Introduction
Diabetes mellitus (DM) is a global disease.
According to the International Diabetes Federation, the incidence
of diabetes is increasing worldwide, with an incidence of 5.4% in
2013 and 592 million cases occurring within 25 years (1). Approximately 80% of diabetes-related
deaths occur in developing countries (1). Approximately 1/3 of the patients with
diabetes develop diabetic nephropathy (DN). As a type of
microvascular disease, DN is one of the causes of mortality and
disability to patients, and is also the main cause of end-stage
renal disease (2).
Glomerulosclerosis and tubular fibrosis are the main pathological
characteristics of DN, in which glomerular podocyte injury is an
early event (3). As glomerular
visceral epithelial cells, podocytes are highly differentiated
renal parenchymal cells with various characteristics and functions,
including endocytosis, the secretion of bioactive mediators and
cytokines, and are involved in the local immune response of the
kidneys (4-6).
The mononuclear macrophage system includes monocytes
in the blood and macrophages in the tissues. Monocytes are produced
by bone marrow and remain in the blood for 2-3 days before entering
the tissues to further differentiate into mature macrophages, which
have different types and names in various organs and tissues
(7). Recent studies have found
that activated macrophages play a significant role in the
development of DN and are the major immune cells responsible for
renal tissue stromal hyperplasia, glomerular sclerosis and
irreversible pathological changes of glomeruli (8-10).
Mononuclear macrophages have plasticity and pluripotency, can
acquire different phenotypes, and exert distinct immune effects in
different environments in the body (11). Specifically, they can be divided
into two types in terms of their functions, namely, classical M1
macrophages activated by Th1 cytokines or lipopolysaccharides and
Th2 cytokine-derived alternative M2 macrophages (12). M1 and M2 macrophages play opposing
roles in the regulation of inflammation. More specifically, M1
macrophages play important roles in antigen presentation and immune
inflammatory effects, whereas M2 macrophages mainly release
cytokines that inhibit inflammation and exert anti-inflammatory
effects (7,13).
As a member of the Sirtuin family, Sirt6 is located
on chromosome 19p13.3 (14) and
consists of a N-terminal, C-terminal and conserved central domain.
The N-terminus is associated with chromosome binding and catalytic
activity, while the C-terminus is involved in nuclear localization
(15-17). Sirt6 has the activities of
NAD+-dependent deacetylase and ADP-ribonyltransferase;
therefore, it not only regulates the expression of certain genes,
DNA repair and gene stability and the homeostasis of hematopoietic
cells, but also maintains glucose homeostasis in the body and plays
a role in the occurrence of obesity (17-21).
A previous study on the zinc-dependent histone deacetylase revealed
that histone deacetylase 4 (HDAC4) can promote the injury of
podocytes in DN (22). However, to
the best of our knowledge, the roles of class III histone
deacetylases, Sirtuins, in the kidneys have not yet been fully
investigated. In the present study, we thus aimed to explore the
associations among Sirt6, macrophages and DN, and to elucidate the
role of Sirt6 in DN.
Materials and methods
Animal model, tissues samples and
monocyte separation
A total of 36 male Sprague-Dawley (SD) rats were
purchased from the Animal Experimental Center of Guangdong Medical
(Guangdong, China). The rats (weighing 220-250 g) were kept in
cages at room temperature (22±3°C) with a stable humidity (50±10%)
with free access to food/water in a light/dark (12-h/12-h) cycle.
The animal experiments were performed according to the Guide for
the Care and Use of Laboratory Animals and approved by the First
People's Hospital of Yuhang District, Hangzhou Animal Ethics
Committee (Hangzhou, China). A diabetic model was established using
18 rats, which were intraperitoneally injected with 60 mg/kg
streptozotocin over a period of 5 consecutive days and
streptozotocin was dissolved in citrate buffer, as previously
described (23). A total of 18
control rats were intraperitoneal injected with citrate buffer (pH
4.5). At 1, 3 and 5 weeks, blood glucose levels in the rats (n=6 at
each time point) with or without streptozotocin injections were
measured by obtaining tail venous blood. Rats with post-fasting
blood glucose levels >250 mg/dl were defined as diabetic.
The rats (n=6) with or without streptozotocin
injections were sacrificed at 1, 3 and 5 weeks and their kidneys
were removed and cut into slices. The kidney tissues were divided
into two parts; one part was stored in 4% formaldehyde solution for
a routine pathological diagnosis, and the other was stored in −80°C
to help perform the following reverse transcription-quantitative
PCR (RT-qPCR) and western blot analyses.
Fresh renal tissues that were cut into small
sections were transferred to DMEM with collagenase at 37°C for 45
min and filtered through a nylon cell strainer. The digested cells
were added to DMEM (Gibco/Thermo Fisher Scientific, Waltham, MA,
USA) medium and were centrifuged at 1,200 × g for 10 min at 4°C
(23).
Hematoxylin and eosin (H&E) and
immunohistochemical (IHC) staining
Renal tissues specimens were obtained from 4%
paraformaldehyde fixative, and then dehydrated and embedded in
paraffin. The embedded tissues were sliced into 4-μm-thick
sections, which were stained with hematoxylin (Sigma-Aldrich, St.
Louis, MO, USA) for 5 min at room temperature, then stained with
eosin (Sigma-Aldrich) for 3 min at room temperature, and observed
under a DMi8 optical microscope (Leica, Hamburg, Germany) at ×200
magnification.
Sirt6 expression in the renal tissues from the rats
with or without DN were examined by IHC staining. The sections were
dewaxed using two types of xylene solution, hydrated by ethanol at
various gradients and antigen retrieval was achieved by quenching
with endogenous peroxidase. The sections were then washed with PBS
for 3 times and incubated with primary Sirt6 antibodies (1:100;
ab135566, Abcam, Cambridge, MA, USA) at 4°C overnight and then
washed with PBS. The sections were then incubated with secondary
antibody biotin-conjugated goat anti-rabbit IgG (Proteintech Group,
Inc./Thermo Fisher Scientific; SA00004-2; 1:200) at 25°C for 30
min. Subsequently, diaminobenzidine (DAB) was used as a chromogen
and hematoxylin was used to re-dye the sections, at 25°C for 3 min.
The stained sections were examined using a DMi8 optical microscope
(Leica) in randomly selected sections at ×200 magnification.
THP-1 cell culture and macrophage
transformation
Human monocyte THP-1 cells were purchased from the
Shanghai Cell Bank of Chinese Academy of Sciences (Shanghai,
China). The cells were cultured at 37°C with 5% CO2 in
RPMI-1640 medium (GENOM, Hangzhou, China) with 10% fetal bovine
serum (Thermo Fisher Scientific), 1% penicillin/streptomycin
(Gibco/Thermo Fisher Scientific). To form macrophages, 100 ng/ml
phorbol 12-myristate 13-acetate (PMA; Sigma-Aldrich), which is also
named tetradecanoylphorbol 13-acetate (TPA), was added to the THP-1
cells and cultured at 37°C for 48 h. The cells were then washed
with PBS twice and continuously cultured in RPMI-1640 medium.
M1 type macrophages were transformed by glucose and
incubated at 37°C for 24 h. The cells were washed with PBS 3 times
and cultured in normal RPMI-1640 meduma for a further 24 h. The
supernatants were then collected and centrifuged at 2,000 × g at
4°C for 20 min. There were 4 different groups of cells as follows:
The controls (exposed to 5.5 mmol/l glucose), the HG1 group
(exposed to 15 mmol/l glucose), the HG2 group (exposed to 30 mmol/l
glucose) and the HG3 group (exposed to 50 mmol/l glucose).
