Introduction
Acute pancreatitis (AP) is a common acute abdominal
disease, and ~10–20% of AP patients develop severe acute
pancreatitis (SAP)(1). SAP is
associated with poor prognosis and mortality rates as high as
15–20% (2). Current conservative
therapies including inhibition of pancreatic juice synthesis and
secretion, prophylactic use of antibiotics and nutritional support,
are most frequently utilized in the treatment of SAP in clinical
practice, however these strategies are of poor efficacy.
Mesenchymal stem cells (MSCs) are a type of adult
stem cell in humans, characterized by their low immunogenicity and
multiple differentiation potentials. Under certain conditions, MSCs
are able to differentiate into various tissue cell types, including
myocardial cells, endothelial cells and insulin-secreting cells
(3–5). Several studies have demonstrated that
MSCs are involved in the pathogenesis of AP, revealing in their
investigations that transplanted MSCs have the capacity to relieve
the severity and improve the prognosis of SAP (6–8).
While it is understood that MSCs have the ability to migrate into
the damaged organ, the mechanism of this effect is largely
unknown.
Stromal cell-derived factor-1 (SDF-1) is a member of
the CXC chemokine family, and binds to its highly specific
corresponding receptor, CXC-chemokine receptor 4 (CXCR4), forming a
complex from which it regulates numerous physiological functions.
Previous studies have reported that the SDF-1/CXCR4 axis not only
is involved in the invasion and metastasis of malignant tumors
(9,10), however is also critical in the
migration of stem cells. Du et al (11) identified that SDF-1 is able to
significantly enhance the migration and differentiation of
periodontal ligament stem cells to repair damaged periodontal
tissues. Similarly, Theiss et al (12) demonstrated that the SDF-1/CXCR4
axis, which promoted stem cell homing, may be important in the
treatment of myocardial infarction. Furthermore, SDF-1 expression
in tissues appears to be significantly increased under hypoxic
conditions, including during myocardial ischemia (13), cerebral ischemia (14) and renal failure (15). Thus, we hypothesize that
SDF-1/CXCR4 axis may also promote the migration of MSCs towards the
injured pancreas in AP.
In this study, SDF-1 expression in the pancreas of
AP rats, and the in vitro effect of SDF-1 on the migration
of bone marrow mesenchymal stem cells (BMSCs), was investigated. In
addition, the BMSCs were transplanted into an in vivo rat
model of AP via the tail vein to examine the role of SDF-1/CXCR4
axis in the migration of BMSCs in the injured pancreas.
Materials and methods
Induction of AP
All animal procedures were conducted according to
the Shanghai Laboratory Animal Ordinance and approved by the Ethics
Committee of Shanghai Tenth People’s Hospital (Tongji University,
Shanghai, China). Sprague-Dawley (SD) rats were purchased from
Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China). These
animals were housed under standardized conditions in a 12 h
dark/light cycle with free access to water and food. Rats were
fasted overnight preoperatively. Following being anesthetized by
intraperitoneal injection of 3% pentobarbital, a 2 cm midline
laparotomy was performed and the first loop of the duodenum and the
pancreas were separated. A blunt fine catheter was then introduced
into the bile-pancreatic duct and 3% sodium taurocholate (1 ml/kg;
Sigma-Aldrich, St. Louis, MO, USA) was retrogradely injected into
the common bile duct within 60 sec. Leakage of sodium taurocholate
was prevented by temporary ligation of the distal bile duct with a
9-0 prolene suture, while the proximal bile duct was temporarily
occluded with a microvessel clip. Following infusion, the blunt
fine catheter, the microvessel clip and the suture were removed to
allow the physiological flow of bile. The puncture site on the
duodenum was closed with a 6-0 prolene suture and the wound with a
4-0 prolene suture.
Isolation, culture and identification of
rat BMSCs
Four week-old SD rats were sacrificed and the
primary BMSCs were isolated from the femurs and tibias under a
sterile condition. Following flushing the bone marrow cavity with
DMEM-LG medium (Invitrogen Life Technologies, Grand Island, NY,
USA) and removing large tissues with a 200-mesh nylon filter, the
isolated bone marrow cells were cultured in DMEM-LG medium
supplemented with 10% fetal bovine serum (FBS; Invitrogen Life
Technologies), 1% penicillin and streptomycin (Invitrogen Life
Technologies) at 37°C in an environment with 5% CO2. The
medium was refreshed every 3 days and cells were harvested by
multiple digestions and passaged when cell confluence reached
>80%. Cells of the fifth passage were collected and subjected to
flow cytometry analysis. These cells were maintained in adipogenic,
osteogenic and chondrogenic differentiation medium (Cyagen
Biosciences, Sunnyvale, CA, USA) followed by Oil Red O staining,
Alizarin Red S staining and Toluidine Blue staining, respectively.
The surface markers were detected using a BD FACSC™ II flow
cytometry system (BD Biosciences, San Jose, CA, USA). The
antibodies utilized in this analysis included anti-CD44-PE,
anti-CD73-APC, anti-CD90-FITC, anti-CD105-PerCP-Cy5.5,
anti-CD11b-PE, anti-CD19-PE, anti-CD34-PE and anti-CD45-PE (BD
Biosciences). Isotype-matched antibodies were used as controls.
Cells were harvested between 3–5 passages.
Immunohistochemistry and western blot
analysis
Following AP induction, pancreatic tissues were
obtained on days 1, 3, 5, 7 and 10, and normal pancreatic tissues
were collected as controls. Immunocytochemistry and western blot
analysis were employed to detect the SDF-1 expression. Five SD rats
were used at each time point.
After preparation of the paraffin sections,
Histostain-Plus kit (DAB, Rabbit; Invitrogen Life Technologies) was
used to perform immunohistochemistry. Following conventional
dehydration, antigen retrieval and antigen blocking, sections were
incubated with rabbit anti-SDF-1 antibody (1:50; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) overnight at 4°C in a wet
box. Then, the biotinylated secondary antibody and streptavidin
conjugated horseradish peroxidase (HRP) were independently
incubated with these sections for 10 min at 37°C. Following
staining with DAB, the sections were counterstained with
hematoxylin and then dehydrated and mounted with neutral resin.
