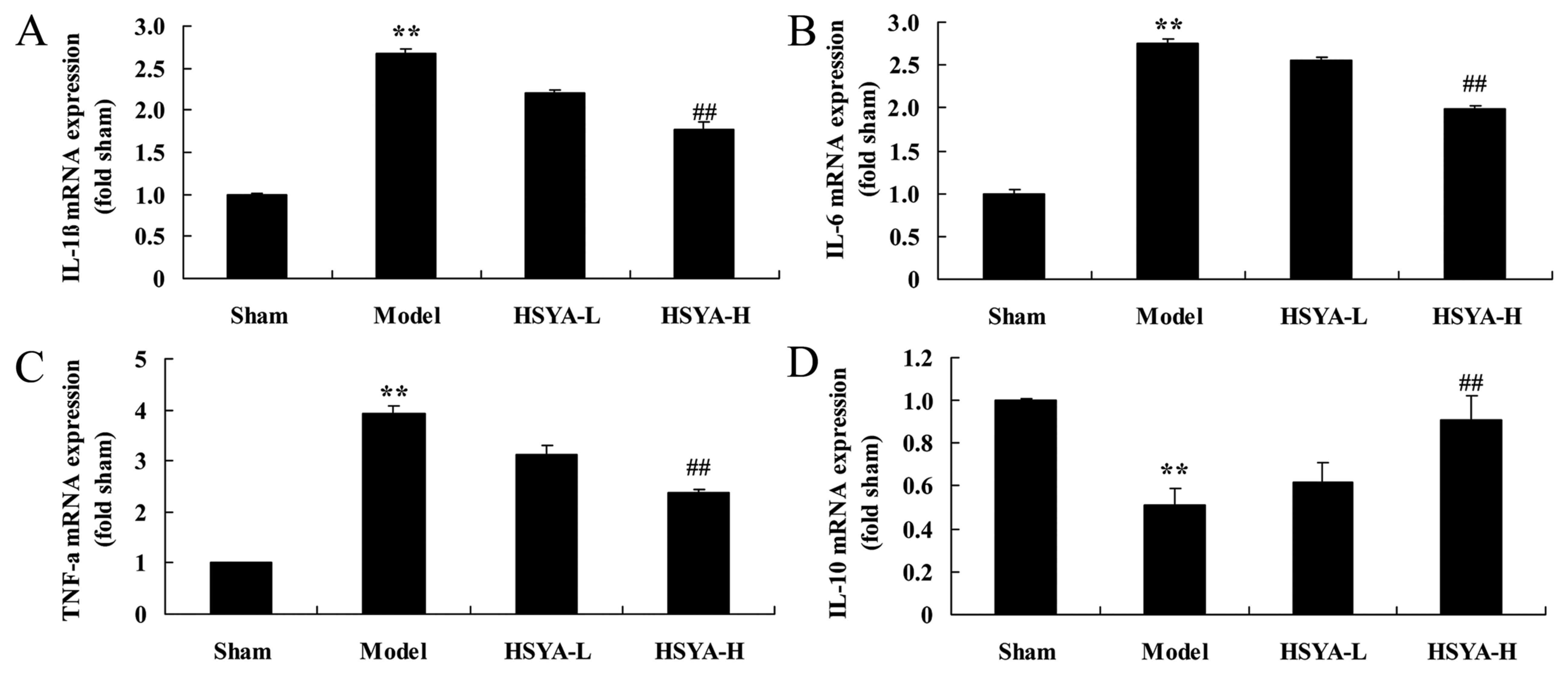 | Figure 4.Effect of HSYA on the mRNA levels of
IL-1β, IL-6, IL-10 and TNF-α in coronary heart disease model. The
effect of HSYA on the mRNA levels of (A) IL-1β, (B) IL-6, (C) TNF-α
and (D) IL-10 in a coronary heart disease model. **P<0.01 vs.