Podocyte culture
Immortalized mouse podocyte MPC-5 cells were
purchased from the Research Facilities of Peking Union Medical
College (PUMC) Cell Bank (Beijing, China). Undifferentiated MPC-5
cells were cultured in a DMEM (Gibco/Thermo Fisher Scientific) with
10 U/ml mouse recombinant interferon-γ (IFN-γ; Peprotech Inc.,
Rocky Hill, NJ, USA) at 33°C in an incubator with 5%
CO2. Podocytes were cultured at 37°C for 10-14 days in
DMEM without IFN-γ to induce cell differentiation and
maturation.
Cell transfection
Sirt6 overexpression and empty control plasmids were
purchased from Invitrogen/Thermo Fisher Scientific. Macrophages and
MPC-5 cells were respectively seeded in 6-well plates
(1.0×105) for 24 h prior to transfection. Transient
transfection was carried out using Lipofectamine 3000
(Invitrogen/Thermo Fisher Scientific) according to the standard
protocol. A total of 20 μM overexpressing-RNA, control,
negative control (NC) and Lipofectamine 3000 were added to
serum-free medium and incubated at 25°C for 10 min, respectively.
Subsequently, Lipofectamine 3000 was mixed into each group that was
cultured in serum-free DMEM. Following 6 h of culture, the fluid
was replaced by DMEM containing 10% fetal bovine serum (FBS).
Co-culture of macrophages with
podocytes
We used a Transwell to establish a co-culture
system. Macrophages were transfected with control/NC/Sirt6
overexpression plasmid and seeded on a 0.4 μm Transwell
insert (Millipore, Billerica, MA, USA) with RPMI-1640 medium and
then co-cultured with podocytes transfected with control/NC/Sirt6
overexpression plasmids for 48 h in the presence of 50 mmol/l
high-glucose treatment. In this experiment, the cells were divided
into 8 groups as follows: i) The NM + HG + Con group (control; 50
mmol/l of high-glucose medium in the upper chamber, and podocytes
with high-glucose medium in the lower chamber); ii) the HG + Con
group (macrophages with high-glucose medium in the upper chamber,
and podocyte with high-glucose medium in the lower chamber); iii)
the HG + NC + Con group (macrophages transfected with NC plasmid
with high-glucose medium in the upper chamber, and podocytes with
high-glucose medium in the lower chamber); iv) the HG + Sirt6 + Con
group (macrophages transfected with Sirt6 overexpression plasmid
with high-glucose medium in the upper chamber, and podocytes with
high-glucose medium in the lower chamber); v) the M-Sirt6 group
(high-glucose medium in the upper chamber, and podocytes
transfected with Sirt6 overexpression plasmid with high-glucose
medium in the lower chamber); vi) the HG + M-Sirt6 group
(macrophages with high-glucose medium in the upper chamber, and
podocytes transfected with Sirt6 overexpression plasmid with
high-glucose medium in the lower chamber); vii) the HG + NC +
M-Sirt6 group (macrophages transfected with NC plasmid with
high-glucose medium in the upper chamber, and podocytes transfected
with Sirt6 overexpression plasmid with high-glucose medium in the
lower chamber); and viii) the HG + Sirt6 + M-Sirt6 (macrophages
transfected with Sirt6 overexpression plasmid with high-glucose
medium in the upper chamber, and podocytes transfected with Sirt6
overexpression plasmid with high-glucose medium in the lower
chamber). The control group and M-Sirt6 (macrophages with
high-glucose medium in the upper chamber, and podocytes transfected
with Sirt6 overexpression plasmid with high-glucose medium in the
lower chamber) group were not cultured with macrophages.
Flow cytometry
Cell apoptosis was determined by flow cytometry with
the Annexin V-FITC/PI staining kit (BD Biosciences, San Jose, CA,
USA). The cells were washed twice using washing buffer, and the
suspension was cultured with Annexin V and propidium iodide (PI;
Yeasen Biotechnology Co., Ltd., Shanghai, China) in the dark at
25°C for 15 min. Binding buffer was then added into each well. The
samples were analyzed by BD FACSCalibur flow cytometer (BD
Biosciences, San Diego, CA, USA) within 1 h.
RT-qPCR
Sirt6, tumor necrosis factor (TNF)-α, inducible
nitric oxide synthase (iNOS), interleukin (IL)-4, IL-10, CD86,
CD206, Bcl-2 and Bax expression levels were detected by RT-qPCR in
the different groups. Total RNA was obtained from the tissues or
cultured cells using TRIzol reagent (Invitrogen/Thermo Fisher
Scientific) according to the manufacturer's instructions. Reverse
transcription was performed using the PrimeScript™ RT reagent kit
(Takara, Shiga, Japan) at 37°C for 15 min and 85°C for 5 min. cDNA
amplified using SYBR Fast qPCR Mix (Invitrogen/Thermo Fisher
Scientific) and the Sirt6 cycle was set at 95°C for 5 min, followed
by 30 cycles at 95°C for 30 sec, at 58°C for 30 sec and at 72°C for
30 sec. The TNF-α cycle was set at 94 °C for 5 min, followed by 40
cycles at 94°C for 30 sec, at 60°C for 30 sec and at 72°C for 60
sec. The iNOS cycle was set at 94°C for 5 min, followed by 35
cycles at 94°C for 30 sec, at 62°C for 30 sec and at 72°C for 30
sec. The IL-4 cycle was set at 94°C for 4 min, followed by 35
cycles at 94°C for 30 sec, at 54°C for 30 sec and at 72°C for 40
sec. The IL-10 cycle was set at 95°C for 3 min, followed by 45
cycles at 95°C for 15 sec, at 53°C for 30 sec and at 60°C for 30
sec. The Bax, Bcl-2 and GAPDH cycles were set at 95°C for 10 sec,
followed by 40 cycles at 95°C for 30 sec, at 58°C for 15 sec and at
72°C for 15 sec. GAPDH served as an internal control. All primers
were used in the synthesis (Sangon Biotech, Shanghai, China) and
sequences are listed in Table I.
Amplified products were electrophoresed through 2% agarose gels.
The amount of RNA was quantified using the 2−∆∆Ct method
(24).
 | Table IPrimers used in RT-qPCR. |
Table I
Primers used in RT-qPCR.
| Gene | Species | Primer | Sequence |
|---|
| TNF-α | Rat | Forward |
5′-CCAACAAGGAGGAGAAGTTCC-3′ |
| | Reverse |
5′-TCTGCTTGGTGGTTTGCTAC-3′ |
| iNOS | Rat | Forward |
5′-GGTGTTCTTTGCTTCTGTGCTA-3′ |
| | Reverse |
5′-CTTCCGACTTTCCTGTCTCAGT-3′ |
| iNOS | Human | Forward |
5′-CCAGCTAGCCAAAGTCACCAT-3′ |
| | Reverse |
5′-GTCTCGGAGCCATACAGGATT-3′ |
| IL-4 | Rat | Forward |
5′-TTACCCGTCTCATTTGC-3′ |
| | Reverse |
5′-TTCCCACTTTGCTGTTCC-3′ |
| IL-10 | Rat | Forward |
5′-CAGAAATCAAGGAGCATTTG-3′ |
| | Reverse |
5′-CTGCTCCACTGCCTTGCTTT-3′ |
| IL-10 | Human | Forward |
5′-GCCAAGCCTTGTCTGAGATGATCC-3′ |
| | Reverse |
5′-AATCGATGACAGCGCCGTAGC-3′ |
| Sirt6 | Human | Forward |
5′-AATTACGCGGCGGGGCT-3′ |
| | Reverse |
5′-CGCGCGCTCTCAAAGGT-3′ |
| Sirt6 | Mouse | Forward |
5′-GGCTACGTGGATGAGGTGAT-3′ |
| | Reverse |
5′-GGCTCAGCCTTGAGTGCTAC-3′ |
| Bax | Mouse | Forward |
5′-AGCCACAAAGATGGTCACT-3′ |
| | Reverse |
5′-GGAGATGAACTGGATAGCAA-3′ |
| Bcl-2 | Mouse | Forward |
5′-ATCTCCCTGTTGACGCTCT-3′ |
| | Reverse |
5′-CATCTTCTCCTTCCAGCCT-3′ |
| GAPDH | Human | Forward |
5′-ACCACAGTCCATGCCATCAC-3′ |
| | Reverse |
5′-TCCACCACCCTGTTGCTGTA-3′ |
Western blot analysis
Sirt6, Nephrin, Desmin, CD86, CD206 proteins were
extracted from the tissues or cells using RIPA lysis buffer
(Beyotime, Shanghai, China). The concentrations of proteins were
detected using the BCA protein kit (Beyotime, Shanghai, China).
