Introduction
Cellular senescence is a process of cellular aging
in which primary cells in culture lose their ability to divide
(1). It has been demonstrated that
senescence is associated with cellular dysfunction and occurs in
vivo in cardiovascular diseases associated with age, such as
atherosclerosis (2,3). In endothelial cells, the
senescence-induced loss of replicative capacity destroys the
integrity of the endothelium and impairs successful angiogenesis
(4,5).
In several recent studies, protein-protein
interactions have been demonstrated to be important in the
molecular recognition and functional modulation of proteins in
numerous signal transduction pathways (6,7). The
kinase non-catalytic C-lobe domain (KIND) is a putative
protein-protein interaction module (8). Four KIND-containing proteins have
been reported: Spir-2 (an actin-nucleation factor), PTPN13 (a
protein tyrosine phosphatase), FRMPD2 (a scaffold protein) and the
Ras guanine exchange factor (RasGEF), very-KIND [v-KIND, also
termed kinase noncatalytic C-lobe domain containing 1, (KNDC1)]
(9,10). v-KIND, a brain-specific Ras guanine
nucleotide exchange factor, has two KIND isoforms, KIND1 and KIND2,
whereas the other three proteins have only one. A previous study
demonstrated that v-KIND interacts with the high-molecular weight
microtubule associated protein 2 (MAP2), a dendritic protein that
drives negative regulation of neuronal dendrite growth. v-KIND
overexpression suppresses the growth and branching of the dendrites
of hippocampal neurons and cerebellar granule cells, whereas
knockdown of endogenous v-KIND expression promotes dendrite growth.
These findings suggest that v-KIND regulates a signaling pathway
that links Ras and MAP2 to control dendrite growth (11). However, its role in vascular cell
biology has not been investigated to date.
In the present study, the expression of KNDC1 with
increasing age, as well as the effects of its depletion by RNA
interference on senescence were investigated in human umbilical
vein endothelial cells (HUVECS).
Materials and methods
Chemicals and reagents
Trypsin was purchased from Invitrogen Life
Technologies (Beijing, China). Antibodies against extracellular
signal-regulated kinase (ERK) 1/2, phospho-ERK1/2 (Thr202/Tyr204;
rabbit polyclonal), p38, phospho-p38 (Thr180/Tyr182; mouse
monoclonal), stress-activated protein kinase (SAPK)/c-Jun
N-terminal kinase (JNK), phospho-SAPK/JNK (Thr183/Tyr185; rabbit
monoclonal), endothelial nitric oxide synthase (eNOS; rabbit
polyclonal), vascular cell adhesion molecule (VCAM-1; rabbit
polyclonal), intercellular adhesion molecule (ICAM-1; rabbit
polyclonal), p53, phospho-p53 (Ser46; rabbit polyclonal), p21
(mouse monoclonal) and p16 (rabbit polyclonal) were purchased from
Cell Signaling Technology, Inc. (Cell Signaling Technology,
Danvers, MA, USA). Antibodies against KNDC1 and β-actin were
purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA). Secondary antibodies
against rabbit and mouse were purchased from Cell Signaling
Technology, Inc. The pre-stained protein marker was purchased from
New England Biolabs, Ltd. (Beijing, China). Luminol reagent and
polyvinylidene fluoride (PVDF) membrane for western blotting were
purchased from Millipore (Millipore, Billerica, MA, USA).
Cells and cell culture
HUVECs were isolated from the umbilical cords of
newborns supplied by Tongren Hospital (Beijing, China) and grown in
M199 cell medium (Hyclone, Logan, UT, USA) containing 100 mg/ml
streptomycin, 100 IU/ml penicillin, 40 μg/ml endothelial cell
growth supplement and 20% fetal bovine serum (Hyclone) at 37°C in a
humidified atmosphere of 95% air and 5% CO2. The cells
were passaged at 80–90% confluence at a ratio of 1:2 and used for
experiments at passages (P) 3–5.
