Introduction
Lumbar spinal stenosis (LSS) is one of the most
common spinal disorders affecting the elderly (1). Degenerative changes in the posterior
structures of the lumbar spine, such as hypertrophy of the facet
joints and ligamentum flavum (LF), in combination with degenerative
spondylolisthesis, can contribute to the development of LSS
(2). The hypertrophy of the LF
has been described in anatomic studies to be 7- to 8-mm-thick in
patients with central stenosis, as opposed to the usual 4 mm or
less (2). Although it is agreed
that spinal mechanical stress (3)
and secreted cytokines (4) from
the herniated disk accelerate the hypertrophy of the LF, which
contributes to the development of LSS, the detailed underlying
mechanisms are not yet fully understood.
Continuous mechanical stress causes degeneration of
the LF (5,6). Common pathological characteristics
in the degenerated LF are the loss of elastic fibers and tissue
fibrosis, and increased collagen in tissues (6–8).
Mechanical stress increases the production of transforming growth
factor (TGF)-β1 in several cell lines, including LF cells isolated
from surgically resected LF (9,10).
TGF-β1 is a key factor in the pathogenesis of tissue fibrosis
(11) and is abundantly expressed
in hypertrophied degenerative LF tissues from LSS (12–14). TGF-β1 increases collagen
expression in LF cells (15).
These previous studies suggest that TGF-β1 plays an important role
in the hypertrophy of the LF and thus in the pathogenesis of LSS.
However, the molecular mechanisms underling the association between
TGF-β1 and LF hypertrophy, particularly the mechanisms underlying
the TGF-β1-induced increase in collagen expression have not yet
been fully elucidated.
Recently, connective tissue growth factor (CTGF) has
been shown to have an increased expression in hypertrophied lumbar
LF and to be involved in the hypertrophy of the LF (16). CTGF is a pro-fibrotic factor
involved in the fibrotic process, such as cell proliferation,
migration, adhesion and extracellular matrix (ECM) accumulation
(17). CTGF has also been
reported to be involved in the biological activities of TGF-β1. For
example, TGF-β1, in association with CTGF, has been shown to
regulate cell proliferation and the synthesis of ECM components
(16–18). TGF-β1 also induces the mRNA
expression of CTGF in human skin fibroblasts (19). TGF-β1 is also a well-known inducer
of ECM components, such as collagen and fibronectin (20). In the presence of CTGF
neutralizing antibody (NA), the pro-fibrogenic effects of TGF-β1,
such as collagen deposition and anchorage-independent growth are
attenuated in fibroblasts (20).
Additionally, mitogen-activated protein kinases (MAPKs) have been
reported to be involved in the regulation of the expression of CTGF
(21,22). However, whether the expression of
CTGF is regulated by TGF-β1 in LF cells and whether it is involved
in the hypertrophy of the LF though the MAPK pathway remains
unknown.
In this study, we examined the viability of cultured
human LF cells, the roles of TGF-β1/CTGF in the proliferation of LF
cells and LF hypertrophy, as well as the role of the MAPK pathway
in the pathogenesis of LSS by measuring the expression of CTGF and
ECM components (collagen I and collagen III) in TGF-β1-treated LF
cells obtained from LF tissues of patients who treated with
posterior pedicle fixation for lumbar fracture or with a standard
nucleotomy for lumbar disc herniation using the Love method.
Materials and methods
Samples
Specimens from 13 patients, who were treated with
posterior pedicle fixation for lumbar fracture or with a standard
nucleotomy for lumbar disc herniation using the Love method at
Zhujiang Hospital of Southern Medical University, Guangzhou, China,
were collected. Informed consent was obtained from each patient,
and this study was approved by the Ethics Committee of Southern
Medical University.
Cell isolation and culture
The cells were isolated from the LF tissues as
previously described (23,24).