Aliquot protein was separated by 12% SDS-PAGE and resolved proteins
were transferred onto polyvinylidene fluoride (PVDF) membranes
(Millipore), which were blocked in 5% skim milk PBS with 0.1%
Triton X-100 and incubated with primary antibodies as follows:
anti-Nephrin antibody (1:1,000; ab216341), anti-Desmin antibody
(1:1,000; ab15200), anti-Sirt6 antibody (1:2,000; ab191385),
anti-CD86 antibody (1:5,000; ab53004 and ab112490), anti-CD206
antibody (1:1,000; ab64693), anti-Bcl-2 antibody (1:2,000;
ab182858) and anti-Bax antibody (1:1,000; ab32503) (all from Abcam)
overnight at 4°C. The membranes were then incubated with the
appropriate HRP-conjugated secondary antibody (Proteintech Group,
Inc./Thermo Fisher Scientific, SA00001-2, 1:5,000). Protein bands
were detected with ECL (Thermo Fisher Scientific) and visualized
using Quantity One software (Bio-Rad, Hercules, CA, USA).
Statistical analysis
Statistical analysis was carried out Prism GraphPad
version 6.0 software. All data were presented as the means ±
standard deviation (SD). Differences were analyzed using one-way
analysis of variance (ANOVA) followed by Tukey's multiple
comparisons test. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
Podocyte injury and low expression of
Sirt6 in DN rat model
The fasting blood glucose levels of all the rats
administered streptozotocin were >250 mg/dl, indicating that the
rat model of diabetes was successfully established. Moreover, the
glucose levels of the rats administered streptozotocin were
noticeably higher than those of the rats not administered
streptozotocin (P<0.01; Fig.
1A). H&E staining revealed that in the DN group, the
glomerular local basement membranes had become thickened, and the
phenomena of mesangial matrix hyperplasia and renal interstitial
fibrosis were observed as well, compared to the controls (Fig. 1B). The expression of Sirt6 in the
renal tissues was examined by IHC staining. As shown in Fig. 1C, Sirt6 protein expression was
positive in the control group; however, the expression of Sirt6 was
downregulated in the rats with DN (Fig. 1C). Western blot analysis was also
used to detect the protein expression levels of Nephrin, the
podocyte injury marker, Desmin, and Sirt6. The results revealed
that the protein levels of Desmin markedly increased, and the
protein expression levels of both Nephrin and Sirt6 significantly
decreased in the tissues of the rats with DN (P<0.01; Fig. 1D), indicating that
streptozotocin-induced DN can lead to podocyte injury and a low
expression of Sirt6.
Immune function of macrophages in rats
with streptozotocin-induced DN
Monocytes were separated from the rat kidneys.
Different types of monocytes and Sirt6 expression were then
examined by western blot analysis. Compared to the controls, the
expression of the marker of the M1 type of monocytes, CD86, was
significantly upregulated, whereas that of the marker of the M2
type of monocytes, CD206, was significantly downregulated in the
rats with DN (P<0.01; Fig. 1E).
In addition, the protein expression of Sirt6 was notably decreased
in the DN group (P<0.01; Fig.
1E). Moreover, the levels of inflammatory factors were
determined to assess the immune function of macrophages. The
results revealed that the mRNA expression levels of the
pro-inflammatory cytokines, TNF-α and iNOS, were significantly
increased in the rats with DN compared with the controls
(P<0.01; Fig. 1F). However, the
mRNA expression levels of IL-4 and IL-10 were significantly
decreased in the rats with DN in comparison with the controls
(P<0.01; Fig. 1F).
Macrophages undergo M1 transformation
under high-glucose conditions in a dose-dependent manner
In order to examine the effects of glucose treatment
on macrophage phenotypes, the typical markers of M1 and M2
macrophages, and related inflammatory factors were analyzed. The
results revealed that the macrophages exhibited increased mRNA
expression levels of CD86 (HG1, P<0.05; HG2 and HG3, P<0.01;
Fig. 2A) and iNOS (P<0.01;
Fig. 2C) as the glucose
concentrations increased. However, the mRNA level of CD206 did not
differ significantly between the controls and the 3 HG groups
(macrophages were exposed to 3 different concentrations of glucose)
(P>0.05; Fig. 2B). The mRNA
levels of IL-10 (HG1, P<0.05; HG2 and HG3, P<0.01; Fig. 2D) and Sirt6 (HG2 and HG3,
P<0.01; Fig. 2E) significantly
decreased by exposing the cells to glucose in a dose-dependent
manner. These results illustrate that high glucose induces the
transformation of macrophages into the M1 phenotype and decreases
the expression of Sirt6 in macrophages.
High glucose promotes podocyte apoptosis
in a dose-dependent manner
As shown in Fig.
3A, the undifferentiated podocytes exhibited a fusiform or
triangular shape. Following subculture, the cells ceased to
proliferate, grew larger in size, and gradually differentiated and
matured (Fig. 3A). Flow cytometry
was used to examine the effects of various concentrations of
glucose on the apoptosis of differentiated podocytes (Fig. 3B and C). As the glucose
concentrations increased, podocyte apoptosis gradually increased in
a dose-dependent manner (HG2 and HG3, P<0.01; Fig. 3C). We also examined the expression
of apoptosis-related genes and Sirt6, and the results revealed that
the mRNA expression of pro-apoptotic Bax (P<0.01; Fig. 3E) significantly increased, whereas
that of Bcl-2 (HG2 and HG3, P<0.01; Fig. 3D) and Sirt6 (P<0.01, Fig. 3F) markedly decreased by exposure to
high-glucose concentrations.
Overexpression of Sirt6 induces
macrophage M2 transformation
The overexpression of Sirt6 was successfully induced
in the macrophages and podocytes by transfection with a Sirt6
overexpression plasmid. In the macrophages, both the mRNA (Fig. 4A) and protein (Fig. 4B and C) levels of Sirt6 exhibited a
noticeable upregulation in comparison with the controls or the NC
group (P<0.01). In addition, in the podocytes, the expression of
Sirt6 was consistent with that observed in the macrophages
(P<0.01; Fig. 4D-F).
Considering the effects of the overexpression of Sirt6 on
macrophage phenotypes, RT-qPCR was performed to detect the mRNA
levels of markers in macrophages. The results revealed that the
overexpression of Sirt6 significantly decreased the mRNA level of
CD86 (P<0.01; Fig. 4G) and
increased that of CD206 (P<0.01; Fig. 4H) compared to the controls,
indicating that the overexpression of Sirt6 induced macrophage M2
transformation.