Transfections
The fourth-passage HUVECs were transfected at 70%
confluence for 24 h with 20 nM small interfering (si)RNAs targeting
human KNDC1 [KNDC1-siRNA1 was obtained from Santa Cruz
Biotechnology, Inc. (SC-90387); KNDC1-siRNA2 was obtained from
Invitrogen Life Technologies and the sense sequence was as follows:
CAUCCAGGAGGAAUUUGCCUUUGAU]. A non-targeting control pool (NT-siRNA;
Santa Cruz Biotechnology, Inc.) was also used. Transfections were
performed using Hyperfect reagent (Qiagen, Shanghai, China)
according to the manufacturer’s instructions. After 4 h, fresh
medium was added and the cells were cultured for a further 72-h
period prior to analysis.
Senescence-associated β-galactosidase
(SA-β-gal) staining
Endothelial cells were transfected with KNDC1-siRNA
or NT-siRNA. Following incubation for 72 h, the cells were washed
twice with phosphate-buffered saline (PBS) and then fixed for 5 min
with PBS containing 2% formaldehyde and 0.2% glutaraldehyde. The
cells were then incubated at 37°C for 10 h in a staining solution
of 40 mM citric acid, sodium phosphate, pH 6.0, 1 mg/ml
5-bromo-4-chloro-3-isolyl-β-d-galactoside (X-gal; Sigma, Shanghai,
China), 5 mM potassium ferrocyanide, 5 mM potassium ferricyanide,
150 mM NaCl and 2 mM MgCl2. SA-β-gal-positive cells were
observed by microscopy (CKX31; Olympus, Beijing, China) and over
400 cells were counted in three independent fields as described
previously (12).
RNA expression analysis
Cellular RNA was extracted with TRIzol reagent
(Invitrogen Life Technologies) according to the manufacturer’s
instructions. RNA expression was measured by quantitative
polymerase chain reaction (qPCR) with the appropriate primers using
the one step SYBR PrimeScript RT-PCR kit (Takara Bio, Inc., Dalian,
China). A 20 μl PCR reaction mixture was initially amplified and
primer pairs for KNDC1 were obtained from Santa Cruz Biotechnology,
Inc. Primer pairs for β-actin were synthesized by Shanghai
Bioengineering Company (Shanghai, China). The PCR was run on an
iCycler (Bio-Rad, Hercules, CA, USA). The thermal profile for SYBR
qPCR was 42°C for 5 min, 95°C for 10 sec followed by 40
amplification cycles of 95°C for 5 sec and 60°C for 20 sec.
Relative mRNA expression levels were calculated by the comparative
cycle threshold (CT) method, using the CT values obtained for
β-actin as internal references.
Western blot analysis
Endothelial cells (7.5×105) were washed
with ice-cold PBS and scraped off the flask into 100 μl lysis
buffer containing 25 mM Tris-HCl, pH 6.8, 1% sodium dodecylsulfate,
1 mM phenylmethylsulphonyl fluoride and protease inhibitor cocktail
(Sigma). The resultant lysates were further disrupted by sonication
for 10 sec at an amplitude of 35% using a VCX 500 Ultrasonic
Processor (Sonics & Materials, Newtown, CT, USA) and then
centrifuged at 12,000 × g for 20 min to remove particulate
material. Proteins (30 μg) were fractionated by SDS-PAGE and
transferred to PVDF membranes. The membranes were incubated as
previously described (12).
Cell cycle analysis
Cells (4×105) were plated in 25
cm2 flasks and cultured in M199 medium. Following
incubation for 24 h, the cells were transfected with KNDC1-siRNA or
NT-siRNA. The cells were harvested at 72 h following transfection,
washed twice in salt buffer (1% bovine serum albumin and 0.5%
sodium azide in PBS) and fixed in 70% ethanol at 4°C overnight.
After washing twice in salt buffer, the cells were stained in 50
μg/ml propidium iodide (PI) solution (Sigma) containing 100 μg/ml
RNase for 1 h. Then, the cells were transferred to flow cytometry
tubes with filters for cell cycle analysis.