sham; ##P<0.01 vs. model. HSYA, hydroxy safflower
yellow A; IL, interleukin; TNF, tumor necrosis factor; sham, sham
group; model, coronary heart disease model group; HSYA-L, low dose
HSYA group; HSYA-H, high dose HSYA group. |
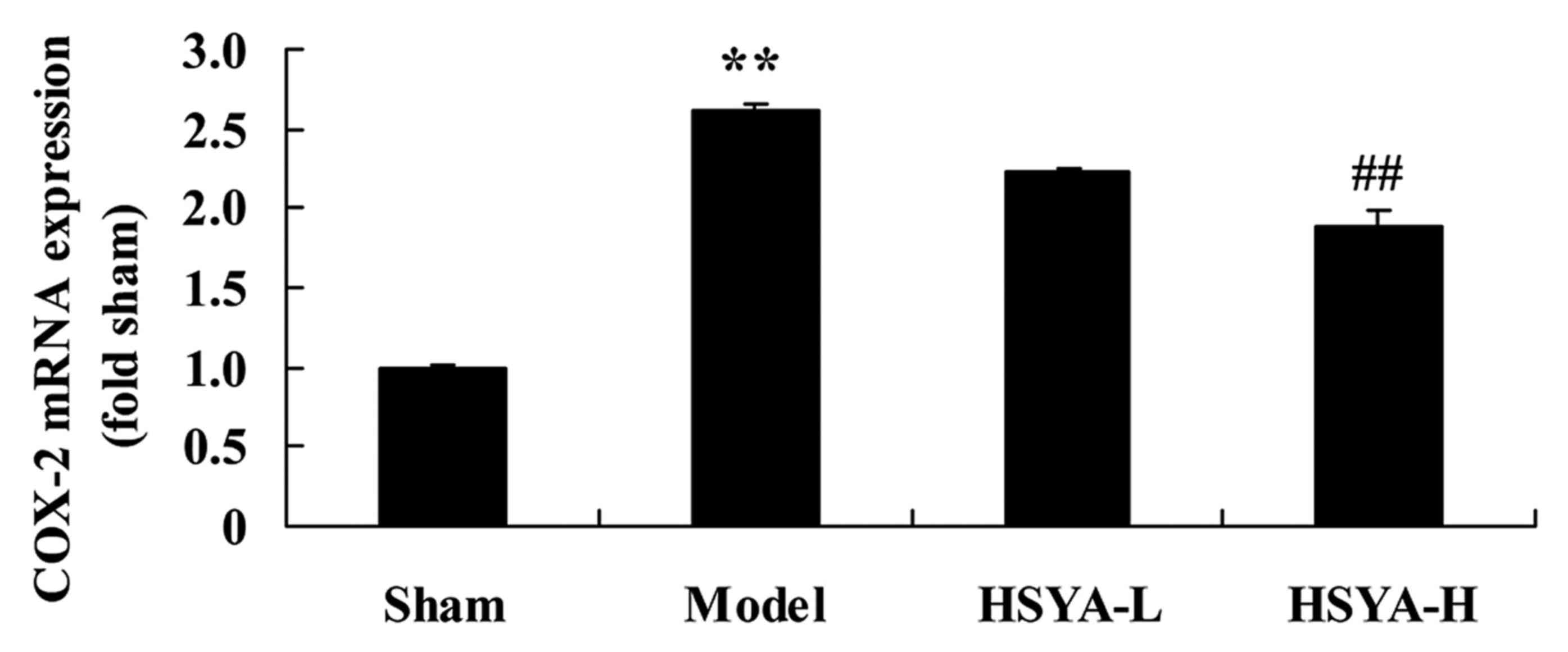
Effect of HSYA on the levels of COX-2
mRNA in the coronary heart disease model
The results of RT-qPCR revealed that the expression
of COX-2 mRNA was significantly higher in the model group compared
with the sham group (P<0.01; Fig.
5). Treatment with high-dose HSYA, however, significantly
suppressed the COX-2 mRNA levels in the model group (P<0.01;
Fig. 5).
