Introduction
Galectin-3, a member of the carbohydrate-binding
protein family, shows high affinity for β-galactoside-containing
glycoconjugates (1). Galectin-3 is
a unique chimera-type galectin containing three distinct structural
domains, including a short NH2-terminal domain of 12
amino acids, which regulates its cellular targeting, a C-terminal
carbohydrate recognition domain (CRD) with an amino-terminal tandem
repeat, the subcellular localization of which determines the wide
distribution and numerous biological functions of galectin-3, and a
repetitive collagen-like sequence rich in glycine, tyrosine and
proline that acts as a substrate for matrix metalloproteinases
(2–4). Galectin-3, dependent on its
sub-cellular location, has been shown to participate in multiple
biological functions, including cell growth, cell adhesion,
migration, metastasis, cell-cycle regulation, malignant
transformation, cell differentiation and inhibition of apoptosis
(5–8). For example, cell-surface galectin-3
is involved in cell adhesion, migration and cell-cell as well as
cell-matrix interaction (9).
Cytoplasmic galectin-3 mainly acts as an anti-apoptotic factor
(10,11), whereas nuclear galectin-3
participates in pre-mRNA splicing (12) and may regulate gene expression at
the transcriptional level (11,13).
Studies have demonstrated that galectin-3 is highly
expressed during neoplastic progression and metastasis in numerous
types of human malignant tumor. In melanoma, upregulation of
galectin-3 expression was implicated in tumor growth and metastasis
(14). Lotan et al
(15) observed that galectin-3
expression in certain primary and metastatic carcinomas was
elevated compared with that in adjacent normal mucosa. A study by
Inohara et al (16)
indicated that galectin-3 is consistently overexpressed in thyroid
carcinomas of follicular cell origin, whereas no expression of
galectin-3 is found in normal thyroid tissues, nodular goiters and
follicular adenomas. Galectin-3 expression in clear-cell renal cell
carcinoma with distant metastasis was significantly higher than in
those without distant metastasis (6).
Esophageal cancer (EC) is a type of invasive
malignant cancer with high mortality due to its early metastasis
and post-operative recurrence. In 2014, an estimated 18,170 people
were diagnosed with esophageal cancer and 15,450 people succumbed
to the disease in the United States (17). Based on the report by the
Esophageal Cancer Collaboration (WECC), patient survival decreased
with increased tumor invasion, as well as presence of regional
lymph node metastases and distant metastases (18). Numerous studies have been performed
to explore the mechanisms of tumor progression and metastasis in EC
(19–24); however, the exact mechanism
underlying the aggressiveness of this cancer type has largely
remained elusive. A previous study by our group (19) demonstrated that overexpression of
galectin-3 exerts important effects on the biological behavior of
Eca109 human EC cells, resulting in enhanced proliferation,
migration and invasion, as well as decreased apoptosis. Thus, the
present study examined the effects of galectin-3 knockdown using
small interfering RNA (siRNA) on multiple biological functions,
including proliferation, migration, invasion and apoptosis, in
Eca109 cells. The present study indicated that siRNA-mediated
knockdown of galectin-3 maybe represent a promising targeted
therapy approach for EC.
Materials and methods
Cell culture
The Eca109 human esophageal cancer cell line was
obtained from the Shandong Academy of Medical Sciences (Jinan,
China). All cells were cultured at 37°C in tissue culture flasks
(Corning-Costar, Corning, NY, USA) containing Dulbecco's modified
Eagle's medium (DMEM)-F12 (Gibco; Thermo Fisher Scientific,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco) and 1% penicillin-streptomycin (HyClone; GE Healthcare,
Little Chalfont, UK) in a humidified incubator containing 95% air
and 5% CO2.