Capillary tube network formation
Endothelial capillary tube network formation was
assessed using Matrigel (BD Biosciences, Franklin Lakes, NJ, USA).
Each well of the 96-well culture plates was coated with 100 μl
Matrigel and immediately incubated at 37°C for 1 h to allow gel
formation. Upon gelation, transfected cells were seeded onto the
Matrigel-coated 96-well plates (3×104 cells/well) in 100
μl M199 medium. Following incubation for 24 h, tubule formation was
analyzed under an Olympus inverted microscope (CKX31; Olympus,
Beijing, China) at a magnification of ×40. Images were captured
under phase contrast using an Olympus digital camera and analyzed
with the Image J software (National Institutes of Health, Bethesda,
MD, USA). Tubule length was determined by drawing a line along each
tubule and measuring the line length in pixels. Branch points were
counted manually.
Statistical analysis
Values are presented as the mean ± standard
deviation. Statistical analysis was performed using SPSS 11.0
(SPSS, Inc., Chicago, IL, USA). Results were evaluated by t-test or
one-way analysis of variance followed by Bonferroni’s post-hoc
tests as appropriate. A value of P<0.05 was considered to
indicate a statistically significant difference.
Results
Senescence-like phenotype and increased
transcription and expression of KNDC1 in HUVECs with increasing