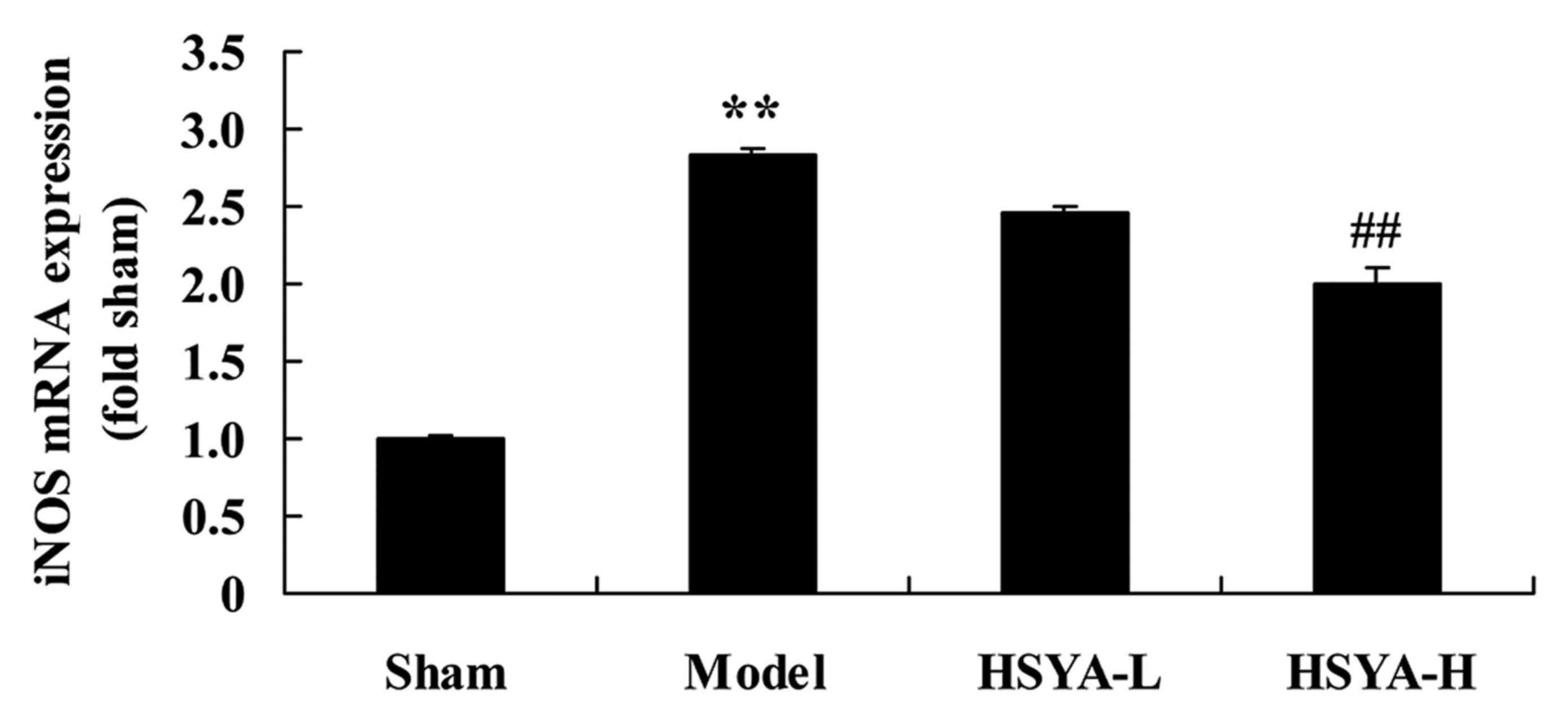
Effect of HSYA on the levels of iNOS
mRNA in the coronary heart disease model
A significant increase was observed in levels of
iNOS mRNA in the model group compared with the sham group
(P<0.01; Fig. 6). The HSYA-H
group exhibited a inhibited a significant downregulation in iNOS
mRNA compared with the model group (P<0.01; Fig. 6).