siRNA transfection
The siRNAs (galectin-3 siRNA and non-silencing
siRNA) were designed and synthesized by Shanghai GenePharma Co.,
Ltd. (Shanghai, China). The siRNA-Gal-3 sequences were as follows:
Gal-3-homo-422 (siGal3-1), 5′-GCCACUGAUUGUGCCUUAUTT-3′;
Gal-3-homo-746 (siGal3-2), 5′-GUACAAUCAUCGGGUUAAATT-3′. As the
non-silencing siRNA (siRNA-control), a scrambled sequence
(5′-UUCUUCGAACGUGUCACGUTT-3′), which does not target any known
mammalian gene, was used as a negative control. The transfection
efficiency of siRNA was evaluated by using negative control
FAM-siRNA (Shanghai GenePharma Co., Ltd.), which emitted faint
green fluorescence. The lyophilized siRNAs were dissolved in
diethylpyrocarbonate-treated water according to the manufacturer's
instructions. Transfection of Eca109 cells with galectin-3 siRNA
was performed using HiperFect transfection reagent (Qiagen, Hilden,
Germany) according to the manufacturer's instructions. Eca109 cells
(10×104 cells in 2 ml complete culture medium) were
seeded into a six-well plate at 24 h prior to transfection. The
cells treated with siRNA were harvested within 48–72 h after
transfection for further experiments.
Western blot analysis
Four groups of Eca109 cells (untreated,
siRNA-control, siGal3-1 and siGal3-2) were harvested within 72 h
after transfection. Total protein was extracted using
radioimmunoprecipitation assay lysis buffer (Sigma-Aldrich, St.
Louis, MO, USA) according to the manufacturer's instructions. The
protein concentration was determined using a Bicinchoninic Acid
Protein Assay kit (Pierce Biotechnology, Inc., Rockford, IL, USA).
10% SDS-PAGE was used to isolate the total protein extracts (30
µg/well), which were transferred onto a polyvinylidene
membrane (Millipore, Billerica, MA, USA). The band representing
galectin-3 was determined using Protein Marker (Beyotime Institute
of Biotechnology, Haimen, China). The membrane was subsequently
blocked with 5% non-fat milk at 37°C for 1 h. Galectin-3 was
immunodetected with monoclonal anti-galectin-3 antibody (cat. no.
ab2785; 1:1,000; Abcam, Cambridge, UK) containing a ~30 kDa
recombinant fragment of human galectin-3 full-length protein,
overnight at 4°C. After washing with Tris-buffered saline
containing Tween-20 three times, the membranes were incubated with
sheep anti-rat immunoglobulin G conjugated to peroxidase (1:500;
Sigma-Aldrich). Based on the manufacturer's instructions,
immunosignals were visualized using the Protein Detector
5-bromo-4-chloro-3-indolyl-phosphate/nitro blue tetrazolium western
blotting kit (Beyotime Institute of Biotechnology). A FluorChem E
instrument (Cell Biosciences, Santa Clara, CA, USA) was utilized to
capture enhanced chemiluminescence images. Quantification of the
galectin-3 band intensity was performed using ImageJ software
(version 1.62; National Institutes of Health, Bethesda, MD, USA).
Another membrane that was incubated with anti-β-actin (1:1,000;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and green
fluorescence protein-conjugated (1:1,000; CoWin Biotech Co., Ltd.,
Beijing, China) antibodies was prepared in an identical manner.
Reverse-transcription quantitative
polymerase chain reaction (RT-qPCR)
Four groups of Eca109 cells (untreated,
siRNA-control, siGal3-1 and siGal3-2) were harvested 48 h
post-transfection. Total RNA was separated using RNA-solv reagent
(Omega Bio-Tek, Norcross, GA, USA) following the manufacturer's
instructions. After detection of the total RNA concentration by
spectrophotometry at a wavelength of 260 nm (SpectraMax 190;
Molecular Devices, Sunnyvale, CA, USA), total RNA incubated at 65°C
for 5 min was used to synthesize cDNA using a Rever Tra
Ace® qPCR-RT kit (Toyobo, Osaka, Japan) in a 10
µl reaction system containing 2 µl 5X RT buffer, 0.5
µl Enzyme Mix, 0.5 µl Primer Mix, 2 µl total
RNA and 5 µl nuclease-free water, with the following
reaction conditions: 5 min at 95°C and 15 min at 37°C. An ABI ViiA7
Dx instrument (Applied Biosystems; Thermo Fisher Scientific, Inc.)