Pancreatic tissues were added to liquid nitrogen and
crushed. The total protein was extracted with the conventional
method and was followed by determination of protein concentration
with bicinchoninic acid method. Subsequently, 30 mg of proteins
were subjected to 12% polyacrylamide-SDS gel electrophoresis and
electroblotted onto nitrocellulose membranes, which were then
incubated with rabbit anti-SDF-1 antibody (dilution, 1:200; Santa
Cruz Biotechnology, Inc.) overnight at 4°C following blocking with
5% skimmed milk for 1 h. Incubation with the secondary antibody at
room temperature for 2 h was conducted and then visualization was
performed, and the protein bands were scanned (Gene Company
Limited, Hong Kong, China) and analyzed using Odyssey 3.0 analysis
software (Li-Cor Bioscience, Lincoln, NE, USA).
Immunofluorescence staining
The CXCR4 expression profile on BMSCs was assessed
by immunofluorescence staining. Cells were collected and fixed in
4% paraformaldehyde for 20 min. Following treatment with 0.5%
Triton X-100 and 1% bovine serum albumin (BSA), cells were
incubated with the rabbit anti-CXCR4 antibody (dilution, 1:50;
Santa Cruz Biotechnology, Inc.) overnight at 4°C. After washing
with PBS, cells were incubated with the fluorescein isothiocyanate
(FITC)-conjugated secondary antibody for 30 min at 37°C followed by
DAPI (Sigma-Aldrich) staining for 2 min, for nuclear
counterstaining. The sections were mounted and then observed under
a Zeiss LSM 710 confocal microscope (Carl Zeiss, Oberkochen,
Germany).
Transwell migration assay
The migration assay was designed using transwell
plates (Corning Costar, Cambridge, MA, USA) that were 6.5 mm in
diameter with 8 μm pore filters. In the chemotaxis assay, the upper
chambers were loaded with 5×104 BMSCs in 200 μl of DMEM
containing 0.1% BSA, and the lower chambers with 500 μl of DMEM
containing 10% FBS and SDF-1 at different concentrations
(Peprotech, Inc., Rocky Hill, NJ, USA). SDF-1 at 0, 10, 50, and 100
ng/ml was used according to the manufacturer’s instructions. In the
chemotaxis inhibition assay, the upper chambers were inoculated
with 5×104 BMSCs that were incubated with rabbit
anti-CXCR4 antibody (10 μg/ml; Santa Cruz Biotechnology, Inc.) or
AMD3100 (10 μg/ml; Sigma-Aldrich), an antagonist of CXCR4, for 2 h
at 37°C. The lower chambers were added with 500 μl of DMEM
containing 10% FBS and SDF-1 at optimal concentration. Following
incubation for 15 h, cells in the upper chamber were removed and
the membranes were fixed in 4% paraformaldehyde for 20 min. The
cells that migrated to the lower side of the filter were stained
with 0.1% crystal violet for 10 min and then observed under a light
microscope. Crystal violet was dissolved in 300 μl of 33% acetic
acid and the absorbance at 573 nm was measured with an
enzyme-labeling measuring instrument (Gene Company Limited).
Animal experiments
To trace BMSCs in vivo, cells were labeled
with
chloromethylbenzamido-1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine
perchlorate (CM-Dil; 1 μg/ml; Invitrogen Life Technologies) at 37°C
for 30 min. Several studies have demonstrated that labeling cells
with CM-Dil does not affect cell viability, proliferation or
differentiation. The labeling efficiency was determined by
fluorescence microscopy (16,17).
Rats were randomly divided into four groups: the
control group (n=15), the AP group (n=15), the BMSCs group (n=15)
and the anti-CXCR4 group (n=15). Rats were sacrificed at days 1, 4
and 7 (n = 5/time point). Rats in the anti-CXCR4 group received an
injection of CM-Dil-labeled BMSCs (1×107 cells/ml/kg)
and were pretreated with anti-CXCR4 antibody via the tail vein.
Rats in the BMSC group received an injection of CM-Dil-labeled
BMSCs (1×107 cells/ml/kg). In the control and AP group,
rats were treated with normal saline of equal volume. Following
sacrificing, the pancreas and blood were collected. Each pancreatic
tissue was divided into two sections: one was fixed in 4%
paraformaldehyde for immunohistochemistry and HE staining, and the
other was prepared for the frozen sections, in order to detect the
migration of BMSCs towards the pancreatic tissues by confocal
microscopy (Zeiss). The images were captured at five randomly
selected fields at a high magnification and the migrated
CM-Dil-labeled BMSCs (red cells) were calculated. Blood was
centrifuged and supernatant collected for serum amylase
analysis.
Histology and serum amylase
Paraffin sections were prepared for
histopathological examination with conventional HE staining. In
order to quantify the pancreatic injury, 20 microscopic fields were
randomly selected as previously described (18). In brief, the edema, inflammatory
cell infiltration and acinar necrosis were divided into four grades
(edema: 0 = absent, 1 = focally lobular edema, 2 = diffusely
lobular edema and 3 = acini edema and separated; inflammatory cell
infiltration: 0 = absent, 1 = infiltration around ductal margins, 2
= infiltration into <50% of lobules and 3 = infiltration of
>50% of lobules; acinar necrosis: 0 = absent, 1 = necrosis in
<5% of lobules, 2 = necrosis in 5–20% of lobules and 3 =
necrosis in 20–50% of lobules).
The amylase activity assay kit (Biovision, Mountain
View, CA, USA) was used to determinate the serum amylase activity.
The nitrophenol standard curve was delineated. Then, the optical
density (OD) immediately prior to reaction (ODT0) and at
10 min following reaction (ODT1) was measured at 405 nm.