passage number
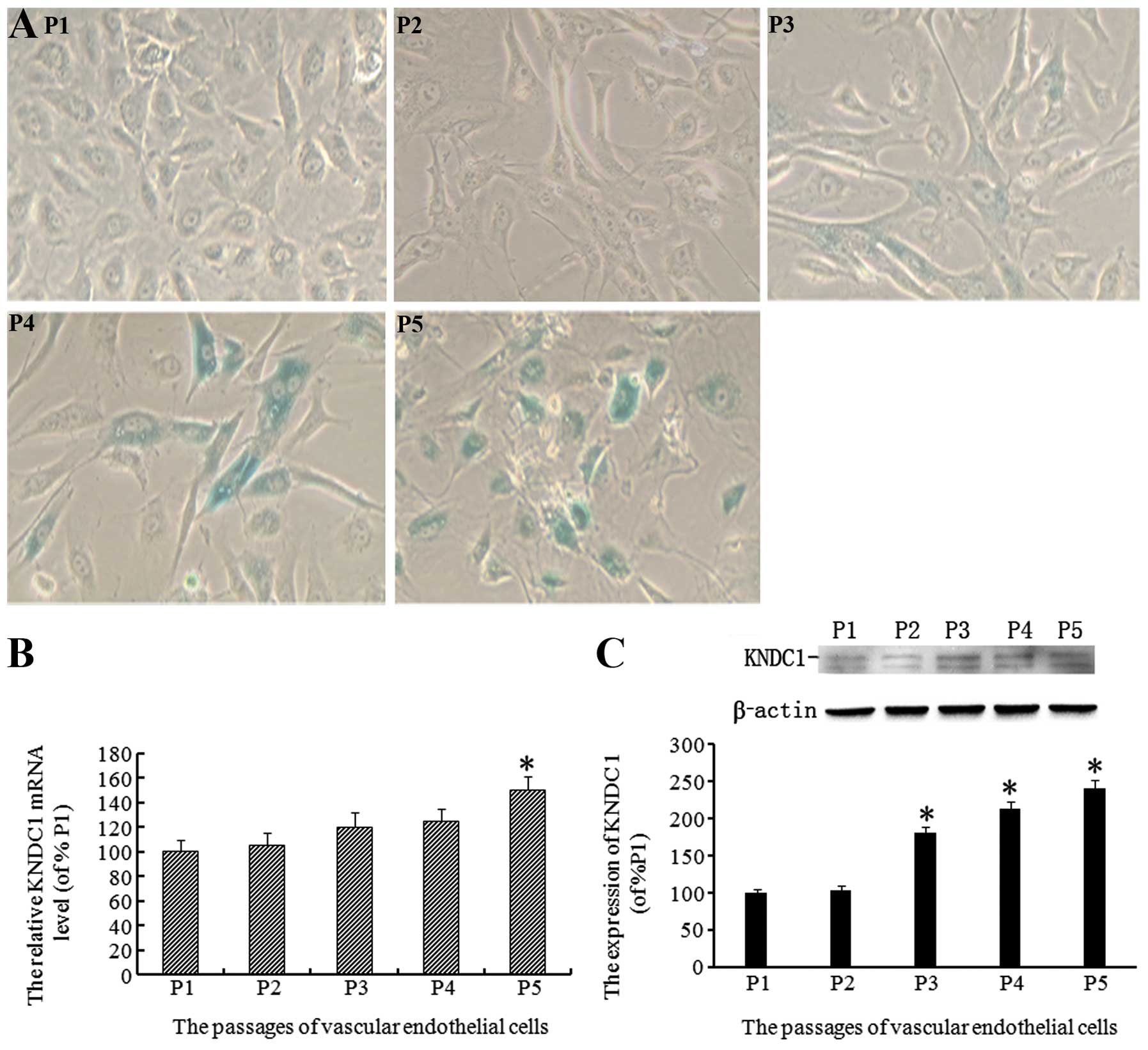
When HUVECs are cultured in vitro, their
growth slows down and proliferation stops at the fifth passage. In
the present study, it was observed that with increasing passage
number, the number of aging HUVEC cells increased, as demonstrated
by SA-β-gal staining. The percentages of aging cells at different
passages were ~2.0±2.0% (P1), 8.0±3.0% (P2), 15.0±3.0% (P3),
35.0±6.0% (P4) and 62.0±10.0% (P5; Fig. 1A). To investigate whether KNDC1 was
associated with the senescence of normal cells, the expression
levels of KNDC1 in HUVECs at different passages were examined by
qPCR and western blot analysis. The levels of KNDC1 mRNA exhibited
a statistically significant increase in HUVECs at P1–P5 (Fig. 1B). Similarly, the levels of KNDC1
protein also demonstrated a statistically significant increase in
P1–P5 HUVECs (Fig. 1C).
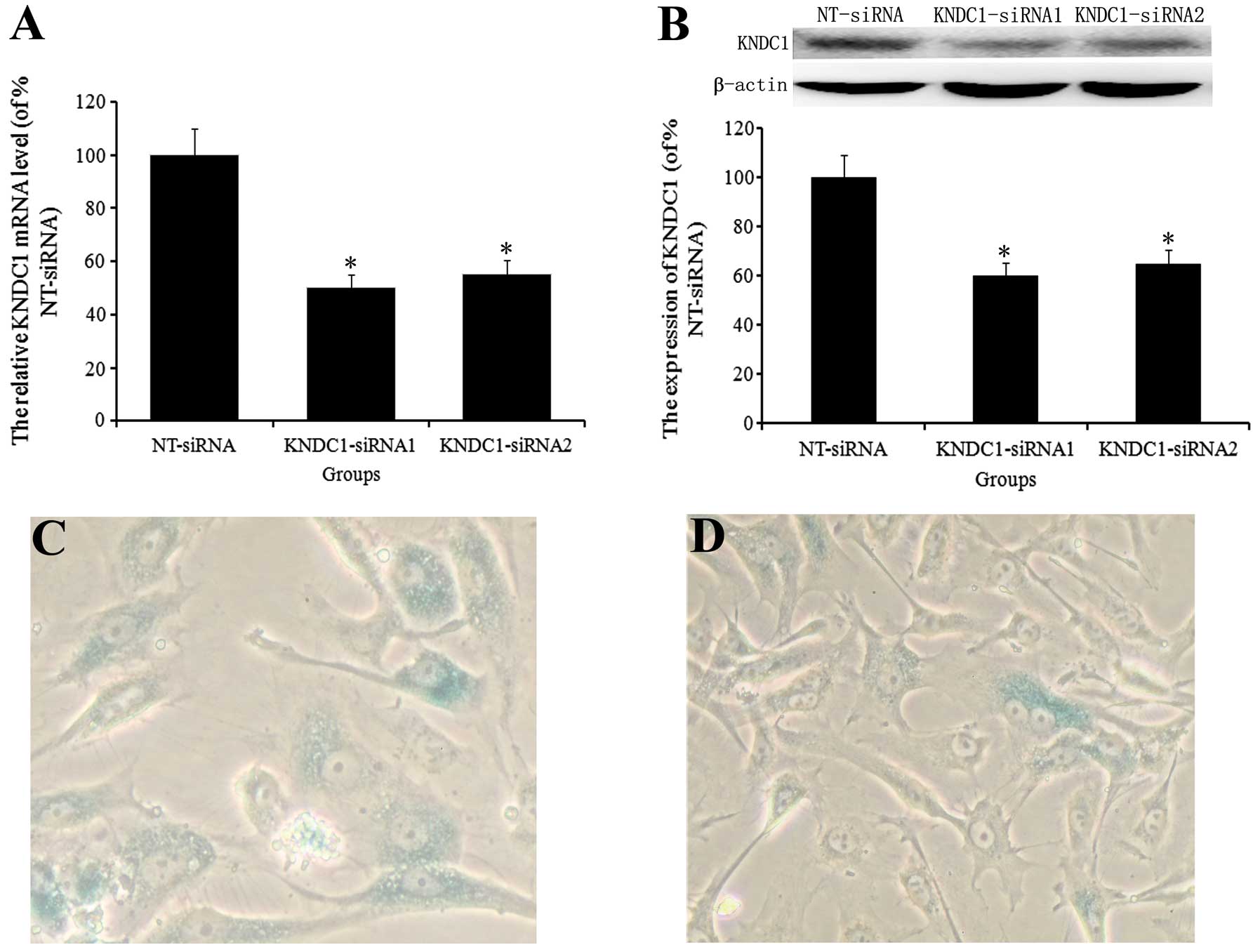
Partial reversal of cellular senescence
in aging HUVECs by KNDC1 knockdown
Aging cells express SA-β-gal, are resistant to
mitogen-induced proliferation and have a characteristically
enlarged and flattened morphology. In the present study, HUVECs at
P4 exhibited senescence phenotypes that distinguished them from
early passage cells. To investigate the role of KNDC1 in cellular
senescence, the levels of KNDC1 mRNA and protein in HUVECs at P4