was used to conduct PCR reactions with a 10 µl reaction
mixture containing 5 µl SYBR® Green (Toyobo), 2
µl nuclease-free water, 1 µl forward primer, 1
µl reverse primer and 1 µl cDNA. The thermocycling
conditions were as follows: 35 cycles of denaturation at 95°C for
15 sec, annealing at 61°C for 15 sec and extension at 72°C for 45
sec. The human galectin-3 mRNA sequences in GenBank (accession no.
NM_02306) were used to design the gene-specific primers (BioSune
Co., Ltd., Jinan, China) as follows: Forward,
5′-GGTGAAGCCCAATGCAAACA-3′ and reverse, 5′-TGCAACCTTGAAGTGGTCAG-3′.
Amplification of human β-actin mRNA was performed as a reference to
normalize sample loading using the following primers (Boshang Co.,
Ltd.): Forward, 5′-TGGCACCCAGCACAATGAA-3′ and reverse,
5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′. PCR results were quantified using
the ΔCt method according to the following formula: Expression ratio
= 2−ΔCt, where ΔCt = ΔCt target gene − ΔCt endogenous
control gene (β-actin) (25).
Cell proliferation assay
Eca-109 cells (3×103 cells/well)
transfected with either control or galectin-3 siRNA were dispensed
into 96-well round-bottomed microtiter plates in a total volume of
100 µl/well and incubated for 24–96 h at 37°C in 5%
CO2. Subsequently, 10 µl Cell Counting Kit-8
(CCK-8; Dojindo, Kumamoto, Japan) solution in 100 µl
complete DMEM was added to each well, followed by incubation for 4
h. The absorbance was measured at 450 nm using a spectrophotometer
(Spectramax 190; Molecular Devices).
Transwell migration and invasion
assay
Transwell polycarbonate filters (8.0-µm pore
size; Costar®; Sigma-Aldrich) in 24-well plate were used
to assess the migration and invasion of Eca109 cells. For the
migration assay, transfected or untransfected Eca109 cells at
4×104/200 µl in serum-free DMEM were seeded into
the upper chambers, while 600 µl DMEM containing 2.5% FBS
was added to the lower chamber. After incubation at 37°C in 5%
CO2 for 48 h, cells on the upper surface of the filter
were removed using a cotton swab. The cells migrated to the lower
side of the filter were fixed in methanol for 15 min at room
temperature, stained with 0.25% crystal violet (Beyotime Institute
of Biotechnology) for 10 min at room temperature and washed with
phosphate-buffered saline following staining. Images of the cells
migrated to the lower side of the filter were captured
(magnification, ×40) and counted (magnification, ×100) under an
inverted fluorescence microscope by an independent
investigator.
For the invasion assay, transfected or untransfected
Eca109 cells at 8×104/200 µl in serum-free DMEM
were seeded into the upper Transwell chambers, which were
pre-coated with 100 µl 1:5 diluted Matrigel (BD Biosciences,
Franklin Lakes, NJ, USA), while 600 µl DMEM containing 10%
FBS was added to the lower chambers. Following incubation at 37°C
in 5% CO2 for 48 h, the cells and Matrigel on the upper
surface of the membrane were removed using a cotton swab. The
number of invaded cells on the lower side of the membrane was
determined in an identical manner to that stated for the migration
assay.
Cell apoptosis assay
Apoptotic cells were analyzed by using an Annexin
V-Phycoerythrin (PE) Apoptosis Detection kit (BD Biosciences) and
flow cytometric analysis (FACSAria II; BD Biosciences).