Overexpression of Sirt6 in macrophages
inhibits podocyte apoptosis under high-glucose conditions
In order to examine the effects of the
overexpression of Sirt6 on podocyte apoptosis, we mimicked a
diabetic internal environment using a Transwell co-culture system
under high-glucose conditions to co-culture the macrophages and
podocytes. Moreover, flow cytometry was performed to detect the
apoptotic level (Fig. 5A).
Compared to the NM + HG + Con group [high-glucose medium (50
mmol/l) in the upper chamber and podocytes with high-glucose medium
in lower chamber], apoptosis was significantly increased in the HG
+ Con group (macrophages with high-glucose medium in upper chamber,
podocytes with high-glucose medium in lower chamber), indicating
that the macrophages significantly enhanced podocyte apoptosis in a
high-glucose environment (P<0.01; Fig. 5B). When the overexpression of Sirt6
was induced in the macrophages by transfection with a Sirt6
overexpression plasmid, podocyte apoptosis markedly decreased,
compared to that observed in the group with macrophages transfected
with the NC plasmid (P<0.01). However, once Sirt6 was
overexpressed in the podocytes, no significant differences were
observed in podocyte apoptosis between the M-Sirt6 group and the NM
+ HG + Con group (P>0.05). Of note, the apoptosis of the
podocytes overexpressing Sirt6 (induced by transfection with a
Sirt6 overexpression plasmid) still increased when these podocytes
were co-cultured with macrophages in high-glucose medium (HG +
M-Sirt6 group) (P<0.01). However, cell apoptosis was
significantly decreased when the podocytes overexpressing Sirt6
were co-cultured with macrophages also overexpressing Sirt6 by
transfection with a Sirt6 overexpression plasmid (HG+Sirt6+M-Sirt6
group) (P<0.01). The above-mentioned results suggested that the
overexpression of Sirt6 indirectly affected podocyte apoptosis.
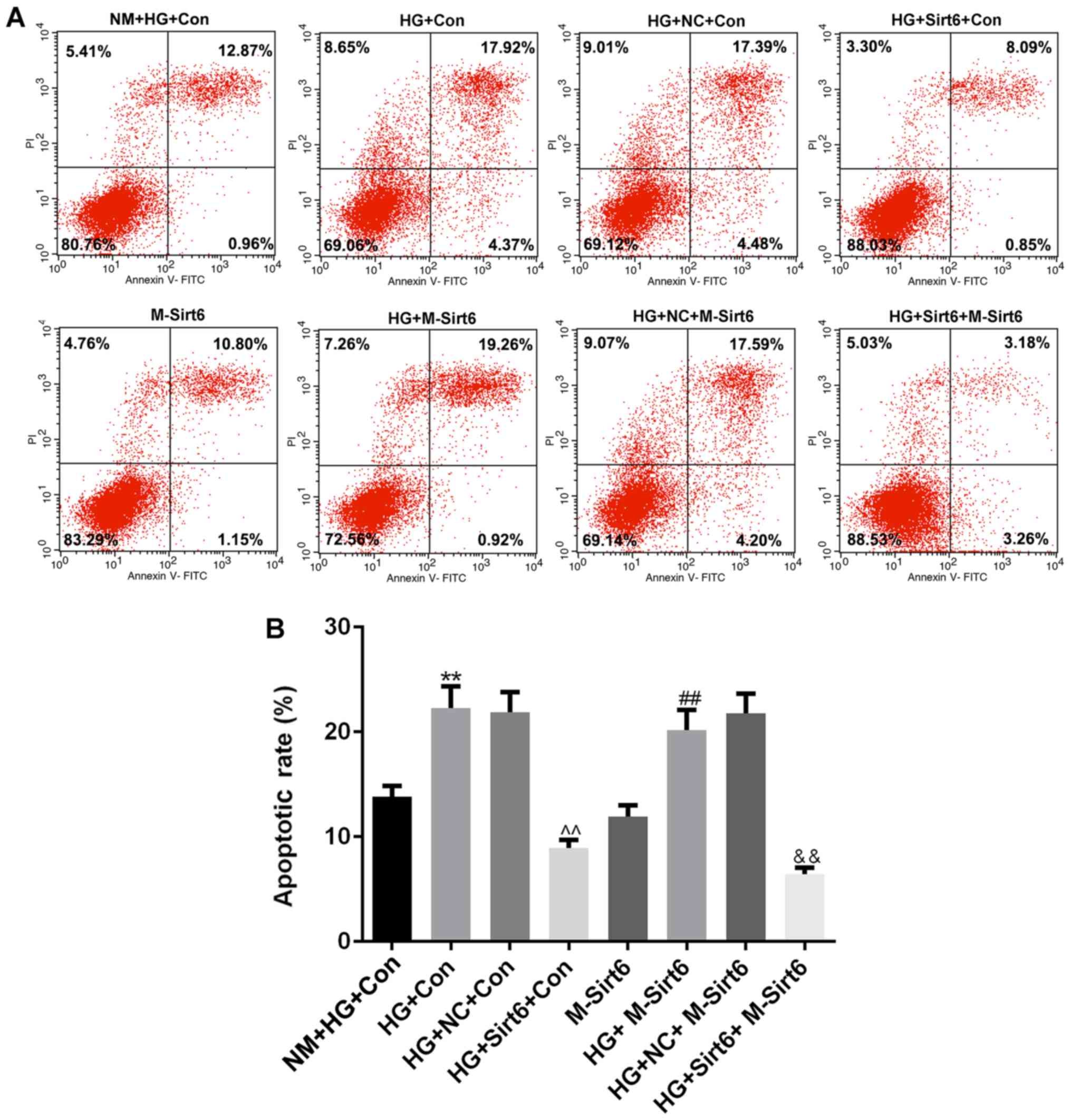 | Figure 5Overexpression of Sirt6 in
macrophages inhibits podocyte apoptosis under high-glucose
conditions. The diabetic internal environment was mimicked by a
Transwell co-culture system under high-glucose conditions to
co-culture the macrophages and podocytes. The cells in this
experiment were divided into 8 groups as follows: i) The NM + HG +
Con group (control; 50 mmol/l of high-glucose medium in the upper
chamber, and podocytes with high-glucose medium in the lower
chamber); ii) the HG + Con group (macrophages with high-glucose
medium in the upper chamber, and podocyte with high-glucose medium
in the lower chamber); iii) the HG + NC + Con group (macrophages
transfected with NC plasmid with high-glucose medium in the upper
chamber, and podocytes with high-glucose medium in the lower
chamber); iv) the HG + Sirt6 + Con group (macrophages transfected
with Sirt6 overexpression plasmid with high-glucose medium in the
upper chamber, and podocytes with high-glucose medium in the lower
chamber); v) the M-Sirt6 group (high-glucose medium in the upper
chamber, and podocytes transfected with Sirt6 overexpression
plasmid with high-glucose medium in the lower chamber); vi) the HG
+ M-Sirt6 group (macrophages with high-glucose medium in the upper
chamber, and podocytes transfected with Sirt6 overexpression
plasmid with high-glucose medium in the lower chamber); vii) the HG
+ NC + M-Sirt6 group (macrophages transfected with NC plasmid with
high-glucose medium in the upper chamber, and podocytes transfected
with Sirt6 overexpression plasmid with high-glucose medium in the
lower chamber); and viii) the HG + Sirt6 + M-Sirt6 (macrophages
transfected with Sirt6 overexpression plasmid with high-glucose
medium in the upper chamber, and podocytes transfected with Sirt6
overexpression plasmid with high-glucose medium in the lower
chamber). (A) Flow cytometry was performed to examine the effect of
the overexpression of Sirt6 on podocyte apoptosis in the Transwell
co-culture system. (B) Apoptotic rates are presented as bar
diagrams. Data are presented as the means ± SD from 3 independent
experiments (**P<0.01, compared with the NM + HG +
Con group; ^^P<0.01, compared with the HG + NC + Con
group; ##P<0.01, compared with the M-Sirt6 group;
&&P<0.01, compared with the HG + M-Sirt6
group). |
Furthermore, we detected the expression levels of
apoptosis-related genes and Sirt6 in podocytes. Under high-glucose
conditions, once the macrophages were transfected with a Sirt6
overexpression plasmid, the mRNA expression of Bcl-2 increased
(P<0.01; Fig. 6A) and that of
Bax expression decreased (P<0.01; Fig. 6B). As regards the mRNA expression
of Sirt6, the results revealed that the Sirt6 mRNA level was
upregulated when Sirt6 was overexpressed in the podocytes (Fig. 6C). The protein levels of Bcl-2, Bax
and Sirt6 were similar to the mRNA levels (Fig. 6D and E).