were downregulated by gene silencing with KNDC1-siRNA1/2.
Transfection with KNDC1-siRNA1/2 caused a ~50% decrease in KNDC1
levels (Fig. 2A and B). In
subsequent studies, only KNDC1-siRNA1 was used, as KNDC1-siRNA1 and
-2 had the same effect. Following repression of KNDC1 levels in
HUVECs at P4, the size of the cells was similar to the size of P1
cells and the activity of SA-β-gal was decreased compared with that
in NT-siRNA-transfected cells (10.0±3 vs. 38.0±7%; Fig. 2C and D).
Knockdown of KNDC1 affects cell cycle
progression
Replicatively senescent cells are known to undergo
arrest in the G1/G0 phase of the cell cycle. To investigate whether
cell proliferation induced by KNDC1-siRNA impacted the cell cycle,
HUVECs at P4 were transfected with KNDC1-siRNA or NT-siRNA for 72 h
and their DNA content was quantified by flow cytometry with PI
staining. The majority of the cells transfected with NT-siRNA for
72 h had undergone G1 phase arrest, which is one of the typical
phenotypes of cellular senescence. However, the number of cells in
G1 phase was decreased in cells transfected with KNDC1-siRNA
(Fig. 3A and B; Table I).
 | Table ICell cycle distribution of P4
generation HUVECs following KDNC1-siRNA treatment for 72 h
(n=3). |
Table I
Cell cycle distribution of P4
generation HUVECs following KDNC1-siRNA treatment for 72 h
(n=3).
