Introduction
Centipedegrass belongs to the genus
Eremochloa (Poaceae), which includes eight species that
inhabit China and Southeast Asia, and is one of the most popular
grasses in South America (1). It
contains C-glycosyl flavones and phenolic constituents as its
common structural skeleton, which is biologically active (2). In a previous study, we isolated and
analyzed the extract from centipedegrass, and found that these
components included maysin, and maysin derivatives such as
luteoin-6-C-boivinopyranose, luteolin, Isoorientin,
rhamnosylisoorientin, and derhamnosylmaysin (3). Particularly, centipedegrass extract
(CGE), a flavonoid-rich chemical compound extracted from
centipedegrass, has been reported to demonstrate free
radical-scavenging activity in vitro biochemical assays
using DPPH-radical scavenging activity (4). It was also reported that maysin (a
C-glycoside flavone from centipedegrass) and its precursor chemical
components could exhibit anti-oxidative activity via DPPH radical
scavenging in vitro antioxidant model (5), and protect SK-N-MC cell lines against
inhibition of H2O2-induced apoptotic cell
death (6). Despite these health
benefits of maysin and flavonoid derivatives, its cytoprotective
effects against ionizing radiation (IR)-induced cell death in
fibroblasts and the underlying mechanisms have not been elucidated
yet.
Cancer remains a big challenge in global healthcare
because it is one of the most rampant causes of death. Radiation
therapy (RT) is one of the most important treatment strategies used
in the treatment of more than two thirds of cancer cases worldwide
(7). Advances in technology over
the last decades have prompted the development of 3-dimensional
conformal RT techniques, including intensity-modulated radiation
therapy and stereotactic body radiation therapy (8). In addition to these advanced medical
technologies, therapeutic strategies in radiation oncology have
grown significantly with advances in physiology, immunology, and
molecular biology, allowing us to explore the treatment outcome
from a better, all-inclusive perspective, and perform radiotherapy
more efficiently.
However, long-term stimulation of IR in human
tissues unavoidably leads to the development of free radical
pathology. Generally, IR affects the body of patients indirectly;
for example, upon interaction with water molecules in cells exposed
to radiation, large amounts of free radicals and reactive oxygen
species (ROS) are produced, which oxidize cellular components,
causing cellular damage (9). The
cells that are exposed to radiation respond via generation of
antioxidant enzymes depending on the degree of exposure, followed
which free radical-induced damage to the cellular structure is
minimized or eliminated (10).
However, when ROS are produced due to exposure of the body to
radiation and excessive amounts of free radicals, they cannot be
completely removed by intrinsic antioxidative enzymes.
Radiation-induced skin reaction (RISR) is one of the
main side effects of radiotherapy, and more than 95% of patients
undergoing RT experience tissue damage (11). Moreover, acute RISR often has a big
impact on the progress of RT due to limitation of the total
therapeutic dose or breaks in radiotherapy (12). Radiation-induced oxidative stress is
known to be the main cause of RISR (13). Oxidative stress induced by IR
results in damages of DNA, lipids, and proteins. The harmful
effects stimulate early transcription factors and molecular signals
leading to cellular damage. As a result, it causes damage to the
skin tissue (14). Thus, in
radiotherapy, IR-induced oxidative stress is an important variable
before and after IR, and it is essential to scavenge the IR-induced
free radicals or ROS to reduce/mitigate the damage caused by
radiation.
Fibroblasts, the main type of cells constituting the
dermal skin, play important roles in the development of healthy
skin by producing the extracellular matrix and collagen. The
occurrence of damaged fibroblasts, resulting in the presence of low
levels of extracellular matrix proteins, is the result of skin
aging, and is consequently responsible for the formation of
wrinkles (15). IR-induced
oxidative stress has been reported to cause damage with ROS
inducing apoptosis in a variety of cells, including fibroblasts and
keratinocytes, thereby reducing cell numbers and regenerative
capacity (16). Additionally,
persistent oxidative stress during RT can lead to severe cellular
damage and irreversible tissue conditions, which makes tissue
regeneration impossible (17). This
consideration marked the beginning of the active development of a
radioprotector for the prevention and treatment of IR-induced free
radical pathology with the use of antioxidant agents (18). Studies involving the use of
vitamins, amino acids, animal- and plant-based agents containing
antioxidant enzymes are considered to be highly significant but
less developed in the field of radiobiology. Considering the lack
of efficient radioprotective agents made of natural raw materials,
many studies have been undertaken with the purpose of finding
radioprotective agents (19,20).
In this study, CGE, which contains flavonoid
derivatives, was evaluated for its radioprotective effects against
IR-induced cell damage in NIH-3T3 fibroblast cells.
Materials and methods
Preparation of CGE
CGE was prepared using the method described in our
previous report (21). Briefly,
seeds of centipedegrass imported from the Fukukaen Nursery (Blu Co.
Ltd.) were cultivated at the Korea Atomic Energy Research Institute
in 2016 (KAERI). The leaves of the centipedegrass were harvested
and stored at -80˚C until use. Dry leaves of centipede (5 kg) were
crushed in a Wiley mill (Indian Weiber) and filtered through a
sieve of pore size 420 µm. The final sample of ground leaves (1 kg)
was extracted thrice using 80% methanol (MeOH, 100 liters; Merck)
for 24 h with constant shaking at ambient temperature in the dark.
The extracts were filtered using a No. 2 filter paper (Advantech)
and concentrated in vacuum. MeOH extracts were fractionated
successively using n-hexane and ethyl acetate (EA). The layer
separated using EA was concentrated in vacuum and the dried
compound was dissolved in MeOH. The active MeOH extracts were
diluted with 20% MeOH and chromatographed on a Toyopearl HW-40C
resin (Tosoh Corp.) column using 70% MeOH (elution volume, 700 ml).
Fractions were evaporated and lyophilized. The dried extract was
diluted in dimethyl sulfoxide (DMSO) to carry out further
experiments. Lastly, to confirm the active substances content,
maysin, and maysin derivatives (the active ingredient of CGE) were
confirmed by high-performance liquid chromatography/mass (HPLC-MS)
analysis as previously described method (3).