Untransfected Eca109 cells or Eca109 cells transfected with siRNA
or control-siRNA for 48 h were re-suspended in 1X working solution
at 1×106 cells/ml. Following addition of 5 µl
Annexin V-PE and 5 µl 7-amino-actinomycin stain (7-AAD;
eBioscience, San Diego, CA, USA), cells were incubated at 25°C in
the dark for 15 min. Subsequently, 400 µl binding buffer was
added and cells were analyzed by flow cytometry within 1 h of
staining.
Statistical analysis
All statistical analyses were performed using SPSS
software (version 13.0; SPSS, Inc., Chicago, IL, USA). Values are
expressed as the mean ± standard deviation. Unpaired Student's
t-tests were utilized for comparisons between values. P<0.05 was
considered to indicate a statistically significant difference.
Results
Confirmation of galectin-3 silencing
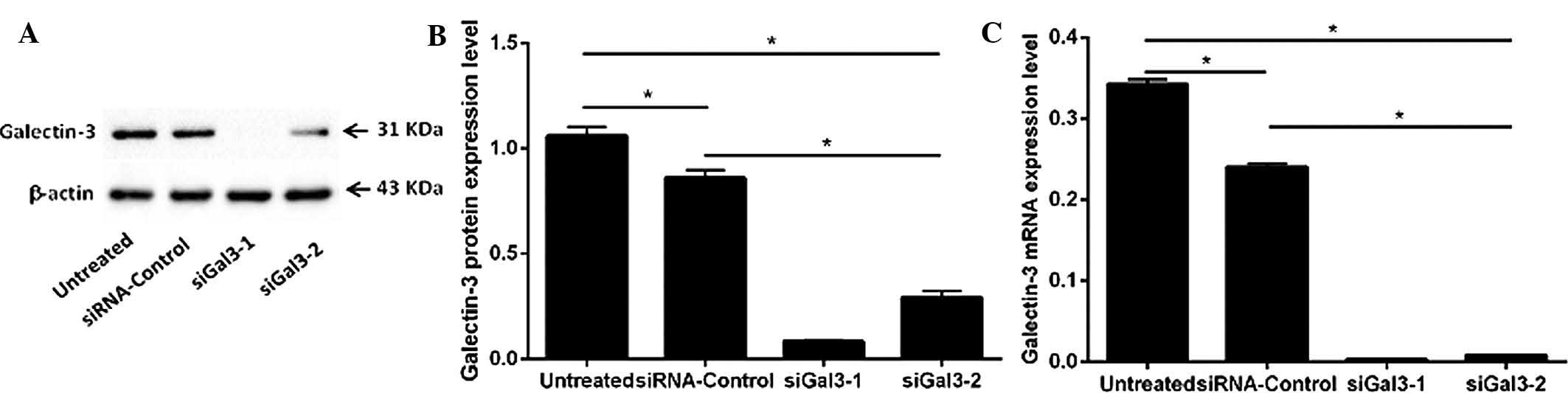
The protein expression of galectin-3 in the four
groups of Eca109 cells (siGal3-1, siGal3-2, siRNA-control and
untreated) was examined by western blot analysis (Fig. 1A and B). Variant expression was
detected in the 31-kDa band representing galectin-3. The expression
of galectin-3 in the two siGal3-transfected groups was
significantly decreased compared with that in the siRNA-control and
untreated groups (siGal3-1, P=0.001 and P=0.001; siGal3-2, P=0.002
and P=0.003, respectively). The galectin-3 suppression rates of
siGal3-1 and −2 reached 90.2 and 67.9%, respectively.
Furthermore, the mRNA levels of galectin-3 in the
four groups (siGal3-1, siGal3-2, siRNA-control and untreated) were
determined by RT-qPCR (Fig. 1C).