The difference of OD was calculated as follows: ΔOD =
ODT1 − ODT0, and the concentration of
nitrophenol was obtained according to the standard curve and then
converted into the amylase activity.
Statistical analysis
Statistical analysis was performed using SPSS
version 14.0 for Windows (SPSS, Inc., Chicago, IL, USA). Data were
presented as the mean ± standard deviation (SD) and were analyzed
with a Student’s t-test. A value of P<0.05 was considered to
indicate a statistically significant result.
Results
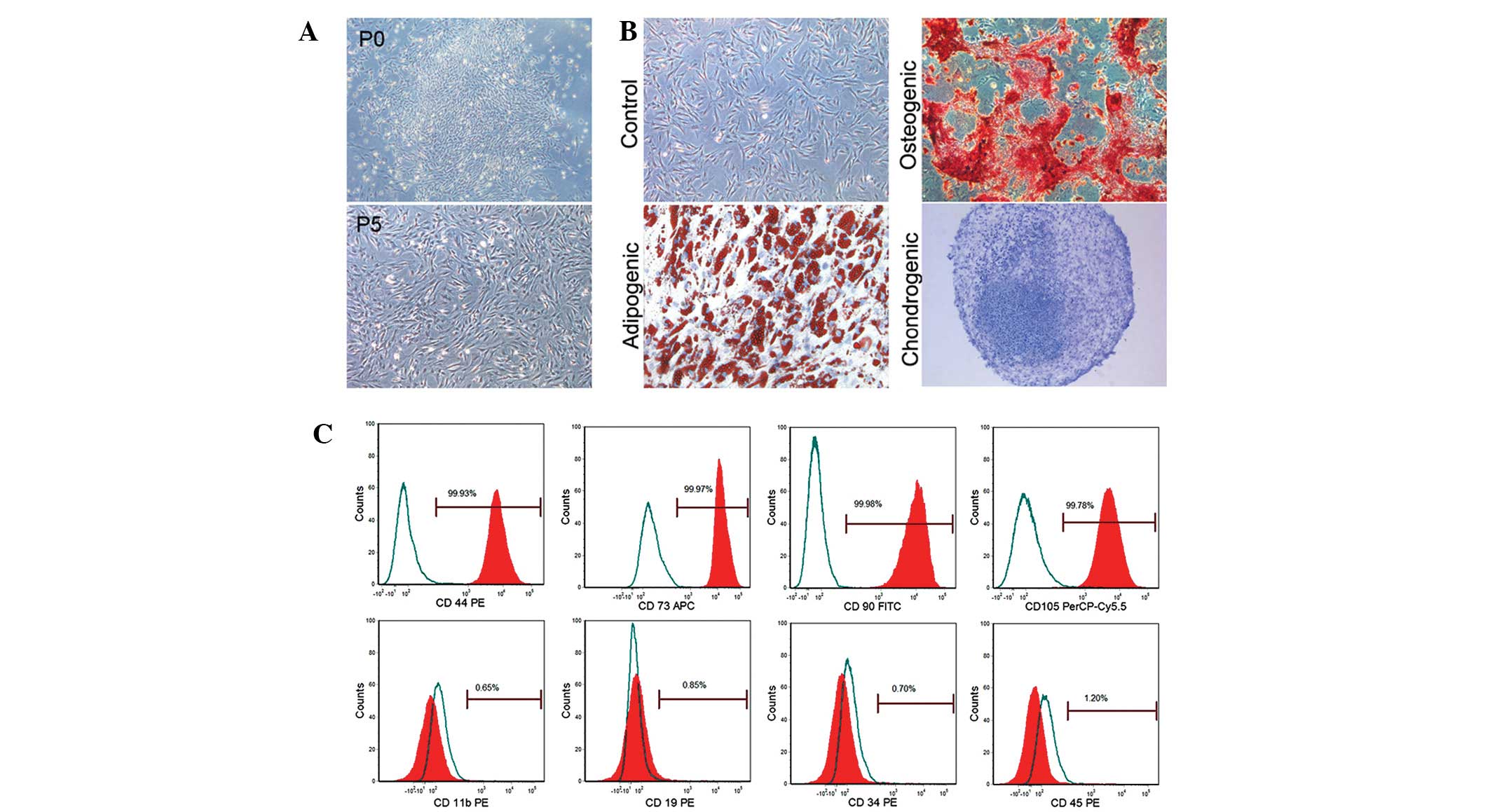
Characterization of BMSCs
A large number of nucleated cells were attached to
the bottom of the plates under a light microscope. These cells
demonstrated fibroblast-like morphology on days 5–6 and cell
colonies were found on days 7–10 (Fig.
1A). These cells gradually aged after passaging 6–7 times, as
identified by an increased cell size, intracellular fine
particulate matter and a decrease in cell proliferation. These
cells demonstrated excellent multi-lineage plasticity and
successfully differentiated into adipocytes, osteoblasts and
chondrocytes following in vitro adipogenic, osteogenic and
chondrogenic differentiation induction for 21, 14 and 28 days,
respectively (Fig. 1B). These
cells were identified by BMSC markers. Flow cytometry identified
that these cells were markedly positive for CD44, CD73, CD90 and
CD105 (99.93, 99.97, 99.98, 99.78%, respectively) but negative for
CD11b, CD19, CD34, and CD45 (0.65, 0.85, 0.70 and 1.20%,
respectively; Fig. 1C).
SDF-1 expression in damaged pancreas
To investigate the effect of the SDF-1/CXCR4 axis on
the migration of BMSCs, the SDF-1 expression in damaged pancreas
was measured and observed at different time points following AP
induction. As summarized in Fig.
2D, in the control group, pancreatic tissues exhibited a low
SDF-1 expression on the cell membrane and in the cytoplasm. The
SDF-1 expression was gradually increased following AP induction,
peaking on days 5–7, however declining from day 10. The SDF-1
protein expression as determined by western blot analysis was
markedly increased in the inflammatory area as compared with the
control group, which was consistent with findings in the
immunohistochemistry assay. The SDF-1/β-actin ratio on day 7 was
the highest, being 8.6-fold greater than that in the control group.