Chemicals and reagents
All the chemicals and reagents were used without
further purification. A cell counting kit-8 was purchased from
Sigma-Aldrich; Merck KGaA. Anti-AKT (#4691), anti-p-AKT (#4060),
anti-ERK (#4695), anti-p-ERK (#4377), anti-p38 (#9212), anti-p-p38
(#9215), anti-JNK (#9252), anti-p-JNK (#9251), anti-caspase-3
(#9662), anti-cleaved caspase-3 (#9664), anti-Bcl-2 (#2870),
anti-Bax (#5023), anti-Bad (#9292), anti-p-Bad (#9291),
anti-poly(ADP-ribose) polymerase (PARP) (#9532), anti-cleaved PARP
(#9541), anti-GAPDH (#2118), and anti-α-tubulin (#2144) were
purchased from Cell Signaling Technology, Inc. Dulbecco's modified
Eagle's medium (DMEM), penicillin/streptomycin (P/S), and fetal
bovine serum (FBS) were purchased from Lonza. Annexin V and
oxidative stress kits were purchased from Millipore.
Cell culture
The NIH-3T3 cell line (mouse, fibroblast) was
obtained from the Korean Cell Line Bank (Seoul, Korea). The cells
were cultured under sterile conditions at 37˚C in a humid
environment containing 5% CO2. The culture medium
consisted of DMEM supplemented with FBS (10%), glutamine (4 mM),
and penicillin/streptomycin (1%).
Morphological analysis and cell
viability
The morphology of NIH-3T3 cells was monitored using
an Olympus IX71 fluorescence microscope (Olympus Corp.). In order
to determine the half-maximal inhibitory concentration
(IC50 value) of H2O2 or the
effects of CGE on NIH-3T3 cells, 1x104 cells (from a
single-cell suspension) were seeded into individual wells of
96-well plates and incubated for 24 h at 37˚C before
H2O2 treatment (125-500 µM). For
H2O2 treatment, each well received a 1:2
sequential dilution of H2O2 from 500 µM to
125 µM. Alternatively, CGE was also added sequentially to the wells
(25-100 µg/ml), after which, the cells were incubated for 24 h.
0.1% DMSO was used as vehicle control. CCK-8 solution was added to
each well and the plates were incubated for 1 h at 37˚C to allow
the reaction to take place before removal of the culture medium.
Cell viability was determined using a spectrophotometer and the
absorbance was measured at 450 nm (Tecan). The cell viability for
each group was calculated as a percentage of that of the control
group.
Measurement of intracellular ROS
levels
NIH-3T3 cells (1x105 cells/well for
fluorescence and oxidative stress assays, 1x104 cells
for the DCF-DA assay) grown on coverslips in 6-well/96-well plates
were incubated with CGE (100 µg/ml) for 24 h at 37˚C before
H2O2 treatment (500 µM) or IR exposure (4
Gy). The following procedures were conducted: i) DCF-DA staining:
the cells were treated with DCF-DA according to the manufacturer's
instructions and examined using a fluorescence microscope or
microplate reader (excitation: 485 nm, emission: 535 nm). ii)
Oxidative stress assay: Cellular populations undergoing oxidative
stress were measured quantitatively using the Muse™ Cell Analyzer
and Muse™ Oxidative Stress kit (EMD Millipore). According to the
manufacturer's protocol, the cells were detached, resuspended to
obtain 1x106 cells/ml and incubated at 37˚C for 30 min
with the Muse™ Oxidative Stress working solution. The number of
oxidized cells was counted using the Muse™ Cell Analyzer based on
the intensity of red fluorescence. The results were obtained from
four independent experiments.
Apoptotic assay
NIH-3T3 cells (3x105 cells/well) grown in
a 65-mm culture dish were incubated with CGE (100 µg/ml) for 24 h
at 37˚C before H2O2 treatment (500 µM). The
following procedures were conducted to determine apoptosis in the
cells: i) Fluorescence imaging: Cells were incubated with DAPI
according to the manufacturer's instructions and examined using a
confocal microscope (Zeiss AG). ii) Annexin V assay: To
quantitatively analyze the apoptotic and necrotic dead cells, Muse
Annexin V and Dead Cell Assay kits (MCH100105; EMD Millipore) were
used. The cells were harvested and washed with DPBS. The cells were
then stained with Annexin V and the Dead Cell reagent for 20 min,
following which flow cytometric assessment was performed using the
Muse™ Cell Analyzer. The number of apoptotic cells were expressed
as the percentage of the live, early/late apoptotic, and dead
cells, which was determined by the Muse analysis software (Muse
1.1.2; EMD Millipore).
gamma-ray irradiation
gamma-ray irradiation was performed at room
temperature at the dose rate of 1 Gy/min using a Cs137
gamma cell (MDS Nordion). The cells were directly irradiated in the
flasks. After irradiation, they were incubated at 37˚C and 5%
CO2 until they could be harvested for direct
experimentation or were stored at appropriate temperatures until
further use.
Clonogenic assay
The NIH-3T3 cells (1,000 cells/well) grown on
coverslips in 6-well plates were incubated with CGE (100 µg/ml) for
24 h at 37˚C prior to IR exposure (4 Gy). After exposure to various
doses (1-12 Gy) of gamma-rays, the cells were incubated with fresh
media for 7 days, fixed using pure MeOH for 20 min at room
temperature and stained using Wright stain (Thermos Fisher
Scientific, Inc.) for 20 min. The number of colonies containing
more than 50 cells was counted.
Western blot analysis
NIH-3T3 cells were washed with PBS and lysed with
radioimmunoprecipitation assay buffer. The proteins (30-50 µg) were
separated via 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and were then transferred onto polyvinylidene
difluoride membranes. The membranes were blocked with 5% non-fat
dry milk for 1 h at room temperature and then incubated overnight
with 1:1,000-diluted primary antibodies at 4˚C. The membranes were
washed with tris-buffered saline and incubated with horseradish
peroxidase (HRP)-conjugated secondary antibodies (anti-rabbit IgG,
HRP-linked antibody, #7074; Cell Signaling Technology, Inc.) for 2
h at room temperature. The proteins were then visualized using an
enhanced chemiluminescence reagent (Millipore Corp.) and exposure
to an X-ray film.
Statistical analysis
Each experiment was performed at least three times,
and the results were expressed as the mean ± standard deviation.
For multiple comparisons, one-way analysis of variance was used
followed by Tukey's multiple comparisons test. P≤0.05 was
considered to indicate a statistically significant difference.