 | Figure 6Overexpression of Sirt6 in
macrophages regulates the expression of apoptosis-related genes and
Sirt6 in podocytes under high-glucose conditions. The diabetic
internal environment was mimicked by a Transwell co-culture system
under high-glucose conditions to co-culture the macrophages and
podocytes. The cells in this experiment were divided into 8 groups
as follows: i) The NM + HG + Con group (control; 50 mmol/l of
high-glucose medium in the upper chamber, and podocytes with
high-glucose medium in the lower chamber); ii) the HG + Con group
(macrophages with high-glucose medium in the upper chamber, and
podocyte with high-glucose medium in the lower chamber); iii) the
HG + NC + Con group (macrophages transfected with NC plasmid with
high-glucose medium in the upper chamber, and podocytes with
high-glucose medium in the lower chamber); iv) the HG + Sirt6 + Con
group (macrophages transfected with Sirt6 overexpression plasmid
with high-glucose medium in the upper chamber, and podocytes with
high-glucose medium in the lower chamber); v) the M-Sirt6 group
(high-glucose medium in the upper chamber, and podocytes
transfected with Sirt6 overexpression plasmid with high-glucose
medium in the lower chamber); vi) the HG + M-Sirt6 group
(macrophages with high-glucose medium in the upper chamber, and
podocytes transfected with Sirt6 overexpression plasmid with
high-glucose medium in the lower chamber); vii) the HG + NC +
M-Sirt6 group (macrophages transfected with NC plasmid with
high-glucose medium in the upper chamber, and podocytes transfected
with Sirt6 overexpression plasmid with high-glucose medium in the
lower chamber); and viii) the HG + Sirt6 + M-Sirt6 (macrophages
transfected with Sirt6 overexpression plasmid with high-glucose
medium in the upper chamber, and podocytes transfected with Sirt6
overexpression plasmid with high-glucose medium in the lower
chamber). RT-qPCR was performed to examine the effects of the
overexpression of Sirt6 on the mRNA expression levels of (A) Bcl-2,
(B) Bax and (C) Sirt6 in the Transwell co-culture system. (D)
Western blot analysis was performed to examine the effects of the
overexpression of Sirt6 on the protein expression levels of Bcl-2,
Bax and Sirt6 in the Transwell co-culture system. (E) Relative
protein levels are presented as bar diagrams. GAPDH was used as an
internal control. Data are presented as the means ± SD from 3
independent experiments (*P<0.05 and
**P<0.01, compared with the NM + HG + Con group;
^^P<0.01, compared with the HG+NC+Con group;
##P<0.01, compared with the M-Sirt6 group;
&&P<0.01, compared with the HG + M-Sirt6
group). |
Moreover, the macrophage phenotypes and the
expression of Sirt6 in the macrophages were subsequently detected
in the mimicked diabetic internal environment in order to examine
the possible mechanisms responsible for the protective effects of
Sirt6 overexpression against podocyte injury. We found that the
overexpression of Sirt6 in macrophages inhibited the expression of
the M1 marker, CD86 (P<0.01; Fig.
7A, D and E), and promoted that of CD206 (P<0.01; Fig. 7B, D and E), which is a marker of
the M2 phenotype expression, in the high-glucose environment at
both the mRNA and protein level. When Sirt6 overexpression was
induced in macrophages by transfection with Sirt6 overexpression
plasmid, the results revealed an increased mRNA and protein
expression of Sirt6 in the mimicked diabetic internal environment
(P<0.01; Fig. 7C-E). On the
basis of the above-mentioned results, we hypothesized that the
overexpression of Sirt6 in macrophages promoted macrophage M2
transformation, thus inhibiting podocyte apoptosis under
high-glucose conditions.
 | Figure 7Overexpression of Sirt6 in
macrophages regulates the expression of markers of macrophages and
Sirt6 under high-glucose conditions. The diabetic internal
environment was mimicked by a Transwell co-culture system under
high-glucose conditions to co-culture the macrophages and
podocytes. The cells in this experiment were divided into 8 groups
as follows: i) The NM + HG + Con group (control; 50 mmol/l of
high-glucose medium in the upper chamber, and podocytes with
high-glucose medium in the lower chamber); ii) the HG + Con group
(macrophages with high-glucose medium in the upper chamber, and
podocyte with high-glucose medium in the lower chamber); iii) the
HG + NC + Con group (macrophages transfected with NC plasmid with
high-glucose medium in the upper chamber, and podocytes with
high-glucose medium in the lower chamber); iv) the HG + Sirt6 + Con
group (macrophages transfected with Sirt6 overexpression plasmid
with high-glucose medium in the upper chamber, and podocytes with
high-glucose medium in the lower chamber); v) the M-Sirt6 group
(high-glucose medium in the upper chamber, and podocytes
transfected with Sirt6 overexpression plasmid with high-glucose
medium in the lower chamber); vi) the HG + M-Sirt6 group
(macrophages with high-glucose medium in the upper chamber, and
podocytes transfected with Sirt6 overexpression plasmid with
high-glucose medium in the lower chamber); vii) the HG + NC +
M-Sirt6 group (macrophages transfected with NC plasmid with
high-glucose medium in the upper chamber, and podocytes transfected
with Sirt6 overexpression plasmid with high-glucose medium in the
lower chamber); and viii) the HG + Sirt6 + M-Sirt6 (macrophages
transfected with Sirt6 overexpression plasmid with high-glucose
medium in the upper chamber, and podocytes transfected with Sirt6
overexpression plasmid with high-glucose medium in the lower
chamber). RT-qPCR was performed to examine the effects of the
overexpression of Sirt6 on the mRNA expression levels of (A) CD86,
(B) CD206, and (C) Sirt6 in the Transwell co-culture system. (D)
Western blot analysis was performed to examine the effects of the
overexpression of Sirt6 on the protein expression levels of CD86,
CD206 and Sirt6 in the Transwell co-culture system. (E) Relative
protein levels are presented as bar diagrams. GAPDH was used as an
internal control. Data are presented as the means ± SD from 3
independent experiments (^^P<0.01, compared with the
HG + Con group; &&P<0.01, compared with the
HG + M-Sirt6 group). |
Discussion
In this study, we used streptozocin to induce
diabetes in rats as previously described (25-28).
