Introduction
Sumoylation, one mode of post-translational
modification for proteins, has been shown to play an important role
in the DNA damage repair, genome maintenance, protein
transportation, activity and stability of proteins (1,2). The
process includes three steps that attach SUMO to other proteins by
activating, conjugating and ligating of enzymes. Among them,
ubiquitin-conjugating protein 9 (Ubc9) is the sole SUMO-conjugating
enzyme. Accumulating evidence has shown that it participates in
many physiological functions, such as oxidative stress, genome
integrity, protein quality control in cardiomocytes and regulation
of meiotic synapsis (3–7).
Ubc9 is also closely related to tumorigenesis and
neoplastic metastasis, suggesting a molecular biomarker of tumors
(8). It is found to highly express
in various cancers, such as colon, lung, head and neck carcinoma,
melanoma, and breast cancers (9–13).
Ubc9 overexpression is greatly associated with neoplastic grades of
breast cancer (14,15). Transfection of the Ubc9
dominant-negative mutant into the human breast MCF-7 tumor cells in
nude mice led to inhibition of tumor growth in vivo
(9). The mechanism underlying this
action can increase the expression of Daxx, a protein that mediates
Fas-associated apoptosis in the cytoplasm (16). Furthermore, compared with wild-type
cells, the expression of bcl-2 proto-oncogene significantly
decreased in the cells transfected with dominant-negative mutant of
Ubc9 (17).
Ubc9 overexpression involves drug resistance to
chemotherapy agents in tumors. It predicts chemoresistance in
breast cancer (18). The
sensitivity to cisplatin, paclitaxel and temozolomide has been
reported increased by knockdown of Ubc9 with specific siRNA in
melanoma (10). It can also
increase the sensitivity of cells to topoisomerase I inhibitor
topotecan and topoisomerase II inhibitor MV-26 (16). Because topoisomerase I
ubiqutination and sumoylation share the same amino acid position,
the sumoylation can prevent the degradation of topoisomerase I
through ubiqutination pathway, which is caused by topoisomerase I
inhibitor, and then decrease the cytotoxicity of topoisomerase I
inhibitors (19). In addition,
SUMO-Ubc9 complexes can modify multiple proteins related with DNA
damage, promote DNA damage repair, and lead to the reduction of
sensitivity to some DNA damage targeting drugs in tumor cells
(20).
Although there are several reports on the
Ubc9-mediated resistance to antitumor drugs, it remains unclear how
it affects different drugs with various actions on tumor cells. It
requires to be further clarified whether Ubc9 is one of the
biomarkers for the prediction of individual tumor response to drug
treatment in clinic. Ubc9 can bind several proteins in vivo,
such as bleomycin hydrolase (BLH) and poly(ADP-ribose) polymerase-1
(PARP-1) (21,22), which are associated with drug
resistance. BLH can deactivate antitumor antibiotic bleomycin
(23) and was confirmed as one of
the biomarkers for determination of bleomycin action in our
previous study (24). PARP-1 is a
critical enzyme in DNA damage repair (25). Its targeted inhibitors, such as
olaparib, have been proved as very effective therapy for advanced
ovarian cancers with BRCA1/BRCA2 mutation (26,27).
Here, we provide the evidence on the sensitivity to
nine antitumor drugs after knockdown of Ubc9 in human breast
carcinoma MCF-7 cells. Our findings reveal that Ubc9-mediated
resistance to them involves BLH and PARP-1.
Materials and methods
Drugs and chemicals
Boningmycin (BON) was kindly provided by Professor
Ruxian Chen at our institute, and its purity was >95%. It was
prepared into a 2-mM solution dissolved in PBS buffer and stored at
−20°C before use. Hydroxycamptothecine (HCPT),
cis-dichlorodiamineplatinum (DDP), 5-fluorouracil (5-FU),
doxorubicin (DOX), vepeside (VP-16), vincristine (VCR),
norcantharidin (NCTD), 3-(4,
5-dimethy1-2-thiazoyl)-2,5-2H-tetrazolium bromide (MTT) and
S-adenosyl methionine (SAMe) were purchased from Sigma-Aldrich (St.
Louis, MO, USA). Gemcitabine (GEM) hydrochloride was obtained from
Lilly France. Unless otherwise stated, all other chemicals were
obtained from Beijing Chemical Factory.
Cell lines and cell culture
Human breast cancer MCF-7 cells were cultured in
RPMI-1640 medium (Hyclone, UT, USA). The media were supplemented
with 10% (v/v) fetal bovine serum (Thermo Fisher Scientific, CA,
USA). The cells were incubated at 37°C in humidified 5%
CO2.