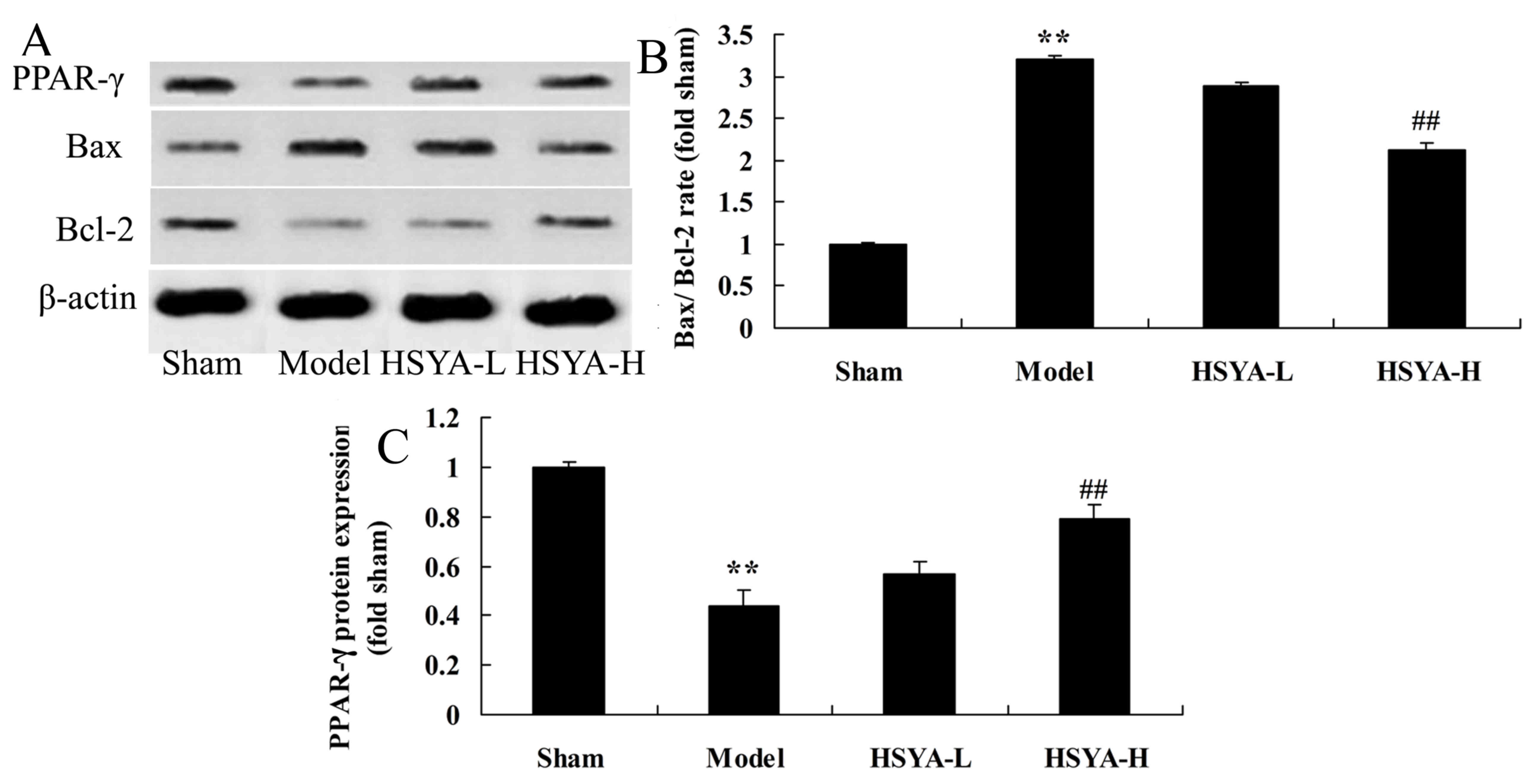
Effect of HSYA on the Bax/Bcl-2 ratio
and PPAR-γ protein expression in the coronary heart disease
model
Results of western blot analysis revealed that the
Bax/Bcl-2 ratio were significantly higher in the model group
compared with the sham group (P<0.01; Fig. 7). The Bax/Bcl-2 ratio was
significantly lower in the HSYA-H group compared with the model
group (P<0.01; Fig. 7). PPAR-γ
protein expression were significantly lower in the model group
compared with the sham group (P<0.01; Fig. 7). However, PPAR-γ protein expression
was significantly higher in the HSYA-H group compared with the
model group (P<0.01; Fig. 7).