Briefly, the specimens were minced with microdissection scissors
under aseptic conditions and washed extensively with
phosphate-buffered saline (PBS) to remove the blood component. The
minced tissue was digested at 37°C for 60 min with 0.2% type I
collagenase (Sigma-Aldrich, St. Louis, MO, USA) in serum-free
Dulbecco's modified Eagle's medium (DMEM; Gibco, Sydney,
Australia). The collagenase-treated ligament chips were washed with
serum-containing DMEM to inhibit collagenase activity. The cells
were then filtered through a sterile nylon mesh filter (75 µm pore
size), and placed in 35-mm Petri dishes at a density of
approximately 5×104 cells/ml in DMEM supplemented with
10% fetal calf serum (Gibco). The cultures were incubated at 37°C
in a humidified atmosphere, air 95% and CO2 5%. The
medium was changed at 2-day intervals, and the explants were
examined daily for cell outgrowth using an inverted light
microscope (IX83; Olympus, Tokyo, Japan). The outgrown cells were
harvested before confluence and subcultured after trypsinization
with 0.2% trypsin/0.02% ethylenediaminetetraacetic acid (EDTA). The
cells at the third-fifth passage were used for the experiments.
Immunofluorescence staining for collagen I and III, and fibronectin
was used to identify the phenotype of the cultured LF cells.
Viability of cultured LF cells
The viability of the LF cells was evaluated using
the 3-(4,5-dimethylthiazol-2-y1)-2,5-diphen-yltetrazolium bromide)
(MTT) (Sigma-Aldrich) colorimetric assay, that is based on the
reduction of formazan crystals by living cells (25). Briefly, LF cells at passages 1, 3,
and 5 were seeded in 96-well tissue culture plates at
2×104 cells/well and incubated at 37°C under 5%
CO2 for 0, 24, 48 and 72 h. The cells was then washed
with PBS and subsequently incubated in 100 µl of 5 mg/ml MTT
solution (Invitrogen Life Technologies, Carlsbad, CA, USA) for 3 h.
MTT was converted into purple colored formazan in living cells,
which was then solubilized with dimethyl sulfoxide (DMSO)
(Invitrogen) and the absorbance of the solution was measured at 450
nm using a microplate reader (Thermo Plate; Rayto Life and
Analytical Science Co., Ltd., Shenzhen, China).
Effect of TGF-β1/CTGF on LF cells
To examine the biological response of human LF cells
to TGF-β1/CTGF, the LF cell cultures were treated with 3 ng/ml
TGF-β1 or 50 ng/ml CTGF (R&D Systems, Minneapolis, MN, USA) in
the absence or presence of CTGF NAs (1:500) for 24 h. The
proliferation of the LF cells was first examined by MTT assay, and
the mRNA expression levels of CTGF, collagen I and collagen III
were then detected by reverse transcription-quantitative polymerase
chain reaction (RT-qPCR), and the protein expression of CTGF in the
cell lysate was detected by western blot analysis.
Determination of the role of the MAPK
pathway
To investigate the role of the MAPK pathway in the
TGF-β1-induced expression of CTGF, collagen I and collagen III, the
LF cell cultures were treated with various MAPK inhibitors
including, the JNK inhibitor, SP600125 (10 µM), the ERK inhibitor,
PD985059 (10 µM), and the p38 inhibitor, SB203580 (100 µM), (all
from Sigma-Aldrich). The cells were pre-treated with the indicated
MAPK inhibitors for 1 h, and were then treated with 3 ng/ml of
TGF-β1 for 24 h. The mRNA expression levels of CTGF, collagen I and
collagen III were then detected by RT-qPCR.
In addition, to further confirm the role of p38, we
examined the expression of p38 and p-p38 in the cells following
treatment with 3 ng/ml TGF-β1 for 2, 4, and 6 h by
immunofluorescence staining, and examined the protein expression of
p38 and p-p38 in the cells following treatment with 3 ng/ml TGF-β1
for 30 min, 1, 2, and 3 h by western blot analysis.
Validation of the critical role of p38
using p38 siRNA
The involvement of p38 in the TGF-β1-induced
expression of CTGF, collagen I and collagen III was further
examined using the siRNA-mediated knockdown of p38. For
transfection with p38 siRNA or non-targeting negative control
(Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), DharmaFECT
reagent (Invitrogen Life Technologies) was used according to the
manufacturer's instructions. After 24 h, the culture medium was
replaced with fresh supplemented medium, and the cells were
cultivated for an additional 24 h following treatment with 3 ng/ml
TGF-β1 for 6 h. The expression levels of p38 and p-p38 were
detected by immunofluorescence staining, and the transfection
efficiency over time (0, 12, 24, 36 h) was also validated by RT-PCR
(data not shown). The mRNA expression levels of CTGF, collagen I,
and collagen III were detected by RT-qPCR. The protein expression
of CTGF was detected by western blot analysis.