| Percentage (%) |
|---|
|
|
|---|
| Groups | G1 | S | G2/M |
|---|
| NT-siRNA | 94.25±5.6 | 4.75±1.0 | 1.00±0.5 |
| KNDC1-siRNA | 85.78±4.5* | 8.29±1.0* | 5.93±1.0* |
Knockdown of KNDC1 improves capillary
tube network formation
Since senescence is associated with impaired
angiogenic function, it was examined whether KNDC1 knockdown
improved the ability of HUVECs to form capillary tube networks
in vitro. Compared with NT-siRNA-treated cells,
KNDC1-silenced cells formed more developed tubule networks, in
which the mean tubule length (15±5.2 mm vs. 30 mm±7.0) and the
number of branches at each point (3±1.0/point vs. 2±1.0/point)
increased significantly (Fig. 3C and
D).
Alterations in signaling pathways induced
by knockdown of KNDC1
In the present study, the effect of KNDC1 knockdown
on the proliferation of endothelial cells [mitogen-activated
protein kinase (MAPK) signaling pathway], the function of
endothelial cells (eNOS, VCAM-1 and ICAM-1) and the senescence of
endothelial cells (p53, p21 and p16) was investigated. It was
observed that the phosphorylation of ERK, but not of p38 and JNK,
was correlated to the proliferation of endothelial cells induced by
the knockdown of KNDC1. The increased expression of eNOS indicated
that the activity of the endothelial cells increased; however, the
expression levels of VCAM-1 and ICAM-1 did not change compared with
NT-siRNA-transfected control cells. In addition, the
phosphorylation of p53 and the expression of p21 and p16 decreased
significantly compared with NT-siRNA-transfected control cells
(Fig. 4).
 | Figure 4Effect of KNDC1 knockdown on signaling
pathways. HUVECs were transfected with KNDC1-siRNA or NT-siRNA.
Following incubation for 72 h, the expression levels of eNOS,
VCAM-1, ICAM-1, p21, p16 and p-ERK, p38, JNK and p53 were examined
by western blot analysis (n=3). Values are presented as the mean ±
standard deviation. *P<0.05 vs NT-siRNA. KNDC1,
kinase noncatalytic C-lobe domain containing 1; HUVECs, human
umbilical vein endothelial cells; NT-siRNA, non-targeting control
pool; p, phosphorylated; siRNA, small interfering RNA; ERK,
extracellular signal-regulated kinase; JNK, c-Jun N-terminal
kinase; eNOS, endothelial nitric oxide synthase; VCAM-1, vascular
cell adhesion molecule 1; ICAM-1, intercellular adhesion molecule
1. |
Discussion
KNDC1 is a brain-specific Ras guanine nucleotide
exchange factor. Overexpression of KNDC1 suppresses dendritic
extension and branching in hippocampal neurons and cerebellar
granule cells, whereas knockdown of endogenous KNDC1 expression
promotes dendrite growth (11).
However, the effect of KNDC1 on cellular senescence had yet to be
elucidated. Cellular senescence, i.e., the limited ability of
primary human cells to divide when cultured in vitro, is
utilized as a model of biological aging. In common with other
normal diploid cells, HUVECs have a limited capacity to divide
(13). Therefore, in the present
study, HUVECs were cultured in vitro as a model of
biological aging, to investigate the effect of KNDC1 on senescence.
It was observed that with increasing passages, the size of the
senile HUVECs increased and the cells were positively stained with
SA-β-gal. Furthermore, the transcription of the KNDC1 mRNA and
expression of the KNDC1 protein increased with cellular senescence
(Fig. 1). When the expression of
KNDC1 was knocked down with a specific siRNA, the size and the
number of the HUVECs decreased, and there was a statistically
significant decrease in the number of HUVECs stained with SA-β-gal
compared with control NT-siRNA-transfected HUVECs of the same
passage. These results suggested that KNDC1 may be important in the
progression of senescence in HUVECs and that knockdown of KNDC1
delayed this senescence. However, further investigation is required
to determine whether overexpression of KNDC1 may accelerate
senescence in HUVECs.