Measurement of cellular viability
assessed using MTT method (28)
In brief, the cells were seeded into a 96-well plate
at a cell density of 3,000 per well for 24 h, followed by drug
treatment for 48 h. Consequently, MTT was added to the medium and
incubated for 2 h, and the crystals were dissolved with dimethyl
sulfoxide. The plates were read using an enzyme-linked
immunosorbent assay plate reader at 570 nm. The control group was
set as 100%, and IC50 values were calculated and plotted
with GraphPad Prism 5.
RNA interference siRNAs duplex against Ubc9 (sense
sequence: GCCUGUCCAUCUUAGAGGAGGACAA), BLH (sense sequence:
UACCCAAACAGAUGCACACCACUCG) was synthesized by Life Technologies
(Carlsbad, CA, USA). The siRNAs duplex against PARP-1 was purchased
from Cell Signaling Technology (Beverly, MA, USA). The RNA
interference with the concentration of 100 pmol RNAi duplex was
performed with Lipofectamine 2000 (Life Technologies) according to
the manufacturer's instructions.
Western blot analysis
Western blot assay was described previously
(28). The sources of primary
antibodies against Ubc9, caspase-3 and PARP-1 were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA). The antibodies
against p53 and actin were from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). The mouse anti-BLH antibody was from Abcam
(Cambridge, UK).
Cell cycle analysis by flow
cytometry
The cells were trypsinized and fixed with cold 70%
ethanol overnight. The fixed cells were washed twice with PBS and
incubated with 100 µg/ml of ribonuclease A at 37°C for 30
min and then stained in PBS containing 50 µg/ml propidium
iodide (Sigma) for 1 h. The fluorescence intensity was detected
using BD FACSCalibur cytometer (BD Biosciences, CA, USA) and the
cell cycle distribution was assayed with the ModFit LT software (BD
Biosciences).
Detection of apoptotic cells by flow
cytometry
The apoptotic cells were stained with Annexin
V-FITC/PI Apoptosis kit (BD Biosciences), following the protocol
provided by the manufacturer. The fluorescence intensities were
measured using a BD FACSCalibur flow cytometer.
The chromosome condensation was stained with Hochest
33342 bisbenzimide. To distinguish the cells at mitosis phase from
G2 phase, DNA specific fluorescent dye Hochest 33342 was
used to stain the cells with the concentration of 2 µg/ml at
room temperature for 15 min. Images were captured with a
fluorescence microscope (Olympus, Japan).
Statistical analysis
Results were expressed as the mean ± SD from at
least three independent experiments. Statistical analysis was
performed with the Student's t-test using SPSS 16.0. The criterion
for statistical significance was set at P<0.05. Combination
index was performed using CompuSyn.
Results
Reduction of cell proliferation by
knockdown of Ubc9
To investigate the functionality of Ubc9 in the role
of cell proliferation, we performed RNA interference experiments.
Three different Ubc9-siRNAs were used to knock down Ubc9 mRNA and
analyzed the levels of Ubc9 by western blotting after MCF-7 cells
were transfected with siRNAs for 72 h. Ubc9 protein levels were
obviously reduced <20%, whereas no such reduction was seen with
treatment of negative siRNA (Fig.
1A). The Ubc9 siRNA-1 and siRNA-2 were chose for the following
experiments. The MCF-7 cells were transfected for 24 h, and cell
numbers were counted every day with Beckman Coulter for a week.
Comparing with the control group, the rate of cell proliferation
was significantly decreased in Ubc9 siRNA-treated group (Fig. 1B). In order to further confirm that
the knockdown of UBC9 can affect cell proliferation, cell cycle
distributions in the MCF-7 cells were analyzed by flow cytometry
following knockdown of Ubc9. As shown in Fig. 1C, knockdown of Ubc9 had increased
accumulation of cells at G2/M-phase from 25.2% in
non-transfected cells to 38.7 and 33.7% in transfected cells,
respectively. To distinguish the cells at G2 with M
phase, we observed the staining by the specific DNA dye Hoechst
33342 (Fig. 1D). The chromosome
condensations showed in Ubc9-interfered group, suggesting the
arrest of cells at M phase. It is consistent with previous research
results in yeast cells (29). The
results suggested that Ubc9 is necessary for cell
proliferation.