Results
CGE protects NIH-3T3 cells from
H2O2-induced damage
The cytotoxic effect of CGE was determined by
measuring the viability of cells treated with increasing
concentrations of CGE using a colorimetric CCK-8 assay. Stimulation
with CGE did not have a significant effect on cell viability at any
tested concentration (25-100 µg/ml) over a period of 24 h (Fig. 1A). Thus, all the subsequent
experiments were conducted at the concentration of 100 µg/ml. Next,
the effect of H2O2 (potent
pro-oxidant)-induced damage in NIH-3T3 cells was examined to
determine the protective effect of CGE against oxidative stress. It
was observed that the viability of
H2O2-stimulated NIH-3T3 cells had reduced to
65 and 43% at the concentrations of 250 and 500 µM, respectively
(Fig. 1B). Therefore, the cell
viability was evaluated next, and the cellular morphologies were
observed to examine whether CGE exhibits any protective effect in
NIH-3T3 cells stimulated with 500 µM of H2O2.
As shown in Fig. 1C, CGE exhibited
a protective effect on H2O2-induced
cytotoxicity; additionally, this effect was confirmed by a decrease
in the irregular morphology observed due to
H2O2-induced cellular damage (Fig. 1D). These data indicate that CGE
could inhibit H2O2-induced cytotoxicity in
NIH-3T3 cells.
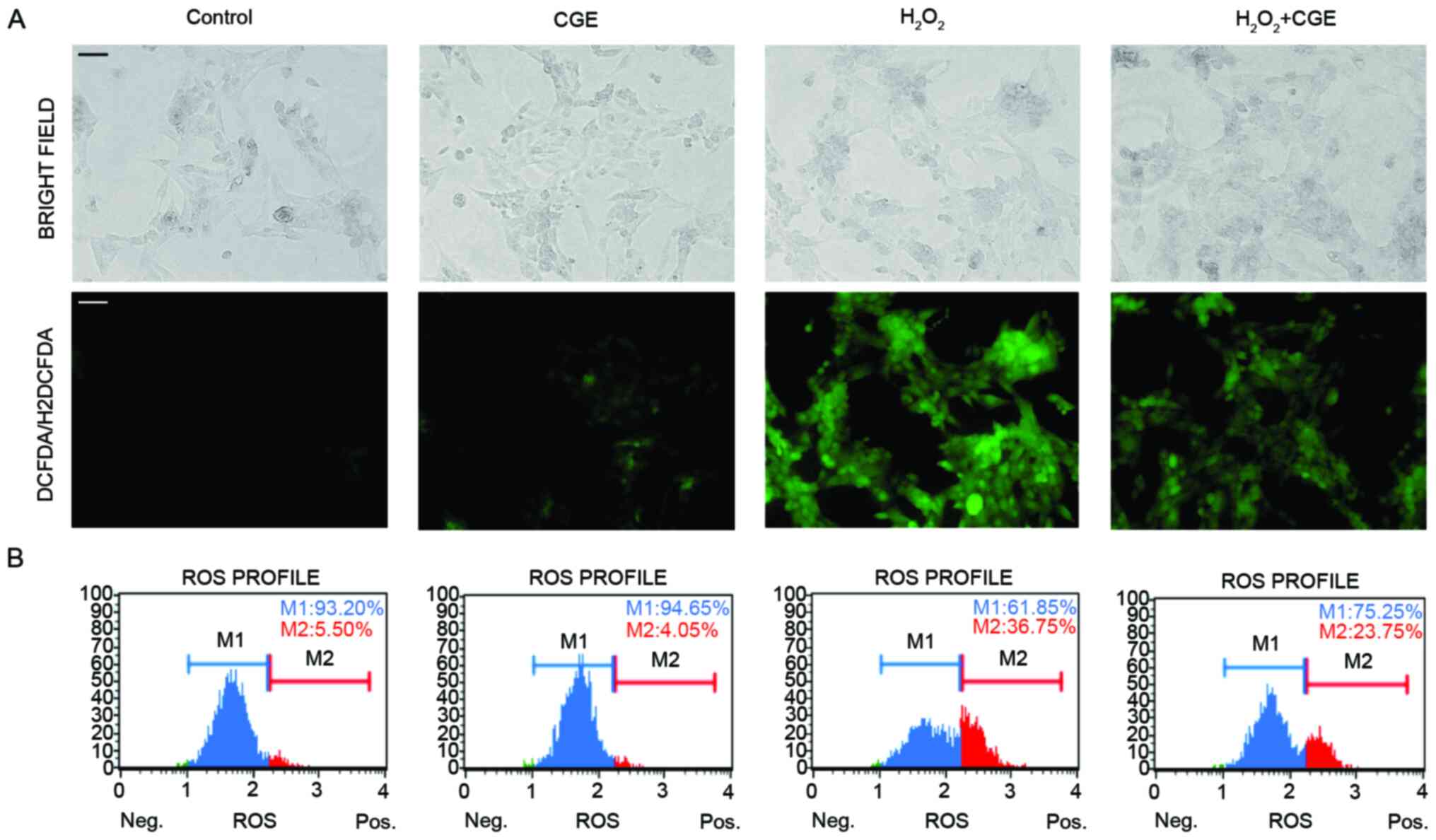
CGE regulates
H2O2-mediated intracellular ROS levels
To investigate the effect of CGE on ROS levels, we
determined whether CGE treatment could attenuate
H2O2-mediated oxidative stress. The NIH-3T3
cells were stimulated with H2O2 in the
presence or absence of CGE, after which, the production of ROS was
observed using the fluorescent probe
2,7-dichlorodihydrofluorescein-diacetate (H2DCFDA) of the cellular
ROS detection kit. Treatment with H2O2 alone
resulted in robust intracellular generation of ROS, whereas
H2O2-induced ROS generation in NIH-3T3 cells
was significantly attenuated by CGE treatment (Fig. 2A). Moreover, decrease in the
oxidative stress was quantified via the dihydroethidium (DHE)
reaction. DHE is cell permeable and is considered to react with
superoxide anions, thus undergoing oxidation upon binding to DNA
(22). As shown in Fig. 2B, CGE treatment led to a decrease in
the number of cells demonstrating high ROS production. A
significant decrease in the oxidative stress was observed in the
NIH-3T3 cells. These results implied that pretreatment with CGE
reduces the accumulation of intracellular ROS in NIH-3T3 cells.