Compared with the siRNA-control cells or untreated cells,
galectin-3 mRNA was significantly reduced in the siGal3-1- and
siGal3-2-treated cells (P<0.05) with galectin-3 suppression
rates of 97.6 and 95.2%, respectively.
While a significant decrease in galectin-3
expression in the siRNA-control group compared with that in the
untreated group was observed at the protein and mRNA level
(P<0.05), this difference was minor compared to the knockdown
effect of siGal3-1 and −2.
The results suggested that Gal-3-homo-422 was more
effective at galectin-3 silencing than Gal-3-homo-746. Therefore,
Gal-3-homo-422 was used for gene knockdown in the subsequent
functional experiments.
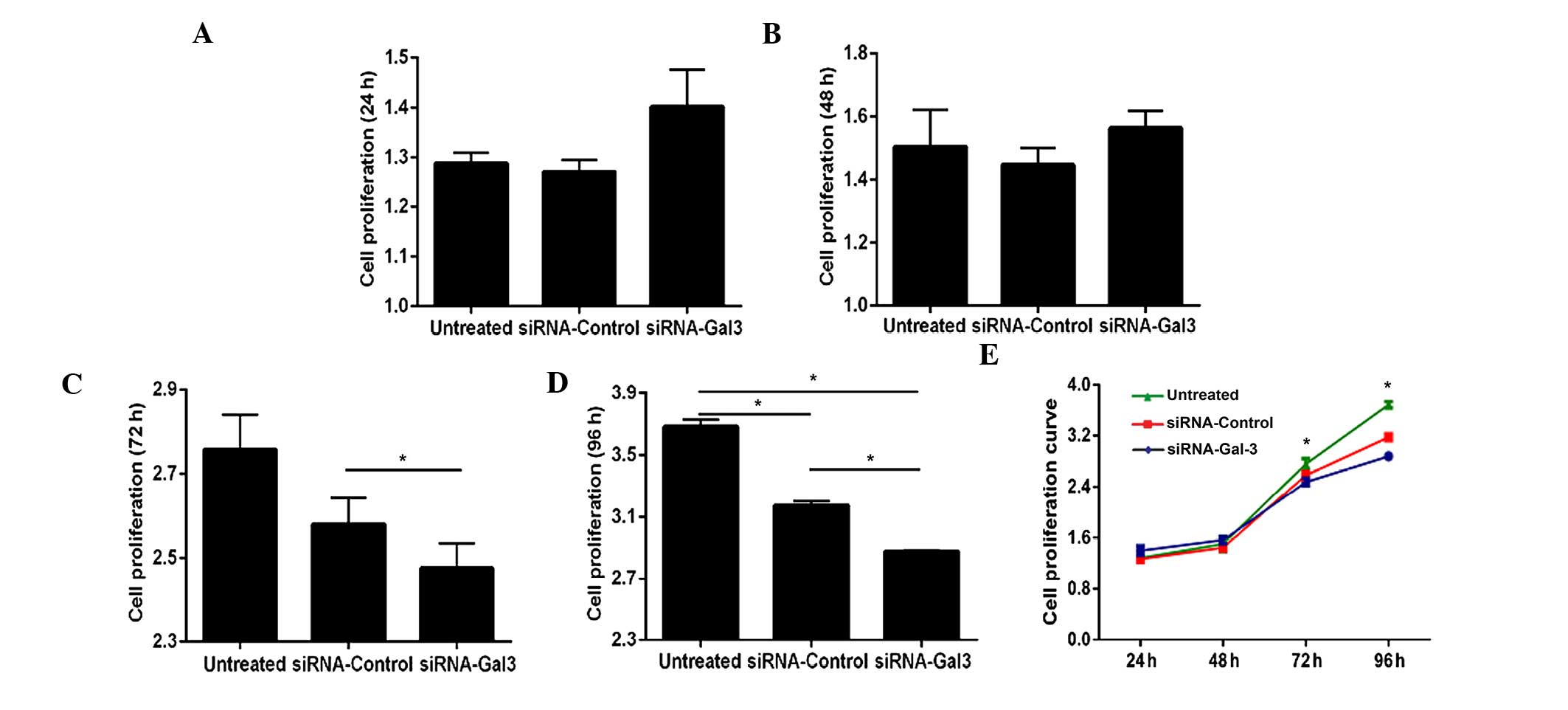
Galectin-3 silencing inhibits EC-cell
proliferation
The effects of galectin-3 silencing on the
proliferation of Eca109 cells were assessed using a CCK-8 assay
(Fig. 2). While transfection with
siRNA for 24 and 48 h did not significantly affect the
proliferation of Eca109 cells (P>0.05) (Fig. 2A, B and E), the proliferative rate
was significantly decreased following 72 h of transfection
(P<0.001), and it was further decreased at 96 h after
transfection (P=0.004 and P=0.001 vs. siRNA-control and untreated
groups, respectively) (Fig.