Increase of susceptibility to antitumor
agents by knockdown of Ubc9
In order to evaluate whether Ubc9 is related to the
susceptibility to antitumor agents, nine types of antitumor agents
with different actions were chosen to treat MCF-7 cells. As
illustrated in Fig. 2, the
susceptibility to BON, HCPT, DDP, GEM, 5-Fu and VP-16 increased
after knockdown of Ubc9. In contrast, the sensitivity to VCR, NCTD,
or DOX did not change obviously. The IC50 values of
negative siRNA and Ubc9 siRNA are summarized in Table I. Among them, the sensitivity to
BON is mostly augmented, reaching 4-fold. The results revealed that
Ubc9 is involved in the susceptibility to various types of
antitumor agents.
 | Table IPotentiation of actions of antitumor
drugs after knockdown of Ubc9 in MCF-7 cells. |
Table I
Potentiation of actions of antitumor
drugs after knockdown of Ubc9 in MCF-7 cells.
| Drugs | Negative-siRNA | Ubc9-siRNA |
|---|
| BON
(µM) | 5.6±1.1 | 1.3±0.4b |
| HCPT
(µM) | 3.7±0.8 | 1.5±0.4b |
| DDP
(µM) | 12.6±1.6 | 4.5±1.1b |
| GEM
(µg/ml) | 46.3±7.1 | 20.0±3.2b |
| 5-Fu
(µM) | 23.9±3.6 | 10.4±4.3a |
| DOX
(µM) | 0.2±0.1 | 0.2±0.1 |
| VP-16
(µM) | 14.3±2.0 | 6.7±1.8a |
| VCR (nM) | 6.1±1.4 | 4.1±0.8a |
| NCTD
(µM) | 9.7±2.4 | 11.6±2.3 |
Increase of apoptosis by knockdown of
Ubc9
In order to clarify the mechanism by which Ubc9
protein level affects the sensitivity of antitumor agents, the
actions of HCPT and DDP were further observed to detect apoptotic
event with Annexin V/PI staining and western blotting. The rates of
early apoptotic cells were 86.5 and 29.7%, respectively, after the
Ubc9-siRNA transfected cells were exposed to 5 µM HCPT or 10
µM DDP (Fig. 3A), higher in
comparison with the groups of negative siRNA transfected cells. The
apoptotic signaling pathways were also detected by western blotting
at time-points after exposure to HCPT for 6, 12, 24, 36 and 48 h
(Fig. 3B). The cleaved PARP-1
fragment was obviously observed at 24 h in the Ubc9-siRNA
transfected cells. In addition, the protein levels of cleaved
caspase-3 and p53 in Ubc9-siRNA transfected cells significantly
increased comparing with the negative siRNA transfected cells.
Reduction of BLH level in apoptosis was also clearly detected after
knockdown of Ubc9.
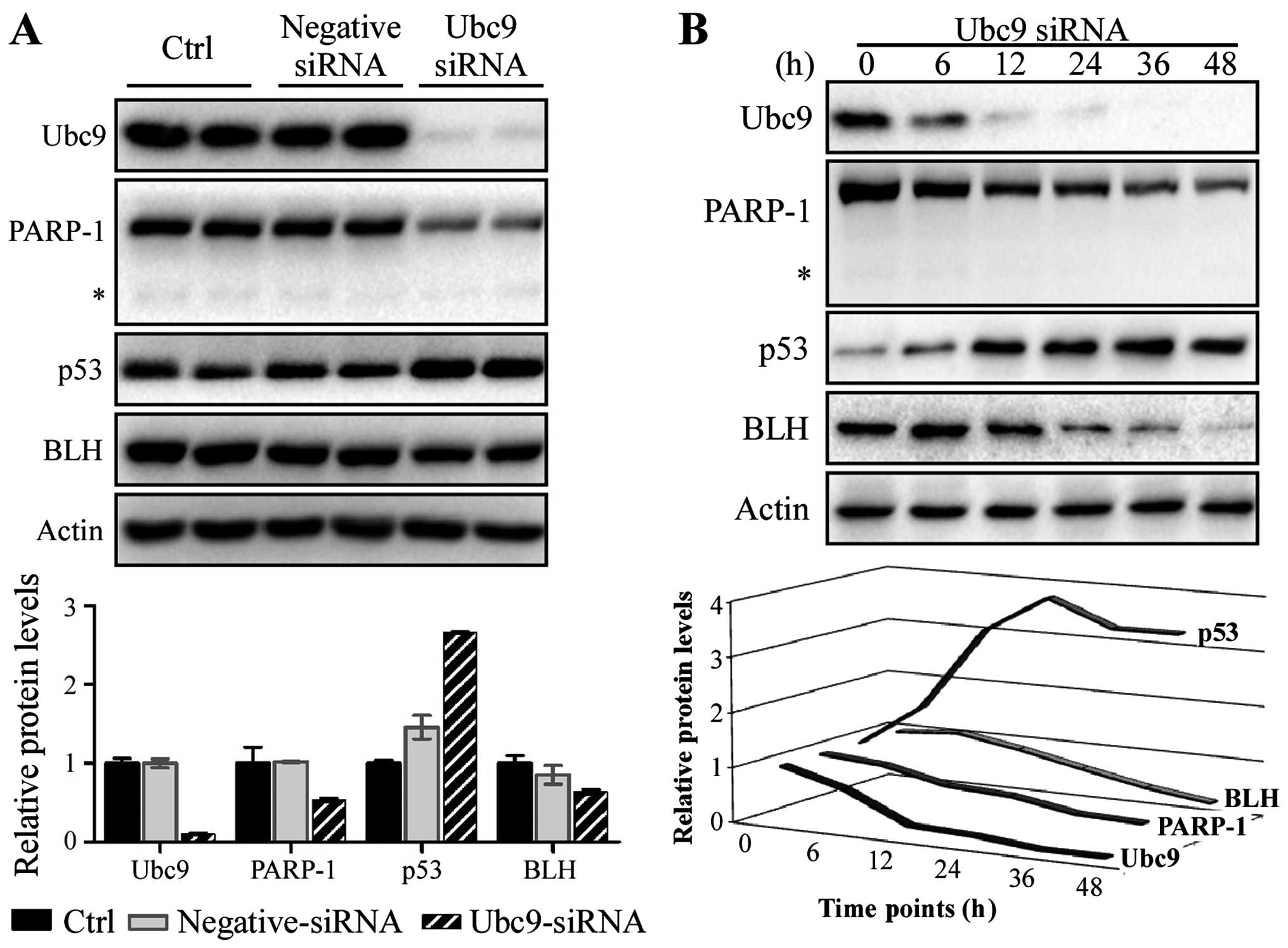
Reduction of BLH and PARP-1 levels in the
Ubc9-siRNA transfected cells
To explore the mechanism of Ubc9-mediated resistance
to antitumor agents, the binding proteins with Ubc9, such as BLH
and PARP-1 were also detected. The protein levels of PARP-1 and BLH
in MCF-7 cells were greatly reduced after the knocking down of Ubc9