CGE inhibits
H2O2-induced cell death in NIH-3T3 cells
To elucidate the protective effects of CGE against
H2O2-induced damage in NIH-3T3 cells, the
effects of CGE on H2O2-mediated apoptosis
were investigated. The results of the cell nucleus staining showed
that treatment with H2O2 alone had
significantly increased the number of cells with condensed or
blebbing nuclei. Contrarily, when cells were pretreated with CGE,
the nuclear damages were markedly reduced (Fig. 3A). The results of the flow cytometry
consistently indicated that H2O2 treatment
had increased the population of Annexin V+/PI-apoptotic
cells. However, as shown in Fig.
3B, pretreatment of the cells with CGE prior to their exposure
to H2O2 led to effective protection of the
cells against apoptosis. Likewise, results of western blotting
indicated a significant increase in the expression of cleaved PARP,
a well-known substrate of caspase-3 in the apoptotic cell death,
compared to that of the control. However, in alignment with
previous results, Fig. 3C showed
that the H2O2-induced PARP degradation was
reduced by pretreatment of the cells with CGE. Taken together,
these results indicated that CGE pretreatment could inhibit
H2O2-induced apoptotic cell death in NIH-3T3
cells.
CGE increases the viability of
IR-exposed NIH-3T3 cells
To investigate whether the IR-induced cell damage
could be prevented by CGE pretreatment, first, the morphological
changes in NIH-3T3 cells were evaluated. As shown in Fig. 4A, it was confirmed that pretreatment
with CGE could decrease the irregular morphology observed in
IR-exposed NIH-3T3 cells. In addition, to further investigate the
relationship between cell fractions retaining reproductive
integrity and absorbed radiation dose, the clonogenic survival
curves were determined. The cell survival fraction in the
IR-exposed NIH3/T3 cells was evaluated by the cell survival curve.
As shown in Fig. 4B, IR caused cell
death in proportion to the amount of exposure. However,
pretreatment with CGE significantly increased cell viability 24 h
after irradiation over the full range of doses. Particularly, the
50% lethal dose (LD 50%) of cells pretreated with CGE reduced from
4.51 to 3.41 Gy. Previous experiments have confirmed that CGE is
capable of inhibiting H2O2-induced cell
damage. As IR has been shown to adversely affect cells via ROS
generation, we investigated whether one possible mechanism for the
cytoprotective effect of CGE against damages induced by IR is based
on its ROS scavenging capacity. To investigate whether CGE could
reduce IR-induced ROS generation, H2DCFDA was used for a
fluorescence assay. Fig. 4C
represents the quantitative analysis of the cellular ROS levels.
When the cells were pretreated with CGE, the fluorescence intensity
of H2DCFDA was found to be significantly reduced. These results
demonstrated that IR caused cell damage by inducing overproduction
and accumulation of intracellular ROS, which was effectively
prevented by CGE.
CGE inhibits apoptosis in IR-exposed
NIH-3T3 cells via regulation of the ERK-, p38-, JNK-MAPKs
signaling
Radiation is known to mainly induce intrinsic
apoptotic cascades such as the mitochondrial release of cytochrome