The inflammatory process and potocyte injury in the rats with DN
were subsequently analyzed. Podocytes play an important role in
maintaining glomerular structural integrity and filtration
function. Podocytes are characterized by being able to express
specific functional protein molecules, such as Nephrin, Podocin,
Podocalyxin and Desmin (29-33).
Podocyte injury is considered to be the leading cause of
proteinuria and glomerular disease (34,35).
In this study, in the tissues of rats with DN, we found that
Nephrin protein expression decreased and Desmin protein expression
increased. These are typical podocyte injury markers that allow
Nephrin gene mutation to lead to proteinuria production and
congenital nephrotic syndrome. In addition, a high expression of
Desmin and its phenotypic transformation causes the re-arrangement
of the podocyte cytoskeleton (36,37).
Previously, Bohle et al reported that a large number of
inflammatory cells, which either were macrophages, or were
lymphocytes, were found in renal tissues of 488 patients with DN by
immunostaining; it was thus hypothesized that the pathogenesis of
DN was related to tubulointerstitial immune inflammation (38). An increasing number of studies have
confirmed that macrophage aggregation is a predictor of renal
dysfunction in patients with DN (9,23,39-41).
In this study, we detected the degree of macrophage infiltration
and associated inflammatory factor expression in rats with DN. Some
studies have found that symbolic molecules of M1 macrophages
include TNF-α, IL-6, monocyte chemoattractant protein-1 (MCP-1) and
iNOS, while representative M2 macrophages molecules include IL-10,
CD206, CD163 and chemokine (C-C motif) ligand 18 (CCL18; also known
as AMAC1) (42, 43). The findings of this study
demonstrated that CD86 expression was significantly upregulated
under high-glucose conditions and that of CD206 exhibited an
opposite trend. Additionally, TNF-α and iNOS mRNA expression levels
increased and the mRNA expression levels of IL-4 and IL-10
decreased under high-glucose conditions. Recently, Assadi-Porter
et al reported the effects of 3-iodothyronamine (T1AM) on
obesity and their results also demonstrated that these effects were
co-regulated by Sirt6 and Sirt4; in detail, Sirt6 regulated
glycolysis, triglyceride synthesis and fat metabolism, while Sirt4
had an opposite effect, negatively regulating fatty acid oxidation
(44). The results of their study
were similar to those of this study in that the expression of Sirt6
was significantly decreased in the tissues of rats with DN.
Furthermore, in future studies, we aim to examine the co-regulation
of Sirt 6 and Sirt4 in diabetes-induced renal injury.
In this study, macrophages and podocytes were
respectively used to conduct in vitro experiments with a
series of detections. The results revealed that glucose promoted
macrophage M1 transformation and podocyte apoptosis in a
dose-dependent manner, and high glucose decreased Sirt6 expression
in rats with DN.
Increasing evidence indicates that Sirtuins,
particularly Sirt1, Sirt3 and Sirt6, are related to kidney disease
(45-47). Several studies have demonstrated
that the expression of Sirt1 in DN decreases, and that the
increased Sirt1 expression can attenuate renal injury in DN
(48-51). Caton et al reported that the
decreased expression of Sirt3 resulted in an impaired function of
β-cells in type 2 diabetes mellitus (52). Sirt3, as a protective agent of
acute tissue injury, has also been shown to play an important role
in acute renal injury model (53).
Sirt6 is considered to be related to the occurrence and development
of chronic diseases, including diabetes, and such an effect is
realized by its role in DNA damage (54-56).
Therefore, in this study, the overexpression of Sirt6 was
successfully indcued in macrophages and podocytes, respectively,
and we found that the overexpression of Sirt6 promoted macrophage
M2 transformation. It is unusual that a lack of Sirt6 would
increase the inflammatory response in tissues. For example, a lack
of Sirt6 in adipose tissue has been shown to increased macrophage
infiltration and adipose tissue inflammation and to promote
HFD-induced insulin resistance (57,58).
In addition, its deficiency causes liver inflammation and fibrosis
in mouse immune cells (59).
However, the effects of Sirt4 on related inflammation, insulin
resistance and oxidative metabolism are not consistent with those
of Sirt6. For example, Sirt4 modulates oxidative metabolism and
directly leads to endothelial dysfunction, and is related to
coronary artery disease (60).
Huynh et al reported that the loss of Sirt4 elevated
glucose- and leucine-stimulated insulin levels in vivo,
which were dependent upon a C57BL/6NJ genetic background, and
enhanced age-induced insulin resistance (61). This is a very interesting research
prospect, which is worthy of further study.
Furthermore, in this study, we mimicked the diabetic
internal environment by a Transwell co-culture system under
high-glucose conditions to co-culture macrophages and podocytes. We
found that the overexpression of Sirt6 in macrophages protected
podocytes from high-glucose-induced injury, whereas the
overexpression of Sirt6 in podocytes exerted no protective effect
itself against injury induced by high glucose. In addition, we
found that the possible mechanism responsible for the protective
effects of Sirt6 overexpression in macrophages against podocyte
injury involved the promotion of the macrophage M2 transformation.
Such a result is in line with the findings from the study by Sun
et al, who reported that by inducing M2 macrophage
differentiation, pentraxin-3 alleviated kidney damage in DN
(23). Zhang et al also
suggested that the inhibition of M1 macrophage activation and the
promotion of M2 macrophage transformation prevented podocyte injury
(62).
Taken together, the findings of this study suggested
that Sirt6 protected against podocytes from injury in a mimicked
diabetic kidney microenvironment by activating M2 macrophages. Our
findings also indicated that Sirt6 functioned as an immune response
regulatory factor in renal inflammatory injury induced by
streptozotocin administration in rats. It would be more convincing
to provide more kidney function indicators [blood urea nitrogen
(BUN) and serum creatinine (SCr)] and a more general method for
modeling DN in animals. These are the limitations of our study and
we aim to perform a more detailed and in-depth analysis in the
future.
Funding
This study was supported by the Hangzhou Science and
Technology Commission 'the Warehousing project of Hangzhou
Agricultural and Social Development Research Project Library'
(164783); the Zhejiang Provincial Chinese Medicine Administration
'Zhejiang Traditional Chinese Medicine Research Fund Project'
(2016ZA155); and the Zhejiang Provincial Health and Family Planning
Commission 'Zhejiang Medical and Health Research Fund Project'
(2017KY129).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
LJ made substantial contributions to the conception
and design of the study. HW and WZ were involved in data
acquisition, data analysis and interpretation. YC and LH were
involved in the cell experiments and the drafting of the article or
critically revising it for important intellectual content. JW and
YL were involved in the animal experiments. All authors are
accountable for data integrity, and have read and approved the
final manuscript.
Ethics approval and consent to
participate
The animal experiments were performed according to
the Guide for the Care and Use of Laboratory Animals and approved
by the First People's Hospital of Yuhang District, Hangzhou Animal
Ethics Committee (Hangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Gallagher H and Suckling RJ: Diabetic
nephropathy: Where are we on the journey from pathophysiology to
treatment? Diabetes Obes Metab. 18:641–647. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dronavalli S, Duka I and Bakris GL: The
pathogenesis of diabetic nephropathy Nat Clin Pract Endocrinol
Metab. 4:444–452. 2008. View Article : Google Scholar
|
|
3
|
Ma R, Liu L, Jiang W, Yu Y and Song H:
FK506 ameliorates podocyte injury in type 2 diabetic nephropathy by
down-regulating TRPC6 and NFAT expression. Int J Clin Exp Pathol.
8:14063–14074. 2015.
|
|
4
|
Eremina V, Baelde HJ and Quaggin SE: Role
of the VEGF–a signaling pathway in the glomerulus: Evidence for
crosstalk between components of the glomerular filtration barrier.