protein level (Fig. 4A). Similar
phenomena were also observed time-dependently (Fig. 4B). It is not due to apoptotic event
as the fragment of cleaved PARP-1 was not detected. Interestingly,
increase of p53 protein level was significantly observed after
reduction of Ubc9. These results suggest that BLH and PARP-1 are
related to Ubc9 action in the resistance to antitumor agents.
Increase of the sensitivity to antitumor
agents by knockdown of BLH
In our previous study, the susceptibility to
bleomycin was obviously augmented after knockdown of BLH protein
level (24). BLH levels were
maintained at a lower level after the cells were transfected with
BLH siRNAs for 72 h (Fig. 5A). The
antitumor agents, BON, HCPT, DDP and GEM were used to treat the
MCF-7 cells (Fig. 5B). The
IC50 values of negative siRNA and BLH siRNA groups were
2.5±0.3 and 0.5±0.1 µM, respectively, showing the increased
sensitivity to BON as 5-fold after knockdown of BLH. Importantly,
the reduction of BLH protein level was affected the sensitivity to
HCPT, DDP and GEM (Fig. 5B). The
results suggested that BLH meditates the Ubc9-associated resistance
to antitumor agents in addition to direct metabolism of
bleomycin.
Increase of the sensitivity to antitumor
agents by knockdown of PARP-1
The PARP-1 protein level was maintained at a lower
level after the cells were transfected with PARP-1 siRNAs for 72 h
(Fig. 6A). At the same time, the
sensitivity to BON, HCPT, DDP and GEM were determined (Fig. 6B). The IC50 values of
BON in the negative siRNA and PARP-1 siRNA groups were 3.1±0.3 and
1.6±0.2 µM, respectively, indicating increase of the
sensitivity to BON after knockdown of PARP-1. Interference on
PARP-1 led to 4.1-fold increase of the sensitivity to HCPT. To our
surprise, the sensitivity to DDP and GEM did not change after
reduction of PARP-1 protein level. Thus, PARP-1 partly contributed
to Ubc9-meditated resistance to antitumor agents.
Potentiation of actions of antitumor
agents in combination with Ubc9 inhibitor SAMe
In order to further demonstrate the action of the
Ubc9-mediated drug resistance, Ubc9 inhibitor SAMe was used to
treat MCF-7 cells. It is a common substrate involved in methyl
group transfers and reduction of Ubc9 expression (30). Treatment with SAMe for 48 h led to
reduction of protein levels of Ubc9 and p53 (Fig. 7A). However, the PARP-1 and BLH
protein levels were less reduced after treatment with non-cytotoxic
concentrations of 0.1 and 0.2 mM SAMe. The combination results of
SAMe and antitumor drugs are shown in Fig. 7B. The survival rate had no
significant difference between BON alone and the combined group.