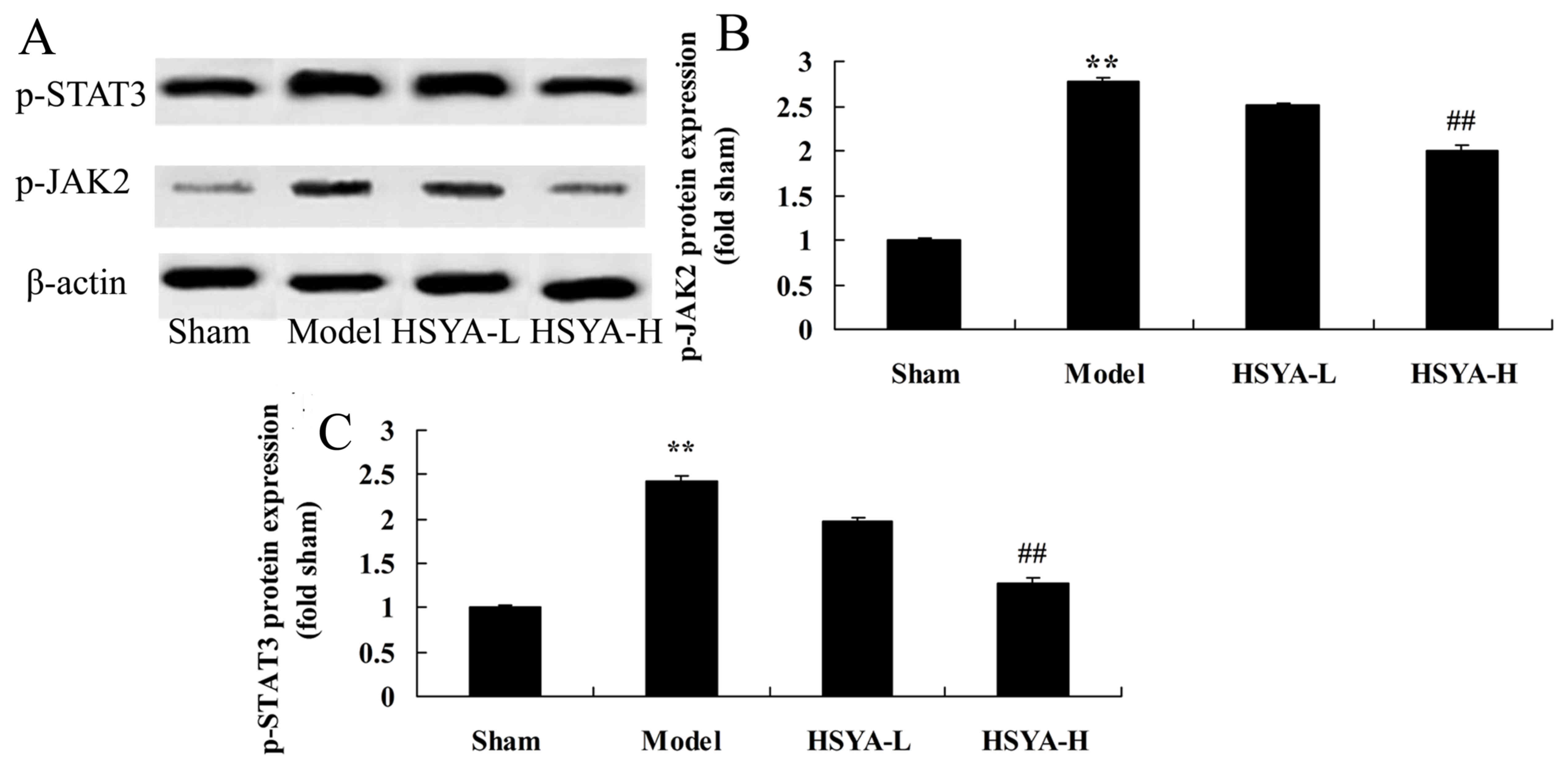
Effect of HSYA on the expression of
p-JAK2 and p-STAT3 protein in the coronary heart disease model
The results of western blot analysis revealed that
p-JAK2 and p-STAT3 levels were significantly higher in the model
group compared with sham group (P<0.01; Fig. 8). However, this increase was
ameliorated by treatment with high-dose HSYA (P<0.01; Fig. 8).