Cellular senescence is considered to be an
irreversible block of cell cycle progression in populations of
otherwise replication-competent cells (14,15).
The proportion of arrested cells in a population rises with
increasing population doublings, rather than all cells becoming
senescent at once (16,17). In the present study, it was
identified that knockdown of KNDC1 decreased the percentage of
cells in the G0/G1 stage and increased the percentage of cells in
the S and G2/M stages, compared with NT-siRNA-transfected control
HUVECs from the same passage. The change in the percentage cells in
G0/G1 phase was similar to the change in the expression of
KNDC1.
The alterations in morphology of HUVECs transfected
with KNDC1-siRNA was minimal with regard to their function.
Therefore, in order to obtain further insight into the effect of
KNDC1 on angiogenesis, endothelial capillary tube network formation
was investigated. The results demonstrated that when HUVECs were
transfected with KNDC1-siRNA, the mean tubule length and the number
of branch points was significantly increased compared with HUVECs
transfected with NT-siRNA. These results indicated that knockdown
of KNDC1 not only delayed senescence in HUVECs but also improved
endothelial capillary tube network formation.
Following this, the mechanism by which transfection
with KNDC1-siRNA altered HUVEC function was investigated. The MAPK
signaling pathway is associated with cell proliferation (18,19),
eNOS, VCAM-1 and ICAM-1 are required for the function of HUVECs
(20,21) and the p53-p21-p16 signaling pathway
controls cell senescence (22,23).
In the present study, it was demonstrated that the knockdown of
KNDC1 promoted HUVEC proliferation via the ERK signaling pathway,
but not the p38 and JNK transduction cascades. Furthermore,
knockdown of KNDC1 had no effect on the expression of VCAM-1 and
ICAM-1, but increased the expression of eNOS, which induces the
relaxation of blood vessels. In addition, the present study
provided the first evidence, to the best of our knowledge, for the
involvement of KNDC1 in the senescence of human primary endothelial
cells, acting through the p53 signaling pathway. It was
demonstrated that knockdown of KNDC1 delayed senescence in HUVECs
by decreasing the phosphorylation of p53 and the expression of the
associated downstream signaling molecules, p21 and p16.
In conclusion, knockdown of KNDC1 promoted the
proliferation and delayed the senescence of HUVECs. Regarding the
mechanism of action, it was demonstrated that knockdown of KNDC1
increased the percentage of cells in the S and G2/M phases by
inhibiting the p53-p21-p16 signaling pathway. In addition, it was
observed that knockdown of KNDC1 increased the expression of eNOS
and improved capillary tube network formation. Therefore, it is
concluded that knockdown of KNDC1 contributed to delayed
endothelial cell senescence. The results suggested that KNDC1 has
the potential to be a novel target in the development of
pharmacological agents to delay the aging process and extend human
life.
Acknowledgements
This study was supported by a grant from the
National Natural Science Foundation of China (no. 81001439).
References
|
1
|
Huang J, Gan Q, Han L, et al: SIRT1
overexpression antagonizes cellular senescence with activated
ERK/S6k1 signaling in human diploid fibroblasts. PLoS One.
3:e17102008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Krouwer VJ, Hekking LH, Langelaar-Makkinje
M, Regan-Klapisz E and Post JA: Endothelial cell senescence is
associated with disrupted cell-cell junctions and increased
monolayer permeability. Vasc Cell. 4:122012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang JC and Bennett M: Aging and
atherosclerosis: mechanisms, functional consequences, and potential
therapeutics for cellular senescence. Circ Res. 111:245–259. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Erusalimsky JD: Vascular endothelial
senescence: from mechanisms to pathophysiology. J Appl Physiol.
1985.106:326–332. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cardus A, Uryga AK, Walters G and
Erusalimsky JD: SIRT6 protects human endothelial cells from DNA
damage, telomere dysfunction, and senescence. Cardiovasc Res.