The cell survival rates were greatly decreased in the combination
of HCPT or DDP with SAMe.
Discussion
In this study, we present evidence to demonstrate
the role of Ubc9 in drug resistance to different actions of
chemotherapy agents. The underlying mechanism involves Ubc9 binding
proteins BLH and PARP-1. These results further support the
hypothesis that Ubc9 overexpression is a biomarker for the
prediction of tumor progression and drug resistance.
In order to systemically illuminate the
Ubc9-mediated drug resistance, we assayed the susceptibility to
nine antitumor agents with different actions. BON, a new member of
bleomycin family, showed more potent suppression of human hepatoma
growth in a mouse model (31).
HCPT and VP-16 are topoisomerase I and topoisomerase II inhibitors,
respectively. DOX and DDP act directly blocking DNA replication and
transcription. 5-Fu and GEM inhibited thymidylate synthase and
ribonucleotide reductase, respectively, which were the key enzyme
in DNA synthesis. VCR inhibits mitosis by suppressing microtubule
association. NCTD increase death rate of cancer cells and activity
of lysosomes. In this study, knockdown of Ubc9 protein level led to
different increase of susceptibility to the above-mentioned drugs.
The increases of susceptibility to HCPT and DDP have been observed
after knowdown of Ubc9 or treatment with Ubc 9 specific inhibitor
in MCF-7 cells. It is consistent with the result in melanoma cells
that cisplatin and paclitaxel augmented the rates of Ubc9-related
apoptosis by as much as 50%, but temozolomide by only 10–15%
(10).
Reduction of Ubc9 augments the sensitivity to BON,
5-FU, VP-16 and GEM, but showed no action with DOX. No association
of Ubc9-mediated resistance with DOX is consistent with the DOX
resistance in sumolyation systems of yeast cells (32). It may be the reason why DOX is
widely used for first line tumor therapy in clinic as Ubc9
overexpression in tumor cells has no effect on DOX action. Although
DOX, BON, DDP and VP-16 can cause DNA damage response, the roles of
Ubc9 in the process are totally different. Another report revealed
that depletion of Ubc9 protein level did not affect the homologous
recombination or alternative non-homologous end joining, but
required conservative non-homologous end joining in DNA
double-strand break response (33).
To our knowledge, this is the first report on
knockdown of BLH increasing the sensitivity to antitumor drugs
other than bleomycin in MCF-7 cells (Fig. 5). BLH can inactivate bleomycin
action and is one of the biomarkers for the determination of
bleomycin action (24).
Accumulating data have shown that BLH can play multiple roles under
different physiological and pathological conditions, such as
preparation of peptides for antigen presentation (34), the pathogenesis of Alzheimer's
disease (35) and skin moisture
(36). Furthermore, BLH protects
mice against L-homocysteine thiolactone toxicity by metabolizing it
to homocysteine, suggesting a mechanism by which it has a role in
cellular detoxification (37,38).
In our previous report, we found that BLH is cleaved by caspase-3
in the process of apoptosis (28).
In this study, we demonstrated that knockdown of Ubc9 expression
can cause BLH content to decrease (Fig. 4), and knockdown of BLH protein
level directly affected the sensitivity to several antitumor agents
in MCF-7 cells (Fig. 6). These
findings indicate that BLH may act as a protective role in cells
that degrades intracellular toxic substances and maintains cell
survival. The function of Ubc9/BLH protein complex in the cells is
being investigated in our laboratory.
PARP-1 plays very important roles in DNA damage
repairs, especially in the repair of single-strand DNA breaks. The
results that reduction of PARP-1 protein level after knocking down
of Ubc9 levels and increase of the sensitivity to antitumor drugs
by knockdown of PARP-1 suggest its role in the Ubc9-mediated drug
resistance. Modification of PARP-1 with the small ubiquitin-related
modifier affects its function as a transcriptional co-activator of
hypoxia-responsive genes (39).
The affinity of PARP-1 is enhanced by Ubc9 upon binding to DNA
(40). The PARP-1-targeted agents
have been approved for use in clinic as tumor therapy. This
encourages development of new PARP-1 inhibitors (41). Our results presented here suggest
that wide applications of them should be considered based on the
effects of cellular interacting proteins, such as Ubc9.
It is a very important to understand the way in
which Ubc9 binds BLH and PARP-1 in the cells. There is no evidence
to show their binding is in a competitive manner. As a matter of
fact, only one percent of sumoylated Ubc9 can function well in
yeast meiosis (7), suggesting the
high efficiency to exact its role. It has been observed that
distinct Ubc9 protein complexes formed in response to DNA
double-strand breaks (33).