2C–E).
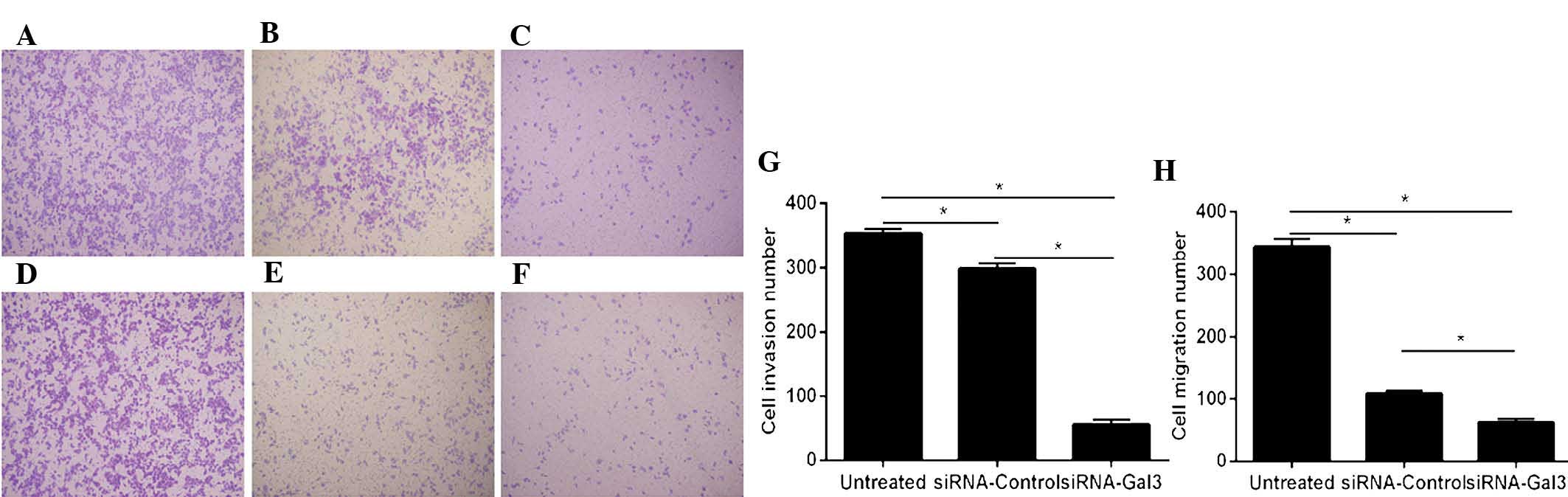
Galectin-3 silencing inhibits the
migration and invasion of EC cells
The migratory and invasive capacities of Eca109
cells were detected using Transwell assays (Fig. 3). A significantly decrease of cell
invasion was observed in the siRNA-Gal-3 group (55.3±7.37) compared
with that in the siRNA-control (298.3±8.33; P=0.001) and untreated
(354±7; P<0.001) groups.