The SDF-1/β-actin ratio in AP group on days 3, 5, 7, 10 was
significantly different from that in control group (P<0.001;
Fig. 2A and B).
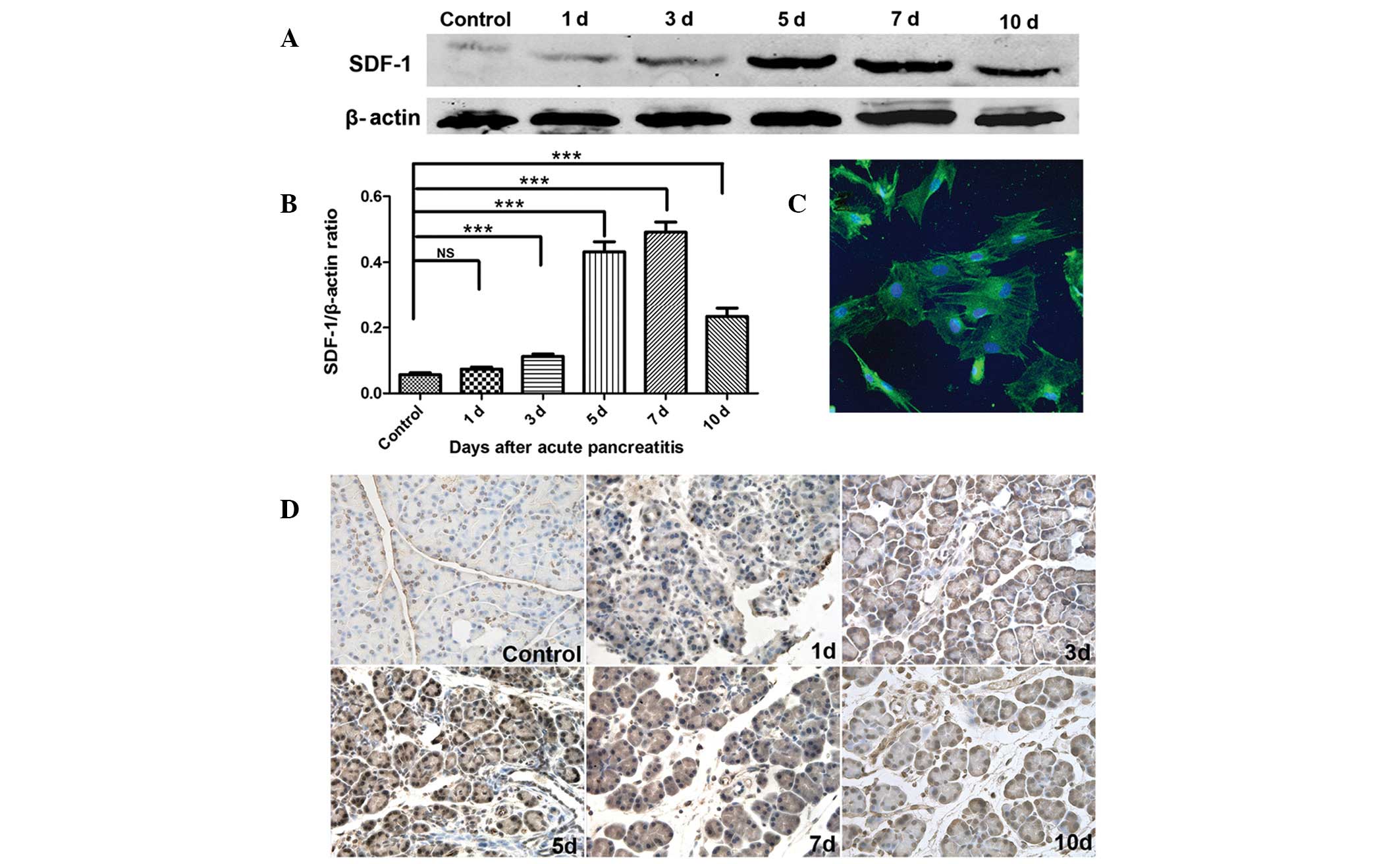 | Figure 2SDF-1 expression in pancreatic tissues
and CXCR4 expression in BMSCs. (A) SDF-1 protein expression in
damaged pancreas detected by western blot analysis at different
time points. (B) Determination of band density, SDF-1/β-actin
ratios were significantly higher on days 3, 5, 7 and 10 as compared
with the control group (P<0.001); (C) CXCR4 expression on BMSCs
was observed by confocal microscopy. Green, CXCR4; blue, DAPI
positive nuclei, original magnification, ×500. (D) SDF-1 expression
in damaged pancreas detected by immunohistochemistry at different
time points. ***P<0.001. ns, not significant; BMSCs,
bone marrow mesenchymal stem cells; CXCR4, CXC-chemokine receptor
4; SDF-1, stromal cell-derived factor-1. |
CXCR4 expression on BMSCs
BMSCs were treated with anti-CXCR4 antibody for 2 h
and then the CXCR4 expression on BMSCs was detected using confocal
microscopy. Five fields were randomly selected for analysis, and
the results demonstrated that >90% of BMSCs expressed CXCR4,
with certain cells having a particularly high expression (Fig. 2C).
SDF-1 induces BMSC migration in
vitro
Our results revealed that SDF-1 induced BMSC
migration in a dose-dependent manner, and that maximum migration
was observed following treatment with SDF-1 at 100 ng/ml, which
indicates that SDF-1 is important in BMSC migration (Fig. 3A and B). The absorption at 405 nm
following treatment with SDF-1 at 100 ng/ml was significantly
increased as compared with the other groups (P<0.001; Fig. 3C). In addition, SDF-1-induced
migration was markedly inhibited when BMSCs were pretreated with
anti-CXCR4 antibody or AMD3100 (Fig.