Therefore, it will be important to determine whether the levels or
activities of these protein complexes are regulated in response to
drug treatment, thereby altering cellular processes through global
or local changes in SUMO modification and binding proteins.
The high expression of Ubc9 in tumor tissues may be
a driving force for tumorigenesis and metastasis as it regulates
the function of many growth-associated oncoproteins. In K-ras
mutant colorectal cancer cells, oncogenesis by Ras/Raf pathway
required Ubc9-mediated sumoylation (42). The phosphorylation of Ubc9 and
SUMO-1 by AKT modulates the substrate sumoylation specificity in
tumor cells (43). The expression
of Ubc9 is regulated by estrogen receptor α and nuclear factor Y in
MCF-7 cells (44). Because of the
complexity and difference of Ubc9 regulation in various tumors, it
will be conducive to the prediction of individual tumor response to
drug treatment in clinic when using Ubc9 as one of the biomarkers.
It is rational to design the compound to target Ubc9 as it is
overexpressed in arrays of tumors and a unique conjugating enzyme
for somoylation. In this study, SAMe inhibited the expression of
Ubc9 and augmented the sensitivity to HCPT and DDP (Fig. 7). Another report showed that
antibiotic spectomycin B1 can directly inhibit Ubc9 in vitro
and in vivo (45).
Developing Ubc9-targeted inhibitors for treatment of various types
of cancers shows promise.
In conclusion, Ubc9 overexpression leads to
resistance of antitumor agents and failure in tumor chemotherapy.
The evidence that BLH and PARP-1 as binding proteins participate in
the process provides new way to overcome this. Ubc9 is a biomarker
for tumorigenesis and the progression in some types of cancer. It
is valuable to precisely detect Ubc9 levels in tumor cells and
screen the compound targeting Ubc9.
Acknowledgments
This study was supported by grants from National
Scientific Foundation of China (81273553, 31471150).
References
|
1
|
Gareau JR and Lima CD: The SUMO pathway:
Emerging mechanisms that shape specificity, conjugation and
recognition. Nat Rev Mol Cell Biol. 11:861–871. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schwertman P, Bekker-Jensen S and Mailand
N: Regulation of DNA double-strand break repair by ubiquitin and
ubiquitin-like modifiers. Nat Rev Mol Cell Biol. 17:379–394. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stankovic-Valentin N, Drzewicka K, König
C, Schiebel E and Melchior F: Redox regulation of SUMO enzymes is
required for ATM activity and survival in oxidative stress. EMBO J.
35:1312–1329. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guervilly JH, Takedachi A, Naim V,
Scaglione S, Chawhan C, Lovera Y, Despras E, Kuraoka I, Kannouche
P, Rosselli F, et al: The SLX4 complex is a SUMO E3 ligase that
impacts on replication stress outcome and genome stability. Mol
Cell. 57:123–137. 2015. View Article : Google Scholar
|
|
5
|
Ouyang J, Garner E, Hallet A, Nguyen HD,
Rickman KA, Gill G, Smogorzewska A and Zou L: Noncovalent
interactions with SUMO and ubiquitin orchestrate distinct functions
of the SLX4 complex in genome maintenance. Mol Cell. 57:108–122.
2015. View Article : Google Scholar
|
|
6
|
Gupta MK, McLendon PM, Gulick J, James J,
Khalili K and Robbins J: UBC9-mediated sumoylation favorably
impacts cardiac function in compromised hearts. Circ Res.
118:1894–1905. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klug H, Xaver M, Chaugule VK, Koidl S,
Mittler G, Klein F and Pichler A: Ubc9 sumoylation controls SUMO
chain formation and meiotic synapsis in Saccharomyces cerevisiae.
Mol Cell. 50:625–636. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mattoscio D and Chiocca S: SUMO pathway
components as possible cancer biomarkers. Future Oncol.
11:1599–1610. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mo YY, Yu Y, Theodosiou E, Ee PL and Beck
WT: A role for Ubc9 in tumorigenesis. Oncogene. 24:2677–2683. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moschos SJ, Smith AP, Mandic M,
Athanassiou C, Watson-Hurst K, Jukic DM, Edington HD, Kirkwood JM
and Becker D: SAGE and antibody array analysis of
melanoma-infiltrated lymph nodes: Identification of Ubc9 as an
important molecule in advanced-stage melanomas. Oncogene.