c and subsequent formation of the apoptosome. However, depending on
dosage and cell type, the extrinsic apoptotic pathway might also
induce cell death (23). In this
study, to explore the possible molecular mechanism underlying the
radioprotective effects of CGE in NIH-3T3 cells exposed to
radiation, first, the protein expression related to the intrinsic
apoptotic pathway was examined. The intrinsic apoptotic pathway is
controlled and regulated by the activities of the members of the
Bcl-2 protein family involved in mitochondrial membrane
permeability and which may be pro-apoptotic or anti-apoptotic
(24). As shown in Fig. 5, radiation activates the expression
of the pro-apoptotic Bcl-2 proteins Bax (Bcl-2 like protein 4),
Bad, PARP, and cleaved caspase-3, whereas it inhibits the
expression of the anti-apoptotic protein Bcl-2. However, when cells
were pretreated with CGE, the expression of anti-apoptotic proteins
was significantly decreased. Additionally, pretreatment with CGE
has been shown to lead to phosphorylation of BAD by inducting AKT
phosphorylation, thereby rendering these signal proteins inactive
in apoptotic processes.
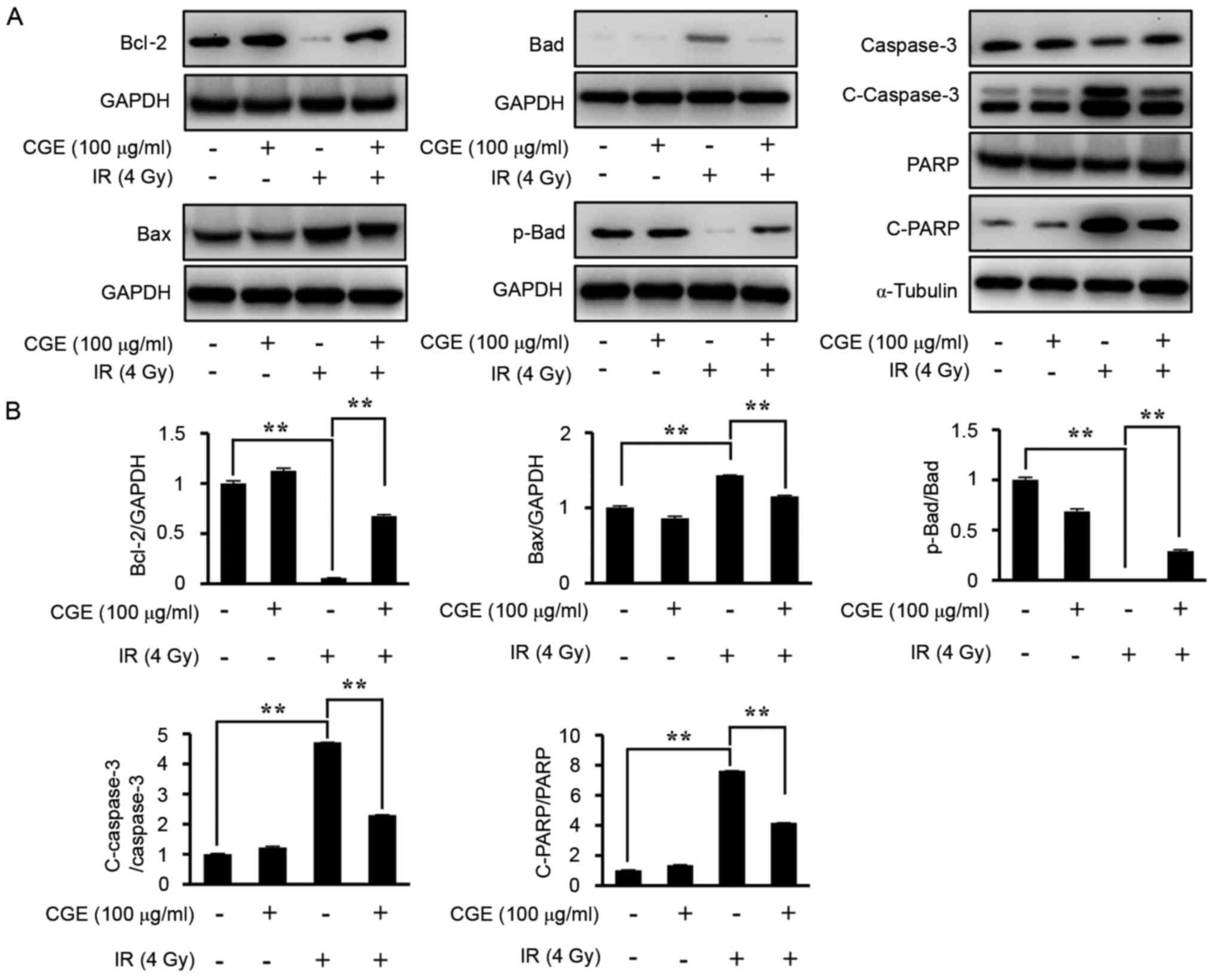 | Figure 5IR exposure-induced oxidative stress
and CGE treatment resulted in altered apoptosis signaling in
NIH-3T3 cells. (A) Lysates of IR-exposed cells in the presence or
absence of CGE were immunoblotted with anti-Bcl-2, anti-Bax,
anti-Bad, anti-p-Bad, anti-caspase-3, anti-C-caspase-3, anti-PARP,
anti-C-PARP, or anti-GAPDH antibodies. (B) The western blots were
analyzed quantitatively. Band intensities were normalized to those
of the normal form of each protein, GAPDH, or α-tubulin.
**P≤0.01 vs. IR only. IR, ionizing radiation; CGE,
centipedegrass extract; Bcl-2, B-cell lymphoma-2; Bax, Bcl-2 like
protein 4; p-, phosphorylated; Bad, Bcl-2-associated death
promoter; PARP, poly(ADP-ribose) polymerase; GAPDH, glyceraldehyde
3-phosphate dehydrogenase; Con, vehicle control (0.1% DMSO); SD,
standard deviation. |
Previous studies have reported that exposing cells
to IR and various other toxic stresses can induce simultaneous
target activation of multiple MAPK pathways (25). This has been related with the
factor-mediated regulation of various cell longevity factors such
as proliferation, differentiation, aging, and apoptosis. Thus, the
expression of MAPK family (ERK1/2, p38 and JNK), which plays a role
in the extrinsic apoptotic pathway, in IR-exposed cells was
explored using immunoblot analysis. Fig. 6A and B shows that the expression levels of p-ERK
in IR-exposed NIH-3T3 cells were downregulated whereas those of
p-JNK and p-p38 were upregulated. Conversely, these changes in the
expression levels were largely alleviated by pretreatment with CGE.
Collectively, these results supported the hypothesis that
pretreatment with CGE protects the NIH-3T3 cells against IR-induced
cell death via inhibition of the intrinsic apoptotic pathway as
well as the regulation of the ERK-, p38-, JNK-MAPKs signaling.
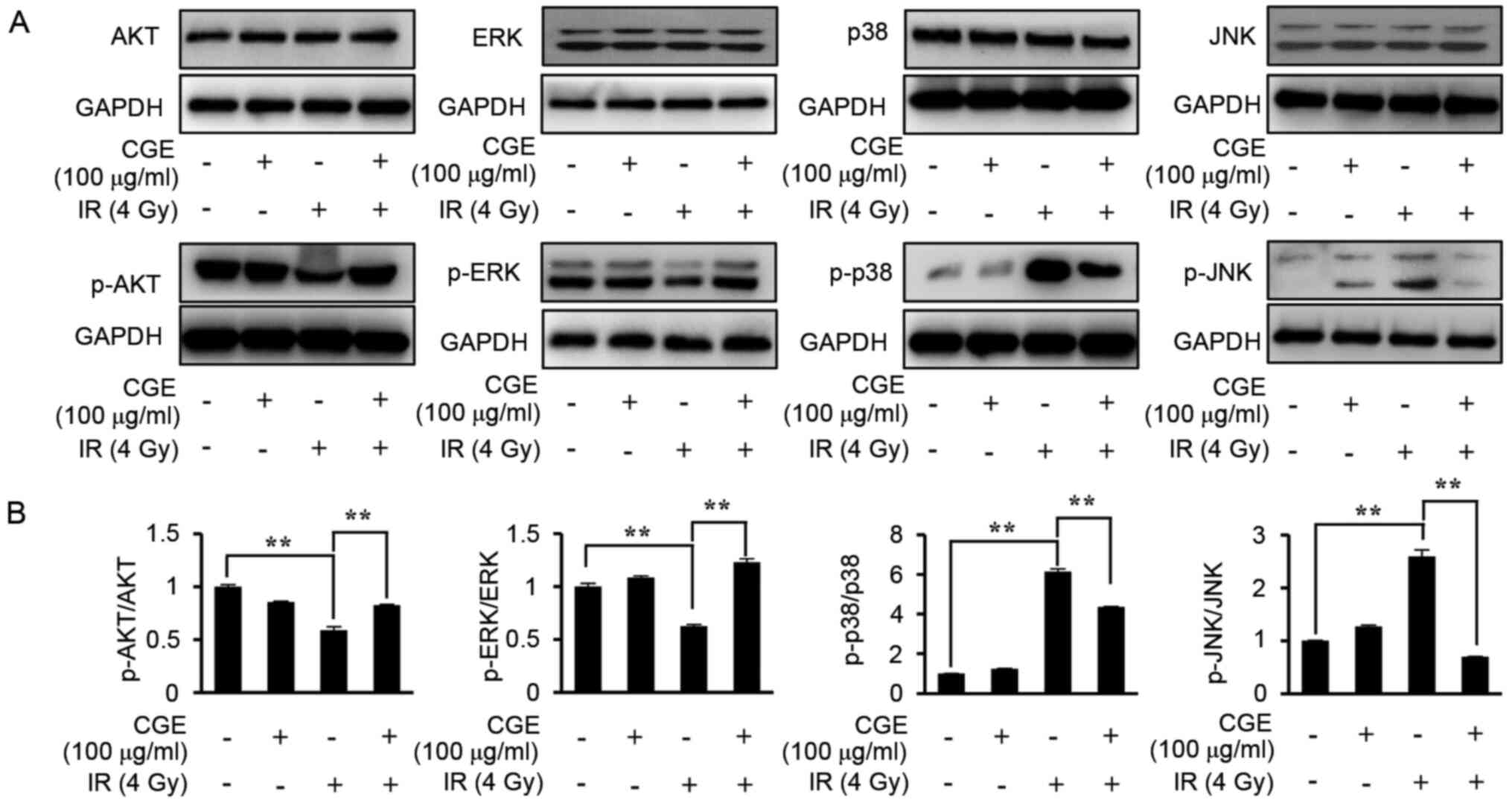 | Figure 6CGE inhibits apoptosis in IR-exposed
NIH-3T3 cells by regulating the ERK-, p38-, JNK-MAPKs signaling.