3D and E). The absorption at 405 nm in the anti-CXCR4 group and
AMD3100 group was significantly decreased as compared with the
SDF-1 group (P<0.001). However, there was no difference between
the anti-CXCR4 group and AMD3100 group (P>0.05; Fig. 3F), which indicate that the
inhibitory effect of anti-CXCR4 antibody is similar to that of
AMD3100.
SDF-1/CXCR4 axis promotes BMSCs migration
into damaged pancreas and pancreatic repair
In order to trace the cells in vivo, CM-Dil
was utilized to label migrating BMSCs. Results demonstrated >95%
of BMSCs were successfully labeled by CM-Dil (Fig. 4A). To investigate the effect of
CM-Dil-labeled BMSCs in damaged pancreas, frozen sections were
prepared as previously described. The results demonstrated CM-Dil
positive BMSCs in the BMSCs and anti-CXCR4 groups, however not in
the AP and control groups. Concurrently, at each time point, the
number of CM-Dil-labeled BMSCs in the anti-CXCR4 group was
significantly lowered as compared with the BMSCs group (Fig. 4B). Cells were counted in five
randomly selected fields at a high magnification and the total
number of cells per section was determined. The results revealed
that blocking CXCR4 expression on BMSCs significantly inhibited the
migration of BMSCs towards the damaged pancreas (Fig. 4C).
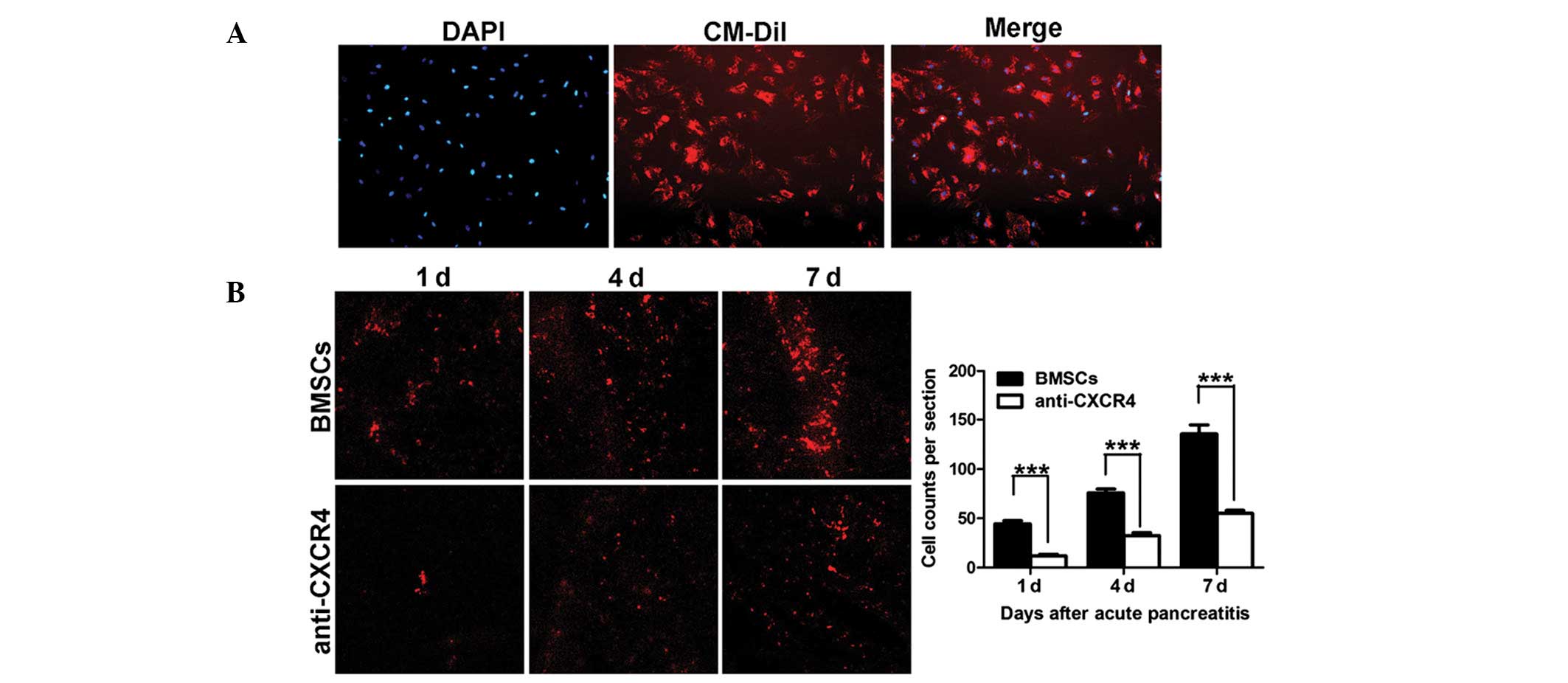 | Figure 4Migration of transplanted BMSCs. (A)
CM-Dil-labeled BMSCs. Red, CM-Dil; blue, DAPI positive nuclei.
Magnification, ×100. (B) Migration of CM-Dil-labeled BMSCs in the
pancreas determined by using confocal microscopy, 25× oil; (C) Cell
counting of migrated BMSCs. The number of migrated BMSCs was
significantly declined in the anti-CXCR4 group as compared with the
BMSCs group at different time points (***P<0.001).