RT-qPCR
Following the afore-mentioned incubation or
treatments, total RNA was extracted from the cells using TRIzol
reagent (Invitrogen Life Technologies). A total of 2 µg of total
RNA was used to synthesize complementary DNA (cDNA) using M-MLV
Reverse Transcriptase (Takara, Dalian, China), and subjected to
RT-qPCR using SYBR-Green real-time master mix (Toyobo, Osaka,
Japan) with the following primers: 5′-GGAGTGGGTGTGTGACGAG-3′
(forward) and 5′-GTCTTCCAGTCGGTAAGCCG-3′ (reverse) for CTGF;
5′-AGATCTGAAGTGTGATGACTCAGG-3′ (forward) and 5′-CAGAT
CACGTCATCGCACAAC-3′ (reverse) for collagen I;
5′-ATGTTCCACGGAAACACTGG-3′ (forward) and
5′-GGAGAGAAGTCGAAGGAATGC-3′ (reverse) for collagen III; CGTGT
TGCAGATCCAGACCA (forward) and GCCAGAATGCAGCCTACAGA and (reverse)
for p38 siRNA; and 5′-ACACCCACTCCTCCACCTTT-3′ (forward) and
5′-TTACTCCTTGGAGGCCATGT-3′ (reverse) for GAPDH. The thermal
treatment was 15 min at 95°C, followed by 35 cycles of 15 sec at
95°C, 30 sec at 60°C, and 30 sec at 72°C. Gene expression was
normalized to the level of GAPDH within each sample using the
relative ΔΔCT method. Gene expression is shown as the expression
relative to the control. The data shown are representative of 3
independent experiments.
Western blot analysis
The LF cells were lysed using SDS lysis buffer
(Beyotime, Shanghai, China) and centrifuged at 14,000 × g for 10
min at 4°C. Equal amounts of proteins were separated by SDS-PAGE on
a 10% gel and then transferred onto a nitrocellulose membrane,
followed by blocking with 5% bovine serum albumin (BSA) for 1 h at
room temperature. The membrane was then incubated overnight at 4°C
with rabbit monoclonal antibody specific to CTGF (1:400; ab6992),
p-p38 (1:1,000; ab47363), or p38 (1:400; ab27986). All antibodies
were purchased from Abcam (Cambridge, MA, USA). Following 3 washes,
the membrane was incubated with anti-rabbit IgG conjugated with
horseradish peroxidase (A0208; Beyotime) for 1 h at room
temperature. Detection was performed with luminal chemiluminescent
systems. Quantitative data were obtained using a computing
densitometer and MultiGauge software version 3.0 (Fuji Photo Film
Co., Ltd., Tokyo, Japan).
Immunofluorescence staining
The LF cells were rapidly washed once with PBS and
fixed with 2% paraformaldehyde, blocked with 2% goat serum (GS;
C0265; Beyotime) for 30 min, stained with rabbit anti-collagen I
(1:100; Ab34710; Abcam), anti-collagen III (1:100; Ab7778; Abcam),
anti-fibronectin (1:100; Ab2413; Abcam), p-p38 (1:100), or anti-p38
(1:200) antibodies, and finally visualized with Alexa Fluor 488
goat anti-rabbit antibody (1:150; #4412; Cell Signaling Technology,
Inc., Danvers, MA, USA) for 30 min at room temperature.
Immunofluorescence staining was imaged using a Zeiss LSM 510 laser
scanning confocal microscope (Zeiss, Jena, Germany) as previously
described (26).
Statistical analysis
All data are expressed as the means ± standard error
of the mean (SEM). One-way analysis of variance (ANOVA) with
Fisher's protected LSD post-hoc test was performed to test the
difference in densitometeric data. Non-parametric one sample
Wilcoxon test was used for the analysis of the results of RT-qPCR.
Each experiment was repeated at least 3 times. The statistical
significance level was defined as P<0.05.
Results
Viability of cultured human lumbar LF
cells
Human lumbar LF cells were isolated from surgical
specimens obtained from 13 patients and cultured.
Immunofluorescence staining was used to identify the cell phenotype
(Fig. 1). The cultured cells had
a typical LF cell phenotype, and uniformly expressed collagen I
collagen and fibronectin in each cell, and very few cells expressed
type III collagen. Cell proliferation assay revealed that there
were no significant differences in cell prolifaration among the LF
cell subcultures (passages 1, 3 and 5) at any experimental time
point (0, 24, 48, or 72 h) (P>0.05; Fig. 2). There was no significant
difference in cell bioactivities between the individual donors
(data not shown). During the 72-h time period, the proliferation of
the LF cells was significantly increased with time in each
subculture, showing a similar viability with the primary cultured
LF cells (Fig. 2).