(A) Lysates of IR-exposed cells in the presence or absence of CGE
were immunoblotted with anti-AKT, anti-p-AKT, anti-ERK, anti-p-ERK,
anti-p38, anti-p-p38, anti-JNK, anti-p-JNK, or anti-GAPDH
antibodies. (B) Western blots were analyzed quantitatively. The
band intensities were normalized to those of the normal form of
each protein or GAPDH. **P≤0.01 vs. IR only. CGE,
centipedegrass extract; IR, ionizing radiation; p-, phosphorylated;
ERK, extracellular-signal-regulated kinase; JNK, c-Jun N-terminal
Kinase; MAPK, mitogen-activated protein kinase; AKT, protein kinase
B; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; Con, vehicle
control (0.1% DMSO); SD, standard deviation. |
Discussion
ROS plays diverse roles depending on the
concentration present in cells. ROS participate in the molecular
signaling and is essential for homeostasis in normal physiological
levels; however, they function as toxic elements at aberrant levels
and are associated with abnormal cell proliferation (26-28).
IR strongly induces intracellular accumulation of ROS and RNS
(29). Humans are exposed to
natural sources of radiation such as those in water, soil, and
vegetation, as well as those from human-made sources such as X-rays
and medical devices (30). Notably,
IR is used for RT, generally, as part of cancer treatment to
control or kill malignant cells. However, some people undergoing RT
experience dryness, itching, blistering, or peeling. This
reversible effect depends on which part of the body has been
exposed to radiation (31). One of
the most common reversible effects is a skin condition called
radiation dermatitis (32), in
which, the ionizing radiation interacts directly or indirectly with
the target macromolecules or water in cells leading to the
occurrence of oxidizing events that alter cell molecular
composition (29,33). In addition, oxidative damage can get
extended from the target to neighboring non-target bystander cells
through a redox-regulated intercellular proximity mechanism
(34). Ultimately, the RT fails,
resulting in unexpected damages to the cancer as well as normal
cells due to IR-induced oxidative stress.
In a previously published study, maysin derived from
corn silk was reported to increase cell viability under conditions
of oxidative stress via upregulation of a neuronal antioxidative
action (17). Conversely, it was
reported that maysin (C-glycosyl flavone) isolated from the silks
of Zea mays L. has a higher antioxidative activity than
other compounds (rutin, quercetin, luteolin), thus, it has
potential as a powerful antioxidant compound. In this study, the
ability of CGE to rescue fibroblast cells from oxidative
stress-induced apoptosis was demonstrated and the underlying
effects were examined. In general, apoptosis has been recognized as
an indication of cell death induced by oxidative stress followed by
cell senescence (35). One of the
most pivotal molecular pathways that are damaged by oxidative
stress is that involving damage to the DNA (36). Previous studies have reported that
DNA damage mediated by reactive oxygen intermediates causes
enzymatic inactivation through cleavage of PARP, which is an
important step in apoptosis (37).
The function of cleaved PARP is to prevent repair of DNA strand
breaks during apoptotic cell death, which is now widely known as
the key marker of type 1 programmed cell death (38). PARP is cleaved by caspase-3 into two
fragments of 89 and 24 kDa during apoptotic cell death in various
cell lines (39). In this study, we
demonstrated that CGE plays a role in cytoprotective effect on
H2O2-induced cell death in mouse-derived
fibroblasts. Pretreatment of NIH-3T3 cells with CGE (up to 100
µg/ml) before H2O2 treatment significantly
attenuated cell death induced by oxidative stress, as observed by
cell density and viability. CGE significantly inhibited PARP
cleavage and prevented sustenance of DNA damage. Additionally, it
was confirmed that CGE could significantly reduce the number of
Annexin V- and PI-positive cells, indicating that pretreatment with
CGE could significantly alleviate apoptosis induced by oxidative
stress. Taken together, the results of this study suggest that due
to its antioxidant activity, CGE has a cellular protective effect
against oxidative stress-induced apoptosis in NIH-3T3 cells and may
potentially act as a protective agent against IR-induced cell
damage.
Furthermore, we have demonstrated that CGE
pretreatment rescues NIH-3T3 cells subjected to IR exposure.
Exposure to IR induced cell death and increased intracellular ROS
levels; however, CGE pretreatment counteracted the cellular
damages. In published reports, IR has been shown to activate three
MAPKs (ERK1/2, p38, JNK MAPK pathway) in a cell type-dependent
manner (25,40), and it has been shown that the
phosphorylated JNK translocates to the nucleus, phosphorylates
c-Jun (41,42). Phosphorylation of c-Jun leads to the
formation of AP-1, JNK-AP-1 pathway is involved in the increased
expression of pro-apoptotic genes (43). Our results have shown that
pretreatment with CGE decreases the activation of JNK and p38 MAPKs
followed by the apoptosis pathway. Taken together, these results
indicated that CGE has protective effects against IR-induced
apoptotic cell death and the mechanism underlying this effect is
ROS scavenging and JNK-, ERK1/2-, p38-MAPK pathway modulation.