CM-Dil, chloromethylbenzamido-1,
1′-dioctadecyl-3,3,3′3′-tetramethylindo-carbocyanine perchlorate;
BMSCs, bone marrow mesenchymal stem cells; CXCR4, CXC-chemokine
receptor 4. |
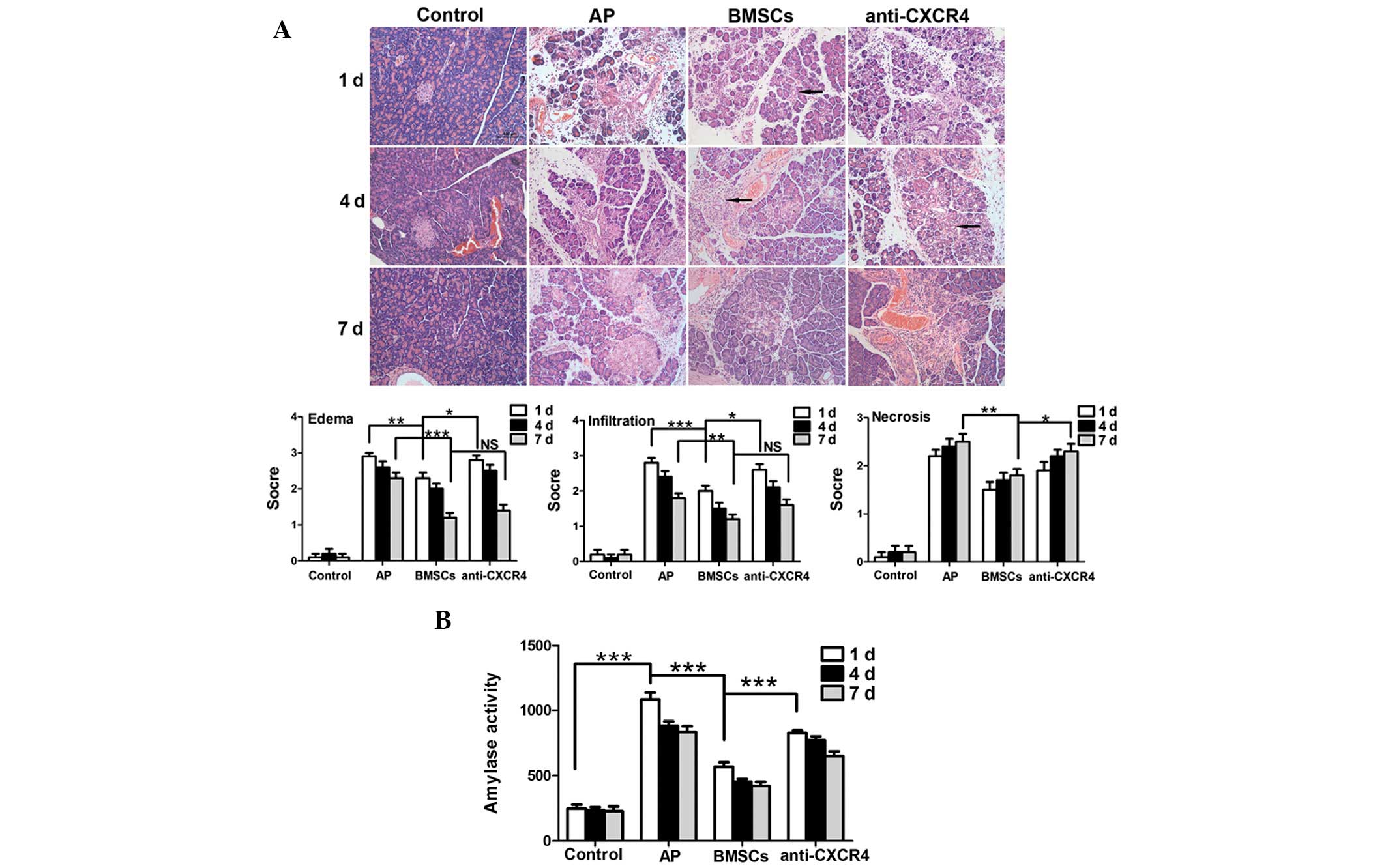
Serum amylase analysis revealed that the amylase
activity in the AP group was significantly higher compared with the
control group on day 1 (P<0.001). Amylase activity appeared to
decrease after BMSCs were transplanted and was significantly
decreased as compared with the AP group (P<0.001). Concurrently,
serum amylase activity in the anti-CXCR4 group was markedly higher
than that in the BMSC group (P<0.001; Fig. 5B). The results of HE staining were
consistent with the change in serum amylase activity, and revealed
that the BMSCs could reduce pancreatic edema, hemorrhage and
necrosis. This effect was compromised following pre-treatment with
the anti-CXCR4 antibody. Furthermore, a large number of tubular
complexes were observed during pancreatic repair in the BMSC group
(Fig. 5A).
Discussion
BMSCs are specialized adult bone marrow stem cells
that have a characteristically potent ability to differentiate. In
the present study, BMSCs were obtained from SD rats and cultured.
Initially, the cell purity was at a low level. The confounding
cells were gradually eliminated by refreshing the medium,
controlling the time of trypsin digestion and passaging cells. The
cell morphology and results of flow cytometry and differentiation
induction were consistent with previously reported data (19,20),
according to the characteristics of MSCs (21).
Several studies have demonstrated that SDF-1 is
expressed by numerous cells, including vascular endothelial cells
(22), acinar cells (23), fibroblasts (24) and cardiomyocytes (13), and that hypoxia upregulates SDF-1
expression. Ceradini et al (25) identified that hypoxia upregulated
SDF-1 expression by activating hypoxia-inducible factor-1 (HIF-1),
to promote the migration of CXCR4-positive progenitor cells into
ischemic tissues. Similarly, Lerman et al (26) confirmed that hypoxia-induced SDF-1
expression was significantly reduced when the HIF-1 gene was
silenced. The pancreatic tissues in AP are ischemic and hypoxic,
and SDF-1 expression in the injured pancreas gradually increased
over time, peaked on days 5–7 and began to decline on day 10, which
confirmed that SDF-1 expression in injured pancreas was
significantly higher than those in normal pancreatic tissues. In
addition, SDF-1 is secreted by the proliferative acinar cells,
vascular endothelial cells and fibroblasts on days 5–7 following AP
induction, which attributed to the peak SDF-1 expression. Jung
et al (7) reported the
number of BMSCs migrated to the damaged pancreas in SAP was larger
than that in mild AP (MAP). The pancreatic ischemia and hypoxia in
SAP were more severe than in MAP, leading to more BMSCs migration
with increased SDF-1 expression in SAP rats.