Furthermore, the migration of Eca109 cells in the
siRNA-Gal-3 group (62.3±5.51) was obviously decreased compared with
that in the siRNA-control (109±4.59; P=0.002) and untreated
(344.3±13.01, P=0.001) groups.
In the invasion as well as in the migration assay,
significant differences were present between the siRNA-control and
untreated groups (P<0.05); however, these effects were minor
compared with those of siRNA-Gal-3.
Galectin-3 silencing enhances apoptosis
of EC cells
Flow cytometry was utilized to determine the
apoptotic rate of Eca109 cells (Fig.
4). The cells treated with siRNA-Gal-3 (8.8±3.08%) exhibited a
higher apoptotic rate than those in the siRNA-control (3.8±1.51%;
P=0.031) and untreated (3.33±0.72%; P=0.047) groups.
Discussion
Esophageal cancer is an aggressive digestive tract
malignancy and patients with increased tumor invasion or metastasis
generally exhibit reduced survival rates (18). At present, the process of
discovering effective treatment methods for EC is challenging due
to the biological behavior of EC cells, including enhanced
migration and invasion as well as evasion of apoptosis. Studies
have demonstrated that galectin-3 is highly expressed in various
tumor types and linked with tumor progression and metastasis
(6,14–16).
A previous study by our group revealed that overexpression of
galectin-3 enhanced the aggressive behavior of human esophageal
Eca109 cells, including increased proliferation, migration and
invasion, as well as reduced apoptosis (19). The present study successfully
performed siRNA-mediated silencing of galectin-3 in order to assess
its effects on multiple biological functions of the Eca109 human EC
cell line.
The migration and invasion of cancer cells has been
reported to be linked with adhesion and proteolysis of
extracellular matrix (ECM) components, which can be regulated by
integrins. Integrins are a family of 24 ubiquitous heterodimeric
transmembrane receptors that are expressed in multiple cell types
and are widely involved as key regulators of inflammation,
apoptosis and metastasis (26). In
numerous malignancies, overexpressed galectin-3 has been shown to
mediate cell migration and invasion by binding to integrins or
regulating their expression (27).
The present study performed knockdown of galectin-3 using
siRNA-Gal-3, which was shown to significantly reduce the migration
and invasion abilities of Eca109 cells compared with those of
untreated cells or those transfected with siRNA-control. This
result indicated a possible link between galectin-3 expression and
cell motility, which was consistent with studies on other malignant
tumor types. Kobayashi et al (28) showed that transient silencing of
galectin-3 reduced the migration and invasion of three human
pancreatic cancer cell lines. Honjo et al (29) revealed that in human breast
carcinoma cells, the suppression of galectin-3 expression blocked
tumorigenicity and led to a significant inhibition of tumor growth
in nude mice. Bresalier et al (30) discovered that a decrease of
galectin-3 expression was linked with a significant reduction of
liver colonization and spontaneous metastasis of human colon cancer
cells, whereas overexpression of galectin-3 led to an enhanced
metastastic potential. It has been reported that inhibition of
galectin-3 expression reduced the motility of human colon cancer
and glioblastoma cells (31,32).
Galectin-3 was shown to regulate metastasis of breast and prostate
cancer cells by binding to cell adhesion-associated molecules and
suppressing cell-cell and cell-ECM interactions in vitro as
well as in vivo (33).
Upregulation of galectin-3 expression in lung carcinoma cells was
reported to promote cell motility and invasion (34). However, a study by Junking et
al (35) indicated that
galectin-3 silencing enhanced cell motility; downregulation of
galectin-3 expression was associated with lymphatic invasion,
metastasis potential and poor differentiation of cholangiocarcinoma
(CCA). Of note, several studies indicated that galectin-3 was
overexpressed or downregulated in the same type of malignant tumor.