TGF-β1/CTGF enhances the proliferation of
human lumbar LF cells
Both TGF-β1 and CTGF markedly elevated the
proliferation of the LF cells (Fig.
3). Of note, the effects of TGF-β1 and CTGF on the
proliferation of the LF cells were attenuated by CTGF NA,
suggesting that TGF-β1 associates with CTGF.
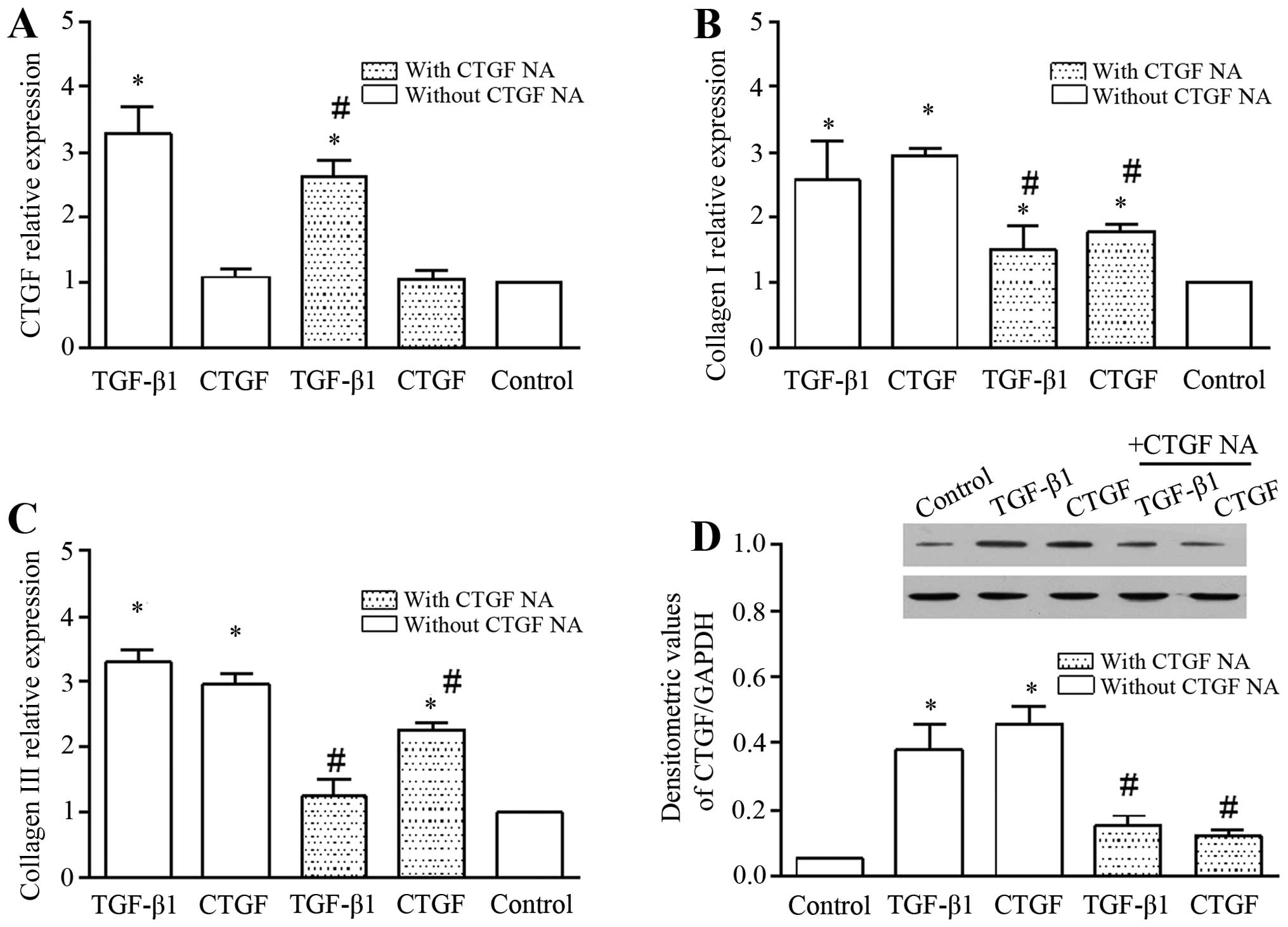
TGF-β1/CTGF induce the mRNA expression of
CTGF, collagen I and collagen III, and the expression of CTGF in
cell lysate
TGF-β1 increased the mRNA expression of CTGF, and
this effect was markedly abolished by CTGF NA (Fig. 4A). To further evaluate the role of
TGF-β1/CTGF in the hypertrophy of the LF, we determined the mRNA
expression of ECM components, such as collagen I (Fig. 4B) and collagen III (Fig. 4C). TGF-β1/CTGF significantly
increased the mRNA expression of collagen I and collagen III. In
addition, the presence of CTGF NA abrogated the promoting effects
of TGF-β1/CTGF on the mRNA expression of collagen I and collagen
III (Fig. 4B and C). We also
examined the expression of CTGF in the cell lysate (Fig. 4D). CTGF expression in the cell
lysate was increased in the presence of TGF-β1 and CTGF, and was
diminished by the addition of CTGF NA. Overall, these findings
suggest that TGF-β1/CTGF plays an important role in the hypertrophy
of the LF.