Our in vitro investigations demonstrated
there was a positive correlation between BMSC migration and SDF-1
concentration. Maximum migration was observed when the SDF-1
concentration was 100 ng/ml, so our results were consistent with
data previously reported (13,27).
Following neutralization or inhibition of SDF-1 (anti-CXCR4
antibody and AMD3100), the migration of BMSCs was significantly
inhibited, which confirms that SDF-1/CXCR4 axis is important in
BMSCs migratory behavior.
Numerous studies have reported that BMSCs utilize
the SDF-1/CXCR4 axis to migrate to damaged tissues in several
pathological conditions, including myocardial ischemia (13), wound area (28), bone fracture (29) and cerebral ischemia (30). The SDF-1/CXCR4 axis links stem cell
migration to cell homing (31).
This study investigated whether BMSCs migrated to the damaged
pancreas in AP and whether the SDF-1/CXCR4 axis was critical in
this process. Firstly, our results revealed >90% of BMSCs
expressed CXCR4 as demonstrated in immunofluorescence staining,
which was consistent with previously reported data (28,32).
In our in vivo investigations, the BMSCs successfully
migrated to the damaged pancreas in the SAP group. Of note, this
effect was significantly inhibited when the SDF-1/CXCR4 axis was
blocked. Thus, we hypothesize that the SDF-1/CXCR4 axis is a key
mediator in BMSC migration to the damaged pancreas. However,
CM-Dil-labeled BMSCs were not completely absent in the anti-CXCR4
group. Thus, other cytokines may also be important regulators of
BMSC migratory behavior. Several studies have revealed that the
macrophage chemoattractant protein-1 (MCP-1) (33) and hepatocyte growth factor (HGF)
(34) are able to promote MSC
migration into malignant gliomas, confirming the potential
applications of genetically modified MSCs for cancer gene therapy.
Xiao et al (35) also
reported that TNF-α could enhance the ability of BMSCs to migrate
into ischemic tissues.
Our results demonstrated that transplanted BMSCs
reduce edema, inflammation and serum amylase activity in the
injured pancreas and act to improve necrosis. There are numerous
inflammatory factors involved in the progression of SAP, which
trigger the activation of inflammatory cascades and may ultimately
result in fatality. A recent study revealed that following BMSC
transplant into SAP animal models, pro-inflammatory cytokine levels
significantly reduced, while anti-inflammatory cytokines levels
increased. They determined that this effect facilitated the control
of pancreatic inflammation and repair of the damaged tissues
(7). Furthermore, Yang et
al (8) postulated that the
paracrine factors of MSCs are important in cellular differentiation
and thus in the repair processes of SAP. Our findings revealed the
expression of numerous tubular complexes in the damaged pancreas
when BMSCs were transplanted. Several studies have indicated that
tubular complexes represent the beginning of regeneration and
repair in the injured pancreas (36). As a result, we have hypothesized
that BMSCs may contribute to pancreatic repair by stimulating the
generation of tubular complexes, however the exact mechanism
underlying this effect remains elusive and requires further
investigation.
In summary, our findings confirmed that damaged
pancreatic tissues in AP exhibit enhanced SDF-1 expression, and
that the SDF-1/CXCR4 axis promotes the migration of transplanted
BMSCs to facilitate in the reparative process that combat the
disease.
Acknowledgements
This study was partially supported by grants from
the National Natural Science Foundation of China (no. 81170436) and
the Shanghai Municipal Health Bureau (no. 2010015).
References
|
1
|
Granger J and Remick D: Acute
pancreatitis: models, markers, and mediators. Shock. 24(Suppl 1):
45–51. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Berezina TL, Zaets SB, Mole DJ, Spolarics
Z, Deitch EA and Machiedo GW: Mesenteric lymph duct ligation
decreases red blood cell alterations caused by acute pancreatitis.
Am J Surg. 190:800–804. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Carvalho PH, Daibert AP, Monteiro BS, et
al: Differentiation of adipose tissue-derived mesenchymal stem
cells into cardiomyocytes. Arq Bras Cardiol. 100:82–89.
2013.PubMed/NCBI
|
|
4
|
Pankajakshan D, Kansal V and Agrawal DK:
In vitro differentiation of bone marrow derived porcine mesenchymal
stem cells to endothelial cells; J Tissue Eng Regen Med. May
18–2012.(Epub ahead of print).
|
|
5
|
Phuc PV, Nhung TH, Loan DT, Chung DC and
Ngoc PK: Differentiating of banked human umbilical cord
blood-derived mesenchymal stem cells into insulin-secreting cells.
In Vitro Cell Dev Biol Anim; 47:54–63. 2011.PubMed/NCBI
|
|
6
|
Tu XH, Song JX, Xue XJ, et al: Role of
bone marrow-derived mesenchymal stem cells in a rat model of severe
acute pancreatitis. World J Gastroenterol. 18:2270–2279. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jung KH, Song SU, Yi T, et al: Human bone
marrow-derived clonal mesenchymal stem cells inhibit inflammation
and reduce acute pancreatitis in rats. Gastroenterology.
140:998–1008. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang B, Bai B, Liu CX, et al: Effect of
umbilical cord mesenchymal stem cells on treatment of severe acute
pancreatitis in rats. Cytotherapy. 15:154–162. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
McIver SC, Loveland KL, Roman SD, Nixon B,
Kitazawa R and McLaughlin EA: The chemokine CXCL12 and its receptor
CXCR4 are implicated in human seminoma metastasis. Andrology.