Nephron, Physiol. 106:32–37. 2007. View Article : Google Scholar
|
|
5
|
Mundel P and Kriz W: Structure and
function of podocytes: An update. Anat Embryol (Berl). 192:385–397.
1995. View Article : Google Scholar
|
|
6
|
Mathieson PW: What has the immune system
got against the glomerular podocyte? Clin Exp Immunol. 134:1–5.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gordon S and Taylor PR: Monocyte and
macrophage heterogeneity Nat Rev Immunol. 5:953–964. 2005.
|
|
8
|
Chow F, Ozols E, Nikolic-Paterson DJ,
Atkins RC and Tesch GH: Macrophages in mouse type 2 diabetic
nephropathy: Correlation with diabetic state and progressive renal
injury. Kidney Int. 65:116–128. 2004. View Article : Google Scholar
|
|
9
|
Nguyen D, Ping F, Mu W, Hill P, Atkins RC
and Chadban SJ: Macrophage accumulation in human progressive
diabetic nephropathy. Nephrology (Carlton). 11:226–231. 2006.
View Article : Google Scholar
|
|
10
|
Tesch GH: Macrophages and diabetic
nephropathy. Semin Nephrol. 30:290–301. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Minton K: Macrophages: A transcription
factor to call their own. Nat Rev Immunol. 11:742011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hofkens W, Storm G, Berg WVD and Lent PV:
Inhibition of M1 macrophage activation in favour of M2
differentiation by liposomal targeting of glucocorticoids to the
synovial lining during experimental arthritis. Ann Rheum Dis.
70(Suppl 2): pp. 702011, View Article : Google Scholar
|
|
13
|
Gordon S: Alternative activation of
macrophages. Nat Rev Immunol. 3:23–35. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gertler AA and Cohen HY: SIRT6, a protein
with many faces. Biogerontology. 14:629–639. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tennen RI, Berber E and Chua KF:
Functional dissection of SIRT6: Identification of domains that
regulate histone deacetylase activity and chromatin localization.
Mech Ageing Dev. 131:185–192. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tennen RI, Bua DJ, Wright WE and Chua KF:
SIRT6 is required for maintenance of telomere position effect in
human cells. Nat Commun. 2:4332011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Beauharnois JM, Bolívar BE and Welch JT:
Sirtuin 6: A review of biological effects and potential therapeutic
properties. Mol Biosyst. 9:1789–1806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Michishita E, McCord RA, Berber E, Kioi M,
Padilla-Nash H, Damian M, Cheung P, Kusumoto R, Kawahara TL,
Barrett JC, et al: SIRT6 is a histone H3 lysine 9 deacetylase that
modulates telomeric chromatin. Nature. 452:492–496. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yuan J, Pu M, Zhang Z and Lou Z: Histone
H3-K56 acetylation is important for genomic stability in mammals.
Cell Cycle. 8:1747–1753. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mostoslavsky R, Chua KF, Lombard DB, Pang
WW, Fischer MR, Gellon L, Liu P, Mostoslavsky G, Franco S, Murphy
MM, et al: Genomic instability and aging-like phenotype in the
absence of mammalian SIRT6. Cell. 124:315–329. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhong L, D'Urso A, Toiber D, Sebastian C,
Henry RE, Vadysirisack DD, Guimaraes A, Marinelli B, Wikstrom JD,
Nir T, et al: The histone deacetylase Sirt6 regulates glucose
homeostasis via Hif1alpha. Cell. 140:280–293. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang X, Liu J, Zhen J, Zhang C, Wan Q, Liu
G, Wei X, Zhang Y, Wang Z, Han H, et al: Histone deacetylase 4
selectively contributes to podocyte injury in diabetic nephropathy.
Kidney Int. 86:712–725. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sun H, Tian J, Xian W, Xie T and Yang X:
Pentraxin-3 attenuates renal damage in diabetic nephropathy by
promoting M2 macrophage differentiation. Inflammation.
38:1739–1747. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
25
|
Zanotto C, Hansen F, Galland F, Batassini
C, Federhen BC, da Silva VF, Leite MC, Nardin P and Gonçalves CA:
Glutamatergic alterations in STZ-induced diabetic rats are reversed
by Exendin-4. Mol Neurobiol. Aug 25–2018.Epub ahead of print.
View Article : Google Scholar : 2018.PubMed/NCBI
|
|
26
|
Nunes KP, de Oliveira AA, Szasz T,
Biancardi VC and Webb RC: Blockade of toll-like receptor 4
attenuates erectile dysfunction in diabetic rats. J Sex Med.
15:1235–1245. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang YC, Wu BH, Chu YL, Chang WC and Wu
MC: Effects of tempeh fermentation with Lactobacillus plantarum and
Rhizopus oligosporus on streptozotocin-induced type II diabetes
mellitus in rats. Nutrients. 10:102018. View Article : Google Scholar
|
|
28
|
Wu M, Yang Y, Wang M, Zeng F, Li Q, Liu W,
Guo S, He M, Wang Y, Huang J, et al: Exogenous pancreatic
kallikrein improves diabetic cardiomyopathy in
streptozotocin-induced diabetes. Front Pharmacol. 9:8552018.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Inoki K, Mori H, Wang J, Suzuki T, Hong S,
Yoshida S, Blattner SM, Ikenoue T, Rüegg MA, Hall MN, et al: mTORC1
activation in podocytes is a critical step in the development of
diabetic nephropathy in mice. J Clin Invest. 121:2181–2196. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang W, Wang Y, Long J, Wang J, Haudek SB,
Overbeek P, Chang BH, Schumacker PT and Danesh FR: Mitochondrial
fission triggered by hyperglycemia is mediated by ROCK1 activation
in podocytes and endothelial cells. Cell Metab. 15:186–200. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Casalena G, Krick S, Daehn I, Yu L, Ju W,
Shi S, Tsai SY, D'Agati V, Lindenmeyer M, Cohen CD, et al: Mpv17 in
mitochondria protects podocytes against mitochondrial dysfunction
and apoptosis in vivo and in vitro. Am J Physiol Renal Physiol.
306:F1372–F1380. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bechtel W, Helmstädter M, Balica J,
Hartleben B, Kiefer B, Hrnjic F, Schell C, Kretz O, Liu S, Geist F,
et al: Vps34 deficiency reveals the importance of endocytosis for
podocyte homeostasis. J Am Soc Nephrol. 24:727–743. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu N, Xu L, Shi Y and Zhuang S: Podocyte
autophagy: A potential therapeutic target to prevent the
progression of diabetic nephropathy. J Diabetes Res.
2017:35602382017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Inoue R, Asanuma K, Seki T, Nagase M and
Osafune K: New therapeutic insights for chronic kidney disease
provided by podocytology. Nihon Yakurigaku Zasshi. 143:27–33.