Several trials are being conducted to develop a
radioprotector via the inhibition of p38 and JNK pathways in normal
tissues, and some derivates from natural plants have shown
protective effects against IR-induced ROS stress (44,45).
In the previous study, it was reported that maysin (the major
constituent of centipedegrass) not only plays a role as a ROS
scavenger but also is able to increase the amount of antioxidant
enzymes in a mammalian cell. Notably, in this study, CGE also
exhibited protective effects against overwhelming ROS deposition
induced by H2O2, and reduced the extent of
apoptotic cell death induced by IR via downregulation of MAPK
signaling (ERK, p38, JNK) in fibroblasts.
Therefore, CGE could be considered as an efficient
radioprotector or a radiation palliative remedy, which could help
reduce tissue damage induced by exposure to radiation in patients
or sufferers unintentionally exposed to radiation or undergoing RT.
However, to demonstrate the protective effect of CGE on skin
tissue, experiments conducted with a single cell line derived from
mice may be a limitation. To address this, human-derived skin cell
experiments need to be performed. Additionally, in this study, the
effects of individual constituents of CGE are still not be explored
and should be explored in further studies. Furthermore, for the
clinical application of any compound as a candidate for radiation
protection, it is essential to avoid unacceptable clinical risks;
therefore, absolute certainty about its safety for normal tissues
is required. In laboratory studies, several compounds have been
tested as radioprotectants; however, most did not reach the
clinical stage due to the toxicity and side effects in animal
models. Similarly, in this study, CGE was found to be non-toxic
under normal conditions in fibroblasts; however, in vivo
studies using animals with tumors are needed to investigate whether
CGE has preferential radioprotective action in normal tissues over
tumor tissues.
In conclusion, CGE contains C-glycosyl flavones and
phenolic components, which protected mouse-derived fibroblasts from
IR-induced apoptotic cell death by blocking ROS production and
inhibiting ERK-, p38-, JNK-MAPKs signaling. Although large-scale
animal studies and clinically relevant tests are needed to confirm
the effectiveness of CGE, potential applications of CGE as a useful
radioprotectant may be proposed.
Acknowledgements
Not applicable.
Funding
This work was supported by the Nuclear R&D Program of the
Ministry of Science and ICT, Republic of Korea.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SHK and DHB designed and performed the experiments.
SSL and HWB interpreted the experimental results and drafted the
manuscript. BYC and BSK performed the statistical analysis and
revised the manuscript critically for important intellectual
content. SHK and DHB confirmed the authenticity of all the raw
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hirata M, Nagakura Y, Yuki N, Adachi K,
Fujii R, Koyakumaru T, Ogura S, Moritake H, Watanabe C and Fukuyama
K: Development and establishment of centipede grass (Eremochloa
ophiuroides) in south-western Japan. Trop Grassl. 41:100–112.
2007.
|
|
2
|
Park HJ, Chung BY, Lee MK, Song Y, Lee SS,
Chu GM, Kang SN, Song YM, Kim GS and Cho JH: Centipede grass exerts
anti-adipogenic activity through inhibition of C/EBPβ, C/EBPα, and
PPARγ expression and the AKT signaling pathway in 3T3-L1
adipocytes. BMC Complement Altern Med. 12(230)2012.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Lee EM, Bai HW, Lee SS, Hong SH, Cho JY,
Lee IC and Chung BY: Stress-induced increase in the amounts of
maysin and maysin derivatives in world premium natural compounds
from centipedegrass. Radiat Phys Chem. 81:1055–1058. 2012.
|
|
4
|
Lee EM, Lee SS, Bai H-W, Cho J-Y, Kim TH
and Chung BY: Effect of gamma irradiation on the pigments and the
biological activities of methanolic extracts from leaves of
centipedegrass (Eremochloa ophiuroides Munro). Radiat Phys
Chem. 91:108–113. 2013.
|
|
5
|
Liu J, Wang C, Wang Z, Zhang C, Lu S and
Liu J: The antioxidant and free-radical scavenging activities of
extract and fractions from corn silk (Zea mays L.) and
related flavone glycosides. Food Chem. 126:261–269. 2011.
|
|
6
|
Choi DJ, Kim SL, Choi JW and Park YI:
Neuroprotective effects of corn silk maysin via inhibition of
H2O2-induced apoptotic cell death in SK-N-MC
cells. Life Sci. 109:57–64. 2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Baskar R, Lee KA, Yeo R and Yeoh KW:
Cancer and radiation therapy: Current advances and future
directions. Int J Med Sci. 9:193–199. 2012.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Johung K, Saif MW and Chang BW: Treatment
of locally advanced pancreatic cancer: the role of radiation
therapy. Int J Radiat Oncol Biol Phys. 82:508–518. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Riley PA: Free radicals in biology:
Oxidative stress and the effects of ionizing radiation. Int J
Radiat Biol. 65:27–33. 1994.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Uttara B, Singh AV, Zamboni P and Mahajan
RT: Oxidative stress and neurodegenerative diseases: A review of
upstream and downstream antioxidant therapeutic options. Curr
Neuropharmacol. 7:65–74. 2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chan RJ, Webster J, Chung B, Marquart L,
Ahmed M and Garantziotis S: Prevention and treatment of acute
radiation-induced skin reactions: A systematic review and
meta-analysis of randomized controlled trials. BMC Cancer.
14(53)2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Bolderston A, Cashell A, McQuestion M,
Cardoso M, Summers C and Harris R: A canadian survey of the
management of radiation-induced skin reactions. J Med Imaging
Radiat Sci. 49:164–172. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wei J, Meng L, Hou X, Qu C, Wang B, Xin Y
and Jiang X: Radiation-induced skin reactions: Mechanism and
treatment. Cancer Manag Res. 11:167–177. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Chen J, Zhu Y, Zhang W, Peng X, Zhou J, Li
F, Han B, Liu X, Ou Y and Yu X: Delphinidin induced protective
autophagy via mTOR pathway suppression and AMPK pathway activation
in HER-2 positive breast cancer cells. BMC Cancer.
18(342)2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zhang S and Duan E: Fighting against Skin
Aging: The Way from Bench to Bedside. Cell Transplant. 27:729–738.