In addition, it is worthwhile to note that exogenous
CTGF did not enhance the mRNA expression of CTGF in the LF cells
(Fig. 4A), but it increased the
expression of CTGF in the cell lysate (Fig. 4D), indicating that although CTGF
was exogenously expressed in the LF cells, it did not increase the
CTGF mRNA level. Importantly, CTGF NA abolished the promoting
effects of TGF-β1 on CTGF expression.
mRNA expression levels of CTGF, collagen
I and collagen III in TGF-β1-treated cells are mediated by p38, but
not by JNK or ERK
The MAPK inhibitors, namely the JNK inhibitor,
SP600125, and the ERK inhibitor, PD985059, did not influence the
mRNA expression of CTGF (Fig.
5A), collagen I (Fig. 5B) or
collagen III (Fig. 5C). The p38
MAPK inhibitor, SB203580, abolished the promoting effects of TGF-β1
on the mRNA expression of CTGF, collagen I and collagen III,
returning the levels close to the baseline levels (Fig. 5).
Expression levels of p38 and p-p38 in
TGF-β1-treated cells
The immunofluorescence imaging of p38 and p-p38
revealed that their expression and activity was directly related to
the duration of TGF-β1 treatment (Fig. 6A). TGF-β1 gradually and slightly
increased the expression of p38 in the LF cells as time progressed
(P>0.05; Fig. 6B). TGF-β1
gradually elevated the expression of p-p38 as time progressed and
its expression reached a significant level at 1 h (Fig. 6B), showing TGF-β1 activates the
p38 MAPK signaling pathway.
Transfection with p38 siRNA abrogates the
effects of TGF-β1
After silencing the expression of p38 by p38 siRNA
in the LF cells, the mRNA expression levels of CTGF (Fig. 7A), collagen I (Fig. 7B) and collagen III (Fig. 7C) in the presence or absence of 3
ng/ml TGF-β1 for 6 h were significantly decreased compared with the
NC siRNA-transfected cells. Similarly, the silencing of the
expression of p38 in the LF cells significantly diminished the
expression and phosphorylation level of p38, as well as the TGF-β1
(3 ng/ml for 6 h)-induced expression and activity of p38 compared
with the NC siRNA-transfected cells (Fig. 8A). The expression of CTGF in the
cell lysate exhibited a similar trend with its mRNA expression.
TGF-β1 (3 ng/ml for 6 h) increased the expression of CTGF, and this
effect was abrogated by the silencing of p38 (Fig. 8B and C). Overall, the results
presented above indicate that the p38 MAPK signing pathway plays a
critical role in the TGF-β1-induced hypertrophy of the LF.
Discussion
Stenotic LF cells can produce a matrix rich in type
I and III collagen and fibronectin (23), and the cultured cells acquire the
LF cell phenotype with the uniform expression of collagen I and
type III collagen, as well as fibronectin in each cell.
Hypertrophy of the LF plays an important role in the
development of LSS (2). Spinal
mechanical stress (3) and
secreted cytokines (4) from
herniated disk accelerate the hypertrophy of the LF. Mechanical
stress increases the production of TGF-β1 (9,10).