97:571–579. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang J, Furuya A, Hayashi K and Furuichi
T: Interaction between very-KIND Ras guanine exchange factor and
microtubule-associated protein 2, and its role in dendrite growth -
structure and function of the second kinase noncatalytic C-lobe
domain. FEBS J. 278:1651–1661. 2011. View Article : Google Scholar
|
|
7
|
Pawson T and Nash P: Assembly of cell
regulatory systems through protein interaction domains. Science.
300:445–452. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ciccarelli FD, Bork P and Kerkhoff E: The
KIND module: a putative signalling domain evolved from the C lobe
of the protein kinase fold. Trends Biochem Sci. 28:349–352. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mees A, Rock R, Ciccarelli FD, et al:
Very-KIND is a novel nervous system specific guanine nucleotide
exchange factor for Ras GTPases. Gene Expr Patterns. 6:79–85. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zeth K, Pechlivanis M, Samol A, Pleiser S,
Vonrhein C and Kerkhoff E: Molecular basis of actin nucleation
factor cooperativity: crystal structure of the Spir-1 kinase
non-catalytic C-lobe domain (KIND)•formin-2 formin SPIR interaction
motif (FSI) complex. J Biol Chem. 286:30732–30739. 2011.PubMed/NCBI
|
|
11
|
Huang J, Furuya A and Furuichi T:
Very-KIND, a KIND domain containing RasGEF, controls dendrite
growth by linking Ras small GTPases and MAP2. J Cell Biol.
179:539–552. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin YJ, Zhen YZ, Wei J, Liu B, Yu ZY and
Hu G: Effects of Rhein lysinate on H2O2-induced cellular senescence
of human umbilical vascular endothelial cells. Acta Pharmacol Sin.
32:1246–1252. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim KS, Kim MS, Seu YB, Chung HY, Kim JH
and Kim JR: Regulation of replicative senescence by insulin-like
growth factor-binding protein 3 in human umbilical vein endothelial
cells. Aging Cell. 6:535–545. 2007. View Article : Google Scholar
|
|
14
|
Jeyapalan JC and Sedivy JM: Cellular
senescence and organismal aging. Mech Ageing Dev. 129:467–474.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mao Z, Ke Z, Gorbunova V and Seluanov A:
Replicatively senescent cells are arrested in G1 and G2 phases.
Aging (Albany NY). 4:431–435. 2012.PubMed/NCBI
|
|
16
|
Passos JF, Saretzki G, Ahmed S, et al:
Mitochondrial dysfunction accounts for the stochastic heterogeneity
in telomere-dependent senescence. PLoS Biol. 5:e1102007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tchkonia T, Morbeck DE, Von Zglinicki T,
et al: Fat tissue, aging, and cellular senescence. Aging Cell.
9:667–684. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fujikawa T, Shiraha H, Ueda N, et al:
Des-gamma-carboxyl prothrombin-promoted vascular endothelial cell
proliferation and migration. J Biol Chem. 282:8741–8748. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cho H, Balaji S, Sheikh AQ, et al:
Regulation of endothelial cell activation and angiogenesis by
injectable peptide nanofibers. Acta Biomater. 8:154–164. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Miyauchi H, Minamino T, Tateno K, Kunieda
T, Toko H and Komuro I: Akt negatively regulates the in vitro
lifespan of human endothelial cells via a p53/p21-dependent
pathway. EMBO J. 23:212–220. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim KS, Kang KW, Seu YB, Baek SH and Kim
JR: Interferon-gamma induces cellular senescence through
p53-dependent DNA damage signaling in human endothelial cells. Mech
Ageing Dev. 130:179–188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kida M, Sugiyama T, Yoshimoto T and Ogawa
Y: Hydrogen sulfide increases nitric oxide production with
calcium-dependent activation of endothelial nitric oxide synthase
in endothelial cells. Eur J Pharm Sci. 48:211–215. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Murikipudi S, Methe H and Edelman ER: The
effect of substrate modulus on the growth and function of
matrix-embedded endothelial cells. Biomaterials. 34:677–684. 2013.
View Article : Google Scholar : PubMed/NCBI
|