26:4216–4225. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ronen O, Malone JP, Kay P, Bivens C, Hall
K, Paruchuri LP, Mo YY, Robbins KT and Ran S: Expression of a novel
marker, Ubc9, in squamous cell carcinoma of the head and neck. Head
Neck. 31:845–855. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Moschos SJ, Jukic DM, Athanassiou C,
Bhargava R, Dacic S, Wang X, Kuan SF, Fayewicz SL, Galambos C,
Acquafondata M, et al: Expression analysis of Ubc9, the single
small ubiquitin-like modifier (SUMO) E2 conjugating enzyme, in
normal and malignant tissues. Hum Pathol. 41:1286–1298. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li H, Niu H, Peng Y, Wang J and He P: Ubc9
promotes invasion and metastasis of lung cancer cells. Oncol Rep.
29:1588–1594. 2013.PubMed/NCBI
|
|
14
|
Synowiec E, Krupa R, Morawiec Z, Wasylecka
M, Dziki L, Morawiec J, Blasiak J and Wozniak K: Efficacy of DNA
double-strand breaks repair in breast cancer is decreased in
carriers of the variant allele of the UBC9 gene c. 73G>A
polymorphism. Mutat Res. 694:31–38. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhu S, Sachdeva M, Wu F, Lu Z and Mo YY:
Ubc9 promotes breast cell invasion and metastasis in a
sumoylation-independent manner. Oncogene. 29:1763–1772. 2010.
View Article : Google Scholar :
|
|
16
|
Mo YY, Yu Y, Ee PL and Beck WT:
Overexpression of a dominant-negative mutant Ubc9 is associated
with increased sensitivity to anticancer drugs. Cancer Res.
64:2793–2798. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lu Z, Wu H and Mo YY: Regulation of bcl-2
expression by Ubc9. Exp Cell Res. 312:1865–1875. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen SF, Gong C, Luo M, Yao HR, Zeng YJ
and Su FX: Ubc9 expression predicts chemoresistance in breast
cancer. Chin J Cancer. 30:638–644. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mo YY, Yu Y, Shen Z and Beck WT: Nucleolar
delocalization of human topoisomerase I in response to topotecan
correlates with sumoylation of the protein. J Biol Chem.
277:2958–2964. 2002. View Article : Google Scholar
|
|
20
|
Han JY, Lee GK, Yoo SY, Yoon SJ, Cho EY,
Kim HT and Lee JS: Association of SUMO1 and UBC9 genotypes with
tumor response in non-small-cell lung cancer treated with
irinotecan-based chemotherapy. Pharmacogenomics J. 10:86–93. 2010.
View Article : Google Scholar
|
|
21
|
Masson M, Menissier-de Murcia J, Mattei
MG, de Murcia G and Niedergang CP: Poly(ADP-ribose) polymerase
interacts with a novel human ubiquitin conjugating enzyme: hUbc9.
Gene. 190:287–296. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Koldamova RP, Lefterov IM, DiSabella MT
and Lazo JS: An evolutionarily conserved cysteine protease, human
bleomycin hydrolase, binds to the human homologue of
ubiquitin-conjugating enzyme 9. Mol Pharmacol. 54:954–961.
1998.PubMed/NCBI
|
|
23
|
Schwartz DR, Homanics GE, Hoyt DG, Klein
E, Abernethy J and Lazo JS: The neutral cysteine protease bleomycin
hydrolase is essential for epidermal integrity and bleomycin
resistance. Proc Natl Acad Sci USA. 96:4680–4685. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen J, Chen Y and He Q: Action of
bleomycin is affected by bleomycin hydrolase but not by caveolin-1.
Int J Oncol. 41:2245–2252. 2012.PubMed/NCBI
|
|
25
|
Pellegrino S and Altmeyer M: Interplay
between ubiquitin, SUMO, and poly(ADP-Ribose) in the cellular
response to genotoxic stress. Front Genet. 7:632016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ledermann J, Harter P, Gourley C,
Friedlander M, Vergote I, Rustin G, Scott C, Meier W,
Shapira-Frommer R, Safra T, et al: Olaparib maintenance therapy in
platinum-sensitive relapsed ovarian cancer. N Engl J Med.
366:1382–1392. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ledermann J, Harter P, Gourley C,
Friedlander M, Vergote I, Rustin G, Scott CL, Meier W,
Shapira-Frommer R, Safra T, et al: Olaparib maintenance therapy in
patients with platinum-sensitive relapsed serous ovarian cancer: A
preplanned retrospective analysis of outcomes by BRCA status in a
randomised phase 2 trial. Lancet Oncol. 15:852–861. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen Y, Xu R, Chen J, Li X and He Q:
Cleavage of bleomycin hydrolase by caspase-3 during apoptosis.