The association between TGF-β1 and LF hypertrophy has not yet been
fully elucidated. In this study, we found that TGF-β1 enhanced the
expression of CTGF at both the mRNA and protein level, further
supporting the existence of an interaction between CTGF and TGF-β1,
as described in a previous study (27). Furthermore, TGF-β1 elevated the
mRNA expression of ECM components, including collagen I and III,
and this effect was abolished by the CTGF NA, indicating that
TGF-β1 contributes to the hypertrophy of the LF in association with
CTGF. Furthermore, the associations between TGF-β1, CTGF and LF
hypertrophy are mediated through the p38 MAPK pathway.
Previous studies have demonstrated that TGF-β1, in
association with CTGF, regulates cell proliferation (16,18). In this study, we found that TGF-β1
enhanced the proliferation of LF cells, and this associated with an
increase in CTGF expression. However, with the addition of CTGF NA,
the promoting effects of TGF-β1 on the proliferation of LF cells
were abrogated (Fig. 3).
Furthermore, TGF-β1, in association with CTGF, regulates the
synthesis of ECM components (16,18). The increased synthesis of collagen
is a major characteristic of LF hypertrophy (10). LF cells have a typical
fibroblast-like phenotype, as they express type I and type III
collagen and fibronectin, but do not stain positive for osteonectin
(23). Normal cells do not
synthesize type II collagen (23). Type I and type III collagen have
been found to be predominant in human LF (28). In this study, we also observed
that TGF-β1 increased the mRNA expression of collagen I and
collagen III, and this effect was abrogated by CTGF NA, indicating
that the TGF-β1-induced synthesis of ECM components is associated
with CTGF. TGF-β1 elevated the expression of CTGF at both the mRNA
and protein level (Fig. 4).
Therefore, TGF-β1, in association with CTGF, contributed to the
hypertrophy of the LF.
In general, the effects of TGF-β are mediated
through the phosphorylation of cytoplasmic R-Smads (29,30). In addition to the activation of
Smad signaling, TGF-β1 can activate members of the MAPK pathway, as
well as other kinases (31,32). Upon stimulation with TGF-β, the
crosstalk between the ERK, p38, JNK and Smad pathways is cell
type-specific (33,34). Previous studies have shown the
pro-fibrotic activities of p38 and ERK signaling, and the
anti-fibrotic activities of JNK signaling (27). The pathway involved in the
TGF-β1-induced hypertrophy of the LF has not been addressed to
date, to the best of our knowledge. In this study, among ERK, p38,
and JNK, only the use of the p38 inhibitor abolished the mRNA
expression of CTGF, and collagen I and III (Fig. 5), indicating the pro-fibrotic
activities of p38 signaling. We further observed that the
expression and phosphorylation level of p38 were enhanced by
TGF-β1. Following the silencing of p38 MAPK (its efficiency over
time was almost the same; data not shown), the expression and
phosphorylation level of p38, as well as the mRNA expression of
CTGF, collagen I and collagen III were attenuated correspondingly,
as well as the expression of CTGF (Fig. 6). Apparently, the expression of
p38 and p-p38 presented in Fig.
6A does not match that shown in Fig. 6B. It is worth noting that the
immunofluorescence staining in this study was focused on the cell
surface, that should limit the detection of intracellular
signaling, but the western blot analysis included all the
intracellular and cell surface information. These results further
confirm that p38 MAPK is a key mediator of the effects of TGF-β1,
and the pro-fibrotic activities of p38 signaling.
In conclusion, TGF-β1, in association with the
increased expression of CTGF, induce the hypertrophy of the LF
through the p38 MAPK pathway. Degenerative changes in the posterior
structures of the lumbar spine, such as the hypertrophy of the
facet joints and LF, in combination with degenerative
spondylolisthesis, contribute to the development of LSS (2). Patients with LSS usually present
with the typical symptoms of neurogenic claudication and/or lumbar
or sacral radiculopathy (35,36). Many patients may also complain of
pain when performing activities requiring the extension of the
spine. These symptoms may improve with appropriate conservative
treatment, although 60–85% of patients undergo surgical treatments
(35). Our observations appear to
be important for evaluating the pathomechanisms of LSS, and may
prove to be helpful in the diagnosis and prevention of LSS in the
early stage, and may also provide an effective alternative to
surgery.
Acknowledgments
This study was supported by funds from the Guangdong
Science and Technology plan project (no. 2011B080701018), and the
Guangdong Medical Science and Technology Research Fund (no.
A2012372).