2014.In Japanese. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lennon R, Randles MJ and Humphries MJ: The
importance of podocyte adhesion for a healthy glomerulus. Front
Endocrinol (Lausanne). 5:1602014. View Article : Google Scholar
|
|
36
|
Kestilä M, Lenkkeri U, Männikkö M,
Lamerdin J, McCready 6P, Putaala H, Ruotsalainen V, Morita T,
Nissinen M, Herva R, et al: Positionally cloned gene for a novel
glomerular protein - nephrin - is mutated in congenital nephrotic
syndrome. Mol Cell. 1:575–582. 1998. View Article : Google Scholar
|
|
37
|
Wang Z, Liu J and Sun W: Effects of
asiaticoside on levels of podocyte cytoskeletal proteins and renal
slit diaphragm proteins in adriamycin-induced rat nephropathy. Life
Sci. 93:352–358. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bohle A, Wehrmann M, Bogenschütz O, Batz
C, Müller CA and Müller GA: The pathogenesis of chronic renal
failure in diabetic nephropathy. Investigation of 488 cases of
diabetic glomerulo-sclerosis. Pathol Res Pract. 187:251–259. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Furuta T, Saito T, Ootaka T, Soma J, Obara
K, Abe K and Yoshinaga K: The role of macrophages in diabetic
glomerulo-sclerosis. Am J Kidney Dis. 21:480–485. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yonemoto S, Machiguchi T, Nomura K,
Minakata T, Nanno M and Yoshida H: Correlations of tissue
macrophages and cytoskeletal protein expression with renal fibrosis
in patients with diabetes mellitus. Clin Exp Nephrol. 10:186–192.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Guo Y, Song Z, Zhou M, Yang Y, Zhao Y, Liu
B and Zhang X: Infiltrating macrophages in diabetic nephropathy
promote podocytes apoptosis via TNF-α-ROS-p38MAPK pathway.
Oncotarget. 8:53276–53287. 2017.PubMed/NCBI
|
|
42
|
Liu H, Dong H, Jiang L, Li Z and Ma X:
Bleomycin inhibits proliferation and induces apoptosis in TPC-1
cells through reversing M2-macrophages polarization. Oncol Lett.
16:3858–3866. 2018.PubMed/NCBI
|
|
43
|
Heusinkveld M, de Vos van Steenwijk PJ,
Goedemans R, Ramwadhdoebe TH, Gorter A, Welters MJ, van Hall T and
van der Burg SH: M2 macrophages induced by prostaglandin E2 and
IL-6 from cervical carcinoma are switched to activated M1
macrophages by CD4+ Th1 cells. J Immunol. 187:1157–1165.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Assadi-Porter FM, Reiland H, Sabatini M,
Lorenzini L, Carnicelli V, Rogowski M, Selen Alpergin ES, Tonelli
M, Ghelardoni S, Saba A, et al: Metabolic reprogramming by
3-iodothyronamine (T1AM): A new perspective to reverse obesity
through co-regulation of sirtuin 4 and 6 expression. Int J Mol Sci.
19:192018. View Article : Google Scholar
|
|
45
|
Aditya R, Kiran AR, Varma DS, Vemuri R and
Gundamaraju R: A review on SIRtuins in diabetes. Curr Pharm Des.
23:2299–2307. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Feng J, Lu C, Dai Q, Sheng J and Xu M:
SIRT3 facilitates amniotic fluid stem cells to repair diabetic
nephropathy through protecting mitochondrial homeostasis by
modulation of mitophagy. Cell Physiol Biochem. 46:1508–1524. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Huynh FK, Hershberger KA and Hirschey MD:
Targeting sirtuins for the treatment of diabetes. Diabetes Manag
(Lond). 3:245–257. 2013. View Article : Google Scholar
|
|
48
|
Kitada M, Takeda A, Nagai T, Ito H,
Kanasaki K and Koya D: Dietary restriction ameliorates diabetic
nephropathy through anti-inflammatory effects and regulation of the
autophagy via restoration of Sirt1 in diabetic Wistar fatty (fa/fa)
rats: A model of type 2 diabetes. Exp Diabetes Res.
2011:9081852011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li C, Cai F, Yang Y, Zhao X, Wang C, Li J,
Jia Y, Tang J and Liu Q: Tetrahydroxystilbene glucoside ameliorates
diabetic nephropathy in rats: Involvement of SIRT1 and TGF-β1
pathway. Eur J Pharmacol. 649:382–389. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Shang G, Gao P, Zhao Z, Chen Q, Jiang T,
Zhang N and Li H: 3,5-Diiodo-l-thyronine ameliorates diabetic
nephropathy in streptozotocin-induced diabetic rats. Biochim
Biophys Acta. 1832:674–684. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang X, Meng L, Zhao L, Wang Z, Liu H, Liu
G and Guan G: Resveratrol ameliorates hyperglycemia-induced renal
tubular oxidative stress damage via modulating the SIRT1/FOXO3a
pathway. Diabetes Res Clin Pract. 126:172–181. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Caton PW, Richardson SJ, Kieswich J,
Bugliani M, Holland ML, Marchetti P, Morgan NG, Yaqoob MM, Holness
MJ and Sugden MC: Sirtuin 3 regulates mouse pancreatic beta cell
function and is suppressed in pancreatic islets isolated from human
type 2 diabetic patients. Diabetologia. 56:1068–1077. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Morigi M, Perico L, Rota C, Longaretti L,
Conti S, Rottoli D, Novelli R, Remuzzi G and Benigni A: Sirtuin
3-dependent mitochondrial dynamic improvements protect against
acute kidney injury. J Clin Invest. 125:715–726. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Blasiak J, Arabski M, Krupa R, Wozniak K,
Zadrozny M, Kasznicki J, Zurawska M and Drzewoski J: DNA damage and
repair in type 2 diabetes mellitus. Mutat Res. 554:297–304. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Grindel A, Guggenberger B, Eichberger L,
Pöppelmeyer C, Gschaider M, Tosevska A, Mare G, Briskey D, Brath H
and Wagner KH: Oxidative stress, DNA damage and DNA repair in
female patients with diabetes mellitus type 2. PLoS One.
11:e01620822016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kuang J, Chen L, Tang Q, Zhang J, Li Y and
He J: The role of Sirt6 in obesity and diabetes. Front Physiol.
9:1352018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kuang J, Zhang Y, Liu Q, Shen J, Pu S,
Cheng S, Chen L, Li H, Wu T, Li R, et al: Fat-specific Sirt6
ablation sensitizes mice to high-fat diet-induced obesity and
insulin resistance by inhibiting lipolysis. Diabetes. 66:1159–1171.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Xiong X, Zhang C, Zhang Y, Fan R, Qian X
and Dong XC: Fabp4-Cre-mediated Sirt6 deletion impairs adipose
tissue function and metabolic homeostasis in mice. J Endocrinol.
233:307–314. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Xiao C, Wang RH, Lahusen TJ, Park O,
Bertola A, Maruyama T, Reynolds D, Chen Q, Xu X, Young HA, et al:
Progression of chronic liver inflammation and fibrosis driven by
activation of c-JUN signaling in Sirt6 mutant mice. J Biol Chem.
287:41903–41913. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Tarantino G, Finelli C, Scopacasa F,
Pasanisi F, Contaldo F, Capone D and Savastano S: Circulating
levels of sirtuin 4, a potential marker of oxidative metabolism,
related to coronary artery disease in obese patients suffering from
NAFLD, with normal or slightly increased liver enzymes. Oxid Med
Cell Longev. 2014:9206762014. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Huynh FK, Hu X, Lin Z, Johnson JD and
Hirschey MD: Loss of sirtuin 4 leads to elevated glucose- and
leucine-stimulated insulin levels and accelerated age-induced
insulin resistance in multiple murine genetic backgrounds. J
Inherit Metab Dis. 41:59–72. 2018. View Article : Google Scholar :
|
|
62
|
Zhang XL, Guo YF, Song ZX and Zhou M:
Vitamin D prevents podocyte injury via regulation of macrophage
M1/M2 phenotype in diabetic nephropathy rats. Endocrinology.
155:4939–4950. 2014. View Article : Google Scholar : PubMed/NCBI
|


