1:517–529. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mukherjee D and Zhao J: The role of
chemokine receptor CXCR4 in breast cancer metastasis. Am J Cancer
Res. 3:46–57. 2013.PubMed/NCBI
|
|
11
|
Du L, Yang P and Ge S: Stromal
cell-derived factor-1 significantly induces proliferation,
migration, and collagen type I expression in a human periodontal
ligament stem cell subpopulation. J Periodontol. 83:379–388. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Theiss HD, Vallaster M, Rischpler C, et
al: Dual stem cell therapy after myocardial infarction acts
specifically by enhanced homing via the SDF-1/CXCR4 axis. Stem Cell
Res. 7:244–255. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu J, Li M, Qu Z, Yan D, Li D and Ruan Q:
SDF-1/CXCR4-mediated migration of transplanted bone marrow stromal
cells toward areas of heart myocardial infarction through
activation of PI3K/Akt. J Cardiovasc Pharmacol. 55:496–505.
2010.PubMed/NCBI
|
|
14
|
Shen LH, Li Y, Chen J, et al: Therapeutic
benefit of bone marrow stromal cells administered 1 month after
stroke. J Cereb Blood Flow Metab. 27:6–13. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Togel F, Isaac J, Hu Z, Weiss K and
Westenfelder C: Renal SDF-1 signals mobilization and homing of
CXCR4-positive cells to the kidney after ischemic injury. Kidney
Int. 67:1772–1784. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Perides G, van Acker GJ, Laukkarinen JM
and Steer ML: Experimental acute biliary pancreatitis induced by
retrograde infusion of bile acids into the mouse pancreatic duct.
Nat Protoc. 5:335–341. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Weir C, Morel-Kopp MC, Gill A, Tinworth K,
Ladd L, Hunyor N and Ward C: Mesenchymal stem cells: isolation,
characterisation and in vivo fluorescent dye tracking. Heart Lung
Circ. 17:395–403. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wittel UA, Wiech T, Chakraborty S, et al:
Taurocholate-induced pancreatitis: a model of severe necrotizing
pancreatitis in mice. Pancreas. 36:e9–e21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kemp KC, Hows J and Donaldson C: Bone
marrow-derived mesenchymal stem cells. Leuk Lymphoma. 46:1531–1544.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin RZ, Moreno-Luna R, Zhou B, Pu WT and
Melero-Martin JM: Equal modulation of endothelial cell function by
four distinct tissue-specific mesenchymal stem cells. Angiogenesis.
15:443–455. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dominici M, Le Blanc K, Mueller I, et al:
Minimal criteria for defining multipotent mesenchymal stromal
cells. The International Society for Cellular Therapy position
statement. Cytotherapy. 8:315–317. 2006. View Article : Google Scholar
|
|
22
|
Li M, Yu J, Li Y, et al: CXCR4 positive
bone mesenchymal stem cells migrate to human endothelial cell
stimulated by ox-LDL via SDF-1alpha/CXCR4 signaling axis. Exp Mol
Pathol. 88:250–255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu L, Cecil J, Peng SB, et al:
Identification and expression of novel isoforms of human stromal
cell-derived factor 1. Gene. 374:174–179. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Toksoy A, Müller V, Gillitzer R and
Goebeler M: Biphasic expression of stromal cell-derived factor-1
during human wound healing. Br J Dermatol. 157:1148–1154. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ceradini DJ, Kulkarni AR, Callaghan MJ, et
al: Progenitor cell trafficking is regulated by hypoxic gradients
through HIF-1 induction of SDF-1. Nat Med. 10:858–864. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lerman OZ, Greives MR, Singh SP, et al:
Low-dose radiation augments vasculogenesis signaling through
HIF-1-dependent and -independent SDF-1 induction. Blood.
116:3669–3676. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ryu CH, Park SA, Kim SM, et al: Migration
of human umbilical cord blood mesenchymal stem cells mediated by
stromal cell-derived factor-1/CXCR4 axis via Akt, ERK, and p38
signal transduction pathways. Biochem Biophys Res Commun.
398:105–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu X, Zhu F, Zhang M, et al: Stromal
cell-derived factor-1 enhances wound healing through recruiting
bone marrow-derived mesenchymal stem cells to the wound area and
promoting neovascularization. Cells Tissues Organs. 197:103–113.
2013. View Article : Google Scholar
|
|
29
|
Kitaori T, Ito H, Schwarz EM, et al:
Stromal cell-derived factor 1/CXCR4 signaling is critical for the
recruitment of mesenchymal stem cells to the fracture site during
skeletal repair in a mouse model. Arthritis Rheum. 60:813–823.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Y, Deng Y and Zhou GQ:
SDF-1alpha/CXCR4-mediated migration of systemically transplanted
bone marrow stromal cells towards ischemic brain lesion in a rat
model. Brain Res. 1195:104–112. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Miller RJ, Banisadr G and Bhattacharyya
BJ: CXCR4 signaling in the regulation of stem cell migration and
development. J Neuroimmunol. 198:31–38. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kucia M, Reca R, Miekus K, et al:
Trafficking of normal stem cells and metastasis of cancer stem
cells involve similar mechanisms: pivotal role of the SDF-1-CXCR4
axis. Stem Cells. 23:879–894. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu F, Shi J, Yu B, Ni W, Wu X and Gu Z:
Chemokines mediate mesenchymal stem cell migration toward gliomas
in vitro. Oncol Rep. 23:1561–1567. 2010.PubMed/NCBI
|
|
34
|
Vogel S, Peters C, Etminan N, et al:
Migration of mesenchymal stem cells towards glioblastoma cells
depends on hepatocyte-growth factor and is enhanced by
aminolaevulinic acid-mediated photodynamic treatment. Biochem
Biophys Res Commun. 431:428–432. 2013. View Article : Google Scholar
|
|
35
|
Xiao Q, Wang SK, Tian H, et al: TNF-alpha
increases bone marrow mesenchymal stem cell migration to ischemic
tissues. Cell Biochem Biophys. 62:409–414. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kitamura T, Asanuma N, Inaba M, et al:
Regeneration of tubular complex is promoted by a free space.
Pancreas. 30:174–179. 2005. View Article : Google Scholar : PubMed/NCBI
|