2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Panich U, Sittithumcharee G, Rathviboon N
and Jirawatnotai S: Ultraviolet radiation-induced skin aging: The
role of DNA damage and oxidative stress in epidermal stem cell
damage mediated skin aging. Stem Cells Int. 2016:7370642.
2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Kim JH, Jenrow KA and Brown SL: Mechanisms
of radiation-induced normal tissue toxicity and implications for
future clinical trials. Radiat Oncol J. 32:103–115. 2014.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Smith TA, Kirkpatrick DR, Smith S, Smith
TK, Pearson T, Kailasam A, Herrmann KZ, Schubert J and Agrawal DK:
Radioprotective agents to prevent cellular damage due to ionizing
radiation. J Transl Med. 15:232. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Painuli S and Kumar N: Prospects in the
development of natural radioprotective therapeutics with
anti-cancer properties from the plants of Uttarakhand region of
India. J Ayurveda Integr Med. 7:62–68. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Mun GI, Kim S, Choi E, Kim CS and Lee YS:
Pharmacology of natural radioprotectors. Arch Pharm Res.
41:1033–1050. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Badaboina S, Bai HW, Park CH, Jang DM,
Choi BY and Chung BY: Molecular mechanism of apoptosis induction in
skin cancer cells by the centipedegrass extract. BMC Complement
Altern Med. 13(350)2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Bindokas VP, Jordán J, Lee CC and Miller
RJ: Superoxide production in rat hippocampal neurons: Selective
imaging with hydroethidine. J Neurosci. 16:1324–1336.
1996.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Green DR and Kroemer G: The
pathophysiology of mitochondrial cell death. Science. 305:626–629.
2004.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Hata AN, Engelman JA and Faber AC: The
BCL2 family: Key mediators of the apoptotic response to targeted
anticancer therapeutics. Cancer Discov. 5:475–487. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Dent P, Yacoub A, Fisher PB, Hagan MP and
Grant S: MAPK pathways in radiation responses. Oncogene.
22:5885–5896. 2003.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Di Meo S, Reed TT, Venditti P and Victor
VM: Role of ROS and RNS sources in physiological and pathological
conditions. Oxid Med Cell Longev. 2016:1245049. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Schieber M and Chandel NS: ROS function in
redox signaling and oxidative stress. Curr Biol. 24:R453–R462.
2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Lee AY, Choi JM, Lee MH, Lee J, Lee S and
Cho EJ: Protective effects of perilla oil and alpha linolenic acid
on SH-SY5Y neuronal cell death induced by hydrogen peroxide. Nutr
Res Pract. 12:93–100. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Azzam EI, Jay-Gerin JP and Pain D:
Ionizing radiation-induced metabolic oxidative stress and prolonged
cell injury. Cancer Lett. 327:48–60. 2012.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Canadian Nuclear Safety Commission (CNSC):
Types and Sources of Radiation. Ottawa, ON, 2014.
|
|
31
|
Bray FN, Simmons BJ, Wolfson AH and Nouri
K: Acute and chronic cutaneous reactions to ionizing radiation
therapy. Dermatol Ther (Heidelb). 6:185–206. 2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Leventhal J and Young MR: Radiation
dermatitis: Recognition, prevention, and management. Oncology
(Williston Park). 31:885–887, 894-899. 2017.PubMed/NCBI
|
|
33
|
Reisz JA, Bansal N, Qian J, Zhao W and
Furdui CM: Effects of ionizing radiation on biological molecules -
mechanisms of damage and emerging methods of detection. Antioxid
Redox Signal. 21:260–292. 2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Mladenov E, Li F, Zhang L, Klammer H and
Iliakis G: Intercellular communication of DNA damage and oxidative
status underpin bystander effects. Int J Radiat Biol. 94:719–726.
2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Redza-Dutordoir M and Averill-Bates DA:
Activation of apoptosis signalling pathways by reactive oxygen
species. Biochim Biophys Acta. 1863:2977–2992. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Whitaker AM, Schaich MA, Smith MR, Flynn
TS and Freudenthal BD: Base excision repair of oxidative DNA
damage: From mechanism to disease. Front Biosci. 22:1493–1522.
2017.PubMed/NCBI View
Article : Google Scholar
|
|
37
|
Rodríguez-Vargas JM, Ruiz-Magaña MJ,
Ruiz-Ruiz C, Majuelos-Melguizo J, Peralta-Leal A, Rodríguez MI,
Muñoz-Gámez JA, de Almodóvar MR, Siles E, Rivas AL, et al:
ROS-induced DNA damage and PARP-1 are required for optimal
induction of starvation-induced autophagy. Cell Res. 22:1181–1198.
2012.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Ko HL and Ren EC: Functional Aspects of
PARP1 in DNA Repair and Transcription. Biomolecules. 2:524–548.
2012.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Soldani C, Lazzè MC, Bottone MG, Tognon G,
Biggiogera M, Pellicciari CE and Scovassi AI: Poly(ADP-ribose)
polymerase cleavage during apoptosis: When and where? Exp Cell Res.
269:193–201. 2001.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Valerie K, Yacoub A, Hagan MP, Curiel DT,
Fisher PB, Grant S and Dent P: Radiation-induced cell signaling:
Inside-out and outside-in. Mol Cancer Ther. 6:789–801.
2007.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Davis RJ: Signal transduction by the JNK
group of MAP kinases. In: Inflammatory Processes. Springer,
pp13-21, 2000.
|
|
42
|
Chang L and Karin M: Mammalian MAP kinase
signalling cascades. Nature. 410:37–40. 2001.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Fan M and Chambers TC: Role of
mitogen-activated protein kinases in the response of tumor cells to
chemotherapy. Drug Resist Updat. 4:253–267. 2001.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Santabárbara-Ruiz P, López-Santillán M,
Martínez-Rodríguez I, Binagui-Casas A, Pérez L, Milán M, Corominas
M and Serras F: ROS-induced JNK and p38 signaling is required for
unpaired cytokine activation during Drosophila regeneration. PLoS
Genet. 11:e1005595. 2015.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Choi EK, Yeo JS, Park CY, Na H, Lim J, Lee
JE, Hong SW, Park SS, Lim DG and Kwak KH: Inhibition of reactive
oxygen species downregulates the MAPK pathway in rat spinal cord
after limb ischemia reperfusion injury. Int J Surg. 22:74–78.
2015.PubMed/NCBI View Article : Google Scholar
|