Oncol Rep. 30:939–944. 2013.PubMed/NCBI
|
|
29
|
Dieckhoff P, Bolte M, Sancak Y, Braus GH
and Irniger S: Smt3/SUMO and Ubc9 are required for efficient
APC/C-mediated proteolysis in budding yeast. Mol Microbiol.
51:1375–1387. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tomasi ML, Tomasi I, Ramani K, Pascale RM,
Xu J, Giordano P, Mato JM and Lu SC: S-adenosyl methionine
regulates ubiquitin-conjugating enzyme 9 protein expression and
sumoylation in murine liver and human cancers. Hepatology.
56:982–993. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gao N, Shang B, Zhang X, Shen C, Xu R, Xu
H, Chen R and He Q: Potent antitumor actions of the new antibiotic
boningmycin through induction of apoptosis and cellular senescence.
Anticancer Drugs. 22:166–175. 2011. View Article : Google Scholar
|
|
32
|
Huang RY, Kowalski D, Minderman H, Gandhi
N and Johnson ES: Small ubiquitin-related modifier pathway is a
major determinant of doxorubicin cytotoxicity in Saccharomyces
cerevisiae. Cancer Res. 67:765–772. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu Y and Parvin JD: Small ubiquitin-like
modifier (SUMO) isoforms and conjugation-independent function in
DNA double-strand break repair pathways. J Biol Chem.
289:21289–21295. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Towne CF, York IA, Watkin LB, Lazo JS and
Rock KL: Analysis of the role of bleomycin hydrolase in antigen
presentation and the generation of CD8 T cell responses. J Immunol.
178:6923–6930. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Suszynska J, Tisonczyk J, Lee HG, Smith MA
and Jakubowski H: Reduced homocysteine-thiolactonase activity in
Alzheimer's disease. J Alzheimers Dis. 19:1177–1183.
2010.PubMed/NCBI
|
|
36
|
Son ED, Kim Y, Joo KM, Kim HJ, Lee E, Nam
GW, Cho EG, Noh M, Chung JH, Byun SY, et al: Skin dryness in
apparently healthy human skin is associated with decreased
expression of bleomycin hydrolase in the stratum corneum. Clin Exp
Dermatol. 40:247–253. 2015. View Article : Google Scholar
|
|
37
|
Zimny J, Sikora M, Guranowski A and
Jakubowski H: Protective mechanisms against homocysteine toxicity:
The role of bleomycin hydrolase. J Biol Chem. 281:22485–22492.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Borowczyk K, Tisończyk J and Jakubowski H:
Metabolism and neurotoxicity of homocysteine thiolactone in mice:
Protective role of bleomycin hydrolase. Amino Acids. 43:1339–1348.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Messner S, Schuermann D, Altmeyer M,
Kassner I, Schmidt D, Schär P, Müller S and Hottiger MO:
Sumoylation of poly(ADP-ribose) polymerase 1 inhibits its
acetylation and restrains transcriptional coactivator function.
FASEB J. 23:3978–3989. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zilio N, Williamson CT, Eustermann S, Shah
R, West SC, Neuhaus D and Ulrich HD: DNA-dependent SUMO
modification of PARP-1. DNA Repair (Amst). 12:761–773. 2013.
View Article : Google Scholar
|
|
41
|
Scott CL, Swisher EM and Kaufmann SH: Poly
(ADP-ribose) polymerase inhibitors: Recent advances and future
development. J Clin Oncol. 33:1397–1406. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yu B, Swatkoski S, Holly A, Lee LC, Giroux
V, Lee CS, Hsu D, Smith JL, Yuen G, Yue J, et al: Oncogenesis
driven by the Ras/Raf pathway requires the SUMO E2 ligase Ubc9.
Proc Natl Acad Sci USA. 112:E1724–E1733. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lin CH, Liu SY and Lee EH: SUMO
modification of Akt regulates global SUMOylation and substrate
SUMOylation specificity through Akt phosphorylation of Ubc9 and
SUMO1. Oncogene. 35:595–607. 2016. View Article : Google Scholar
|
|
44
|
Ying S, Dünnebier T, Si J and Hamann U:
Estrogen receptor alpha and nuclear factor Y coordinately regulate
the transcription of the SUMO-conjugating UBC9 gene in MCF-7 breast
cancer cells. PLoS One. 8:e756952013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hirohama M, Kumar A, Fukuda I, Matsuoka S,
Igarashi Y, Saitoh H, Takagi M, Shin-ya K, Honda K, Kondoh Y, et
al: Spectomycin B1 as a novel SUMOylation inhibitor that directly
binds to SUMO E2. ACS Chem Biol. 8:2635–2642. 2013. View Article : Google Scholar : PubMed/NCBI
|