Discussion
Previous research has demonstrated that the
inflammatory response is associated with coronary heart disease
(4). Inflammatory markers and
endothelial function are both effective indicators for predicting
and treating coronary heart disease (4). IL-6 and other pro-inflammatory
cytokines are able to regulate the synthesis of coronary heart
disease and IL-6 is fundamental to this process (15). Similar to CRP, the imbalance of IL-6
may influence vary diseases (5). In
atherosclerosis, IL-6 primarily acts on blood vessel endothelium,
causing it to be impaired and therefore activating a series of
inflammatory responses (5), which
leads to coronary atherosclerotic cardiomyopathy or the conversion
of stable coronary heart disease to unstable (15). In the present study, HSYA was
demonstrated to significantly ameliorate LVEF, LVIDs and
biochemical composition, and suppressed the expression of IL-1β,
IL-6, IL-10 and TNF-α in a coronary heart disease model. Zhou et
al (13) reported that HSYA
weakens myocardial apoptosis following acute myocardial infarction
in rats via affecting Bax/Bcl-2 and PPAR-γ. Zhang et al
(12) demonstrated that HSYA
significantly reduced the IL-1β, IL-4, TNF-α, COX-2, iNOS, and
increased IL-10 mRNA levels in Alzheimer's disease via
phosphorylation of the JAK2/STAT3 pathway.
Endothelial dysfunction is the first stage of
coronary heart disease and manifests as a decrease in
endothelium-dependent vasodilation function, NO synthesis and the
disorder of angiokinesis factors (16). NO is synthesized under the function
of L-arginine. It is an important endothelium-derived relaxing
factor, which serves an important role in regulating angiokinesis
(17). Endothelium-dependent
vasodilatation is damaged following coronary atherosclerosis,
whereas contraction function is strengthened, resulting in the
reduction of myocardial perfusion and myocardial ischemia (17). Furthermore, NO is able to protect
blood vessels from injury and inhibit inflammatory responses and
thrombogenesis (16). It is also
able to inhibit leukocyte adhesion, platelet aggregation and enable
vascular smooth muscles to remain in a quiescent state (16). In the presence of clinical risk
factors such as hypertension, diabetes and hyperlipidemia, the
epithelial defense functions in blood vessels are disordered
(18). In the present study, HSYA
significantly downregulated iNOS and COX-2 mRNA levels in a
coronary heart disease model.
PPARγ is expressed in all vascular cells
(mononuclear leucocytes, macrophages and endothelial cells) and
vascular smooth muscle cells (10).
It serves a fundamental role in cell proliferation, migration,
differentiation, the activation of macrophages and vascular
endothelium inflammation (10).
PPARγ activated by vascular endothelial dysfunction may inhibit
TNF-α, IL-1β and IL-6 secreted by monocyte-macrophages as well as
the expression of platelet-activating factor receptor, VCAM-1, iNOS
and scavenger receptor A (11). It
is able to lower or limit the progress of chronic inflammation,
maintain the stability of membrane permeability, inhibit the
activity of smooth muscle cells and reduce inward flows of calcium
(11). Therefore, vascular
contractile responses and smooth muscle cell proliferation may be
inhibited and elastic resistance may be reduced (9). The results of the present study suggest
that HSYA significantly upregulated PPAR-γ protein expression in
swine with coronary heart disease. Zhou et al (13) reported that the effect of HSYA
weakens myocardial apoptosis following acute myocardial infarction
in rats through Bax/Bcl-2 and PPAR-γ.
Cell apoptosis is a type of programmed cell death
regulated by genes, which is induced by initiation of the apoptosis
signal pathway and the expression of related genes (19). The mitochondrial pathway is one of
the most important apoptosis signal transduction pathways (20). The occurrence and progression of cell
apoptosis is regulated by various genes, of which Bcl-2 serves a
fundamental role via the mitochondrial pathway (19). Bcl-2 is an anti-apoptosis gene while
Bax is the pro apoptotic gene (20).
Additionally, HSYA significantly reduced the Bax/Bcl-2 ratio in
swine with coronary heart disease. Zhou et al (13) reported that HSYA is able to weaken
myocardial apoptosis following acute myocardial infarction in rats
via the expression of Bax/Bcl-2 and PPAR-γ.
The JAK/STAT pathway is associated with the
inflammatory response, oxidative stress, cellular damage and
apoptosis (19). IL-6 is able to
worsen myocardial hypertrophy via activating the JAK/STAT pathway
(21). The JAK/STAT pathway mediates
the reduction in synthesis of the surface-active materials of
pulmonary epithelial cells caused by oxidative stress (22). Coronary heart disease is typically
accompanied by mitochondrial damage, which worsens coronary heart
disease (23). Oxidative damage to
mitochondria includes changes in permeability, swelling, oxidative
phosphorylation and the release of pro-apoptotic proteins (24). Recent studies revealed that the
activation or inhibition of the JAK2/STAT3 pathway may have a
significant effect on oxidative stress injuries and mitochondrial
function (23,25). In the present study, HSYA
significantly suppressed p-JAK2 and p-STAT3 protein expression in
swine with coronary heart disease. Zhang et al (12) demonstrated that HSYA was able to
significantly reduce the expression of IL-1β, IL-4, TNF-α, COX-2,
iNOS, and increase levels of IL-10 mRNA in patients with
Alzheimer's disease via the phosphorylation of the JAK2/STAT3
pathway.