References
|
1
|
Siebert E, Prüss H, Klingebiel R, Failli
V, Einhäupl KM and Schwab JM: Lumbar spinal stenosis: syndrome,
diagnostics and treatment. Nat Rev Neurol. 5:392–403. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Botwin KP and Gruber RD: Lumbar spinal
stenosis: anatomy and pathogenesis. Phys Med Rehabil Clin N Am.
14:1–15. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fukuyama S, Nakamura T, Ikeda T and Takagi
K: The effect of mechanical stress on hypertrophy of the lumbar
ligamentum flavum. J Spinal Disord. 8:126–130. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Behm B, Babilas P, Landthaler M and
Schreml S: Cytokines, chemokines and growth factors in wound
healing. J Eur Acad Dermatol Venereol. 26:812–820. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Matsumoto Y, Fujiwara T, Imamura R, Okada
Y, Harimaya K, Doi T, Kawaguchi K, Okada S, Yamada Y, Oda Y and
Iwamoto Y: Hematoma of the ligamentum flavum in the thoracic spine:
report of two cases and possible role of the transforming growth
factor beta-vascular endothelial growth factor signaling axis in
its pathogenesis. J Orthop Sci. 18:347–354. 2013. View Article : Google Scholar
|
|
6
|
Maezawa Y, Baba H, Uchida K, Kokubo Y,
Kubota C and Noriki S: Ligamentum flavum hematoma in the thoracic
spine. Clin Imaging. 25:265–267. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sairyo K, Biyani A, Goel VK, Leaman DW,
Booth R Jr, Thomas J, Ebraheim NA, Cowgill IA and Mohan SE: Lumbar
ligamentum flavum hypertrophy is due to accumulation of
inflammation-related scar tissue. Spine. 32:E340–E347. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kosaka H, Sairyo K, Biyani A, Leaman D,
Yeasting R, Higashino K, Sakai T, Katoh S, Sano T, Goel VK and
Yasui N: Pathomechanism of loss of elasticity and hypertrophy of
lumbar ligamentum flavum in elderly patients with lumbar spinal
canal stenosis. Spine. 32:2805–2811. 2007. View Article : Google Scholar
|
|
9
|
Nakamura T, Okada T, Endo M, Kadomatsu T,
Taniwaki T, Sei A, Odagiri H, Masuda T, Fujimoto T, Nakamura T, et
al: Angiopoietin-like protein 2 induced by mechanical stress
accelerates degeneration and hypertrophy of the ligamentum flavum
in lumbar spinal canal stenosis. PLoS One. 9:e855422014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nakatani T, Marui T, Hitora T, Doita M,
Nishida K and Kurosaka M: Mechanical stretching force promotes
collagen synthesis by cultured cells from human ligamentum flavum
via transforming growth factor-beta1. J Orthop Res. 20:1380–1386.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ariel A and Timor O: Hanging in the
balance: endogenous anti-inflammatory mechanisms in tissue repair
and fibrosis. J Pathol. 229:250–263. 2013. View Article : Google Scholar
|
|
12
|
Seko Y, Seko Y, Takahashi N, Shibuya M and
Yazaki Y: Pulsatile stretch stimulates vascular endothelial growth
factor (VEGF) secretion by cultured rat cardiac myocytes. Biochem
Biophys Res Commun. 254:462–465. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Löhr M, Hampl JA, Lee JY, Ernestus RI,
Deckert M and Stenzel W: Hypertrophy of the lumbar ligamentum
flavum is associated with inflammation-related TGF-β expression.
Acta Neurochir (Wien). 153:134–141. 2011. View Article : Google Scholar
|
|
14
|
Park JB, Chang H and Lee JK: Quantitative
analysis of transforming growth factor-beta 1 in ligamentum flavum
of lumbar spinal stenosis and disc herniation. Spine. 26:E492–E495.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen YT, Wei JD, Wang JP, Lee HH, Chiang
ER, Lai HC, Chen LL, Lee YT, Tsai CC, Liu CL, et al: Isolation of
mesenchymal stem cells from human ligamentum flavum: implicating
etiology of ligamentum flavum hypertrophy. Spine. 36:E1193–E1200.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhong ZM, Zha DS, Xiao WD, Wu SH, Wu Q,
Zhang Y, Liu FQ and Chen JT: Hypertrophy of ligamentum flavum in
lumbar spine stenosis associated with the increased expression of
connective tissue growth factor. J Orthop Res. 29:1592–1597. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brigstock DR: The connective tissue growth
factor/cysteine-rich 61/nephroblastoma overexpressed (CCN) family.