In conclusion, the neuroprotective effects of HSYA
significantly improved LVEF and LVIDs and biochemical composition,
and suppressed the levels of IL-1β, IL-6, IL-10, TNF-α, iNOS and
COX-2 in a coronary heart disease model, which suggests that PPARγ,
Bax/Bcl-2 and the JAK2/STAT3 signaling pathway were affected by
treatment with HSYA. The results of the present study suggest that
HSYA may be a potential therapeutic treatment for coronary heart
disease in clinical scenarios.
References
|
1
|
Berndt N, Bolman C, Froelicher ES, Mudde
A, Candel M, de Vries H and Lechner L: Effectiveness of a telephone
delivered and a face-to-face delivered counseling intervention for
smoking cessation in patients with coronary heart disease: A
6-month follow-up. J Behav Med. 37:709–724. 2014.PubMed/NCBI
|
|
2
|
Moser DK, McKinley S, Riegel B, Doering
LV, Meischke H, Pelter M, Davidson P, Baker H and Dracup K: The
impact on anxiety and perceived control of a short one-on-one
nursing intervention designed to decrease treatment seeking delay
in people with coronary heart disease. Eur J Cardiovasc Nurs.
11:160–167. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Grunwald V, Karakiewicz PI, Bavbek SE,
Miller K, Machiels JP, Lee SH, Larkin J, Bono P, Rha SY, Castellano
D, et al: An international expanded-access programme of everolimus:
Addressing safety and efficacy in patients with metastatic renal
cell carcinoma who progress after initial vascular endothelial
growth factor receptor-tyrosine kinase inhibitor therapy. Eur J
Cancer. 48:324–332. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Derosa G, Bonaventura A, Bianchi L, Romano
D, D'Angelo A, Fogari E and Maffioli P: A randomized,
placebo-controlled study on the effects of a nutraceutical
combination of red yeast rice, silybum marianum and octasonol on
lipid profile, endothelial and inflammatory parameters. J Biol
Regul Homeost Agents. 28:317–324. 2014.PubMed/NCBI
|
|
5
|
Kuravi SJ, McGettrick HM, Satchell SC,
Saleem MA, Harper L, Williams JM, Rainger GE and Savage CO:
Podocytes regulate neutrophil recruitment by glomerular endothelial
cells via IL-6-mediated crosstalk. J Immunol. 193:234–243. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen G, Qiu H, Ke S, Hu S, Yu S and Zou S:
The fibroblast growth factor receptor 2-mediated extracellular
signal-regulated kinase 1/2 signaling pathway plays is important in
regulating excision repair cross-complementary gene 1 expression in
hepatocellular carcinoma. Biomed Rep. 1:604–608. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Haga S, Tsuchiya H, Hirai T, Hamano T,
Mimori A and Ishizaka Y: A novel ACE2 activator reduces
monocrotaline-induced pulmonary hypertension by suppressing the
JAK/STAT and TGF-β cascades with restored caveolin-1 expression.
Exp Lung Res. 41:21–31. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Han JK, Kim HL, Jeon KH, Choi YE, Lee HS,
Kwon YW, Jang JJ, Cho HJ, Kang HJ, Oh BH, et al: Peroxisome
proliferator-activated receptor-δ activates endothelial progenitor
cells to induce angio-myogenesis through matrix
metallo-proteinase-9-mediated insulin-like growth factor-1
paracrine networks. Eur Heart J. 34:1755–1765. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ellis HP and Kurian KM: Biological
rationale for the use of PPARγ agonists in glioblastoma. Front
Oncol. 4:522014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aydoğan HY, Küçükhüseyin O, Tekeli A and
Isbir T: Associations of receptor for advanced glycation end
products-374 T/A and Gly82 Ser and peroxisome
proliferator-activated receptor gamma Pro12Ala polymorphisms in
Turkish coronary artery disease patients. Genet Test Mol
Biomarkers. 16:134–137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang TH and Roufogalis BD: Healing the
diabetic heart: Modulation of cardiometabolic syndrome through
peroxisome proliferator activated receptors (PPARs). Curr Mol
Pharmacol. 5:241–247. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Z, Wu Z, Zhu X, Hui X, Pan J and Xu
Y: Hydroxy-safflor yellow A inhibits neuroinflammation mediated by
Aβ(1)(−)(4)(2) in BV-2 cells. Neurosci Lett. 562:39–44. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou MX, Fu JH, Zhang Q and Wang JQ:
Effect of hydroxy safflower yellow A on myocardial apoptosis after
acute myocardial infarction in rats. Genet Mol Res. 14:3133–3141.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kaysen GA, Levin NW, Mitch WE, Chapman AL,
Kubala L and Eiserich JP: Evidence that C-reactive protein or IL-6
are not surrogates for all inflammatory cardiovascular risk factors
in hemodialysis patients. Blood Purif. 24:508–516. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dusting GJ: Nitric oxide in coronary
artery disease: Roles in atherosclerosis, myocardial reperfusion
and heart failure. EXS. 76:33–55. 1996.PubMed/NCBI
|
|
17
|
Baptista J, Teles RC, da Silva PC, Pereira
H, Marques L, Santos R, Carvalho H, Martins D, Farto e Abreu P,
Araújo J, et al: Five-year clinical results of coronary angioplasty
with drug-eluting stents. National initiative in strategic
innovation, iNOS. Rev Port Cardiol. 29:243–251. 2010.(In English,
Portuguese). PubMed/NCBI
|
|
18
|
Meng QH, Irvine S, Tagalakis AD, McAnulty
RJ, McEwan JR and Hart SL: Inhibition of neointimal hyperplasia in
a rabbit vein graft model following non-viral transfection with
human iNOS cDNA. Gene Ther. 20:979–986. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wisel S, Khan M, Kuppusamy ML, Mohan IK,
Chacko SM, Rivera BK, Sun BC, Hideg K and Kuppusamy P:
Pharmacological preconditioning of mesenchymal stem cells with
trimetazidine (1-[2,3,4-trimethoxybenzyl]piperazine) protects
hypoxic cells against oxidative stress and enhances recovery of
myocardial function in infarcted heart through Bcl-2 expression. J
Pharmacol Exp Ther. 329:543–550. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Zhang H, Chai F, Liu X and Berk M:
The effects of escitalopram on myocardial apoptosis and the
expression of Bax and Bcl-2 during myocardial ischemia/reperfusion
in a model of rats with depression. BMC Psychiatry. 14:3492014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Roxburgh CS and McMillan DC: Therapeutics
targeting innate immune/inflammatory responses through the
interleukin-6/JAK/STAT signal transduction pathway in patients with
cancer. Transl Res. 167:61–66. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wen SH, Li Y, Li C, Xia ZQ, Liu WF, Zhang
XY, Lei WL, Huang WQ and Liu KX: Ischemic postconditioning during
reperfusion attenuates intestinal injury and mucosal cell apoptosis
by inhibiting JAK/STAT signaling activation. Shock. 38:411–419.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mo ZC, Xiao J, Liu XH, Hu YW, Li XX, Yi
GH, Wang Z, Tang YL, Liao DF and Tang CK: AOPPs inhibits
cholesterol efflux by down-regulating ABCA1 expression in a
JAK/STAT signaling pathway-dependent manner. J Atheroscler Thromb.
18:796–807. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lu Y, Zhou J, Xu C, Lin H, Xiao J, Wang Z
and Yang B: JAK/STAT and PI3K/AKT pathways form a mutual
transactivation loop and afford resistance to oxidative
stress-induced apoptosis in cardiomyocytes. Cell Physiol Biochem.
21:305–314. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Negoro S, Kunisada K, Tone E, Funamoto M,
Oh H, Kishimoto T and Yamauchi-Takihara K: Activation of JAK/STAT
pathway transduces cytoprotective signal in rat acute myocardial
infarction. Cardiovasc Res. 47:797–805. 2000. View Article : Google Scholar : PubMed/NCBI
|