Endocr Rev. 20:189–206. 1999.PubMed/NCBI
|
|
18
|
Taipale J, Miyazono K, Heldin CH and
Keski-Oja J: Latent transforming growth factor-beta 1 associates to
fibroblast extracellular matrix via latent TGF-beta binding
protein. J Cell Biol. 124:171–181. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Igarashi A, Okochi H, Bradham DM and
Grotendorst GR: Regulation of connective tissue growth factor gene
expression in human skin fibroblasts and during wound repair. Mol
Biol Cell. 4:637–645. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Abreu JG, Ketpura NI, Reversade B and De
Robertis EM: Connective-tissue growth factor (CTGF) modulates cell
signalling by BMP and TGF-beta. Nat Cell Biol. 4:599–604.
2002.PubMed/NCBI
|
|
21
|
Chen Z, Gibson TB, Robinson F, Silvestro
L, Pearson G, Xu B, Wright A, Vanderbilt C and Cobb MH: MAP
kinases. Chem Rev. 101:2449–2476. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gu J, Liu X, Wang QX, Tan HW, Guo M, Jiang
WF and Zhou L: Angiotensin II increases CTGF expression via
MAPKs/TGF-β1/TRAF6 pathway in atrial fibroblasts. Exp Cell Res.
318:2105–2115. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Specchia N, Pagnotta A, Gigante A,
Logroscino G and Toesca A: Characterization of cultured human
ligamentum flavum cells in lumbar spine stenosis. J Orthop Res.
19:294–300. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhong ZM and Chen JT: Phenotypic
characterization of ligamentum flavum cells from patients with
ossification of ligamentum flavum. Yonsei Med J. 50:375–379. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mosmann T: Rapid colorimetric assay for
cellular growth and survival: application to proliferation and
cytotoxicity assays. J Immunol Methods. 65:55–63. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zeng Y, Adamson RH, Curry FR and Tarbell
JM: Sphingosine-1-phosphate protects endothelial glycocalyx by
inhibiting syndecan-1 shedding. Am J Physiol Heart Circ Physiol.
306:H363–H372. 2014. View Article : Google Scholar :
|
|
27
|
Leask A and Abraham DJ: TGF-beta signaling
and the fibrotic response. FASEB J. 18:816–827. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yoshida M, Shima K, Taniguchi Y, Tamaki T
and Tanaka T: Hypertrophied ligamentum flavum in lumbar spinal
canal stenosis. Pathogenesis and morphologic and
immunohistochemical observation. Spine. 17:1353–1360. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Feng XH and Derynck R: Specificity and
versatility in tgf-beta signaling through Smads. Annu Rev Cell Dev
Biol. 21:659–693. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Massagué J: How cells read TGF-beta
signals. Nat Rev Mol Cell Biol. 1:169–178. 2000. View Article : Google Scholar
|
|
31
|
Strand DW, Liang YY, Yang F, Barron DA,
Ressler SJ, Schauer IG, Feng XH and Rowley DR: TGF-β induction of
FGF-2 expression in stromal cells requires integrated smad3 and
MAPK pathways. Am J Clin Exp Urol. 2:239–248. 2014.
|
|
32
|
Demagny H, Araki T and De Robertis EM: The
tumor suppressor Smad4/DPC4 is regulated by phosphorylations that
integrate FGF, Wnt, and TGF-β signaling. Cell Reports. 9:688–700.
2014. View Article : Google Scholar
|
|
33
|
Derynck R and Zhang YE: Smad-dependent and
Smad-independent pathways in TGF-beta family signalling. Nature.
425:577–584. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Javelaud D and Mauviel A: Crosstalk
mechanisms between the mitogen-activated protein kinase pathways
and Smad signaling downstream of TGF-beta: implications for
carcinogenesis. Oncogene. 24:5742–5750. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Binder DK, Schmidt MH and Weinstein PR:
Lumbar spinal stenosis. Semin Neurol. 22:157–166. 2002. View Article : Google Scholar
|
|
36
|
Omidi-Kashani F, Hasankhani EG and
Ashjazadeh A: Lumbar spinal stenosis: who should be fused? an
updated review. Asian Spine J. 8:521–530. 2014. View Article : Google Scholar : PubMed/NCBI
|