For example, in human colorectal carcinoma, upregulation of
galectin-3 expression was observed, which was linked with cancer
progression and metastasis (36,37),
and Zaia Povegliano et al (38) discovered that galectin-3 was
highest in the most advanced stage of colorectal cancer. By
contrast, downregulation of galectin-3 expression has also been
observed in colorectal carcinoma (39). These contradictory findings
indicated that differential galectin-3 expression leading to
increased or decreased cell invasion and metastasis may be
associated with the cancer type or stage and their associated
properties (35); however, the
exact mechanisms require further exploration. In the present study,
a significant difference was also observed between siRNA-control
and untreated groups in terms of galectin-3 expression as well as
cell migration and invasion, probably owing to the influences of
transfection reagent and negative control siRNA on Eca109
cells.
The present study also analyzed the effects of
galectin-3 silencing on Eca109-cell proliferation. At 72 and 96 h
after transfection, decreased cell proliferation in the siRNA-Gal-3
treated group was observed compared with that in the siRNA-control
and untreated groups, whereas at 24 and 48 h after transfection, no
significant difference in the proliferative rate was found among
the three groups. Galectin-3 was therefore shown to be associated
with cell proliferation in Eca109 cells, and inhibiting tumor cell
proliferation by downregulation of galectin-3 expression may
contribute to the interference with tumor progression.
Anti-apoptotic activity, the most distinct function
of galectin-3, has been shown to rely on its sub-cellular
localization with cytoplasmic galectin-3 acting as a main
anti-apoptotic factor. Galectin-3 has been demonstrated to exert
critical roles in the evasion of apoptosis by interacting with the
anti-apoptotic nuclear factor (NF)-κB signaling pathway and
inducing the activation of anti-apoptotic transcription factors
(40). In turn, NF-κB was shown to
be involved in the regulation of galectin-3 (41) and the expression of genes
associated with the immune response, inflammatory processes and
apoptosis (42,43). Furthermore, synexin was observed to
mediate the translocation of galectin-3 to perinuclear
mitochondrial membranes to suppress changes in the mitochondrial
membrane potential and to thereby inhibit apoptosis (10). In the present study, Annexin
V/7-AAD double-staining was utilized to assess the effects of
galectin-3 on the apoptotic rate of EC cells. Silencing of
galectin-3 was shown to promote apoptosis of Eca109 cells, which
was concordant with the results of previous studies and confirmed
the anti-apoptotic activity of galectin-3 in tumor cells. Shi et
al (37) revealed that
silencing of galectin-3 with siRNA contributed to the stimulation
of apoptosis in human colorectal cancer cells. Furthermore,
accumulating evidence implied that galectin-3-mediated resistance
to apoptosis is linked with resistance to chemotherapy. Lin et
al (44) demonstrated that
siRNA-mediated galectin-3 knockdown induced apoptosis, while
increased expression of galectin-3 enhanced chemoresistance to
doxorubicin with reduced doxo-rubicin-induced apoptosis and
expression of tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL). In bladder carcinoma cells, upregulation of
galectin-3 expression was also shown to inhibit TRAIL-induced
apoptosis (45). In addition, Choi
et al (46) reported that
breast carcinoma cells with galectin-3 overexpression exhibited
enhanced resistance to N-(4-hydroxyphenyl) retinamide-induced
apoptosis. Tepsiri et al (47) analyzed five human intrahepatic CCA
cell lines, revealing that the KKU-100 CCA cell line, in which
galectin-3 was most highly expressed, was resistant to all
chemotherapeutic drugs tested, whereas the KKU-M055 cell line,
which did not express galectin-3, was most sensitive to the
chemotherapeutic drugs. These studies indicated that targeting of
galectin-3 may represent a potential clinical application for the
treatment of malignant tumors. Although the molecular mechanisms by
which galectin-3 affects biological functions of cancer cells
remain to be fully elucidated, galectin-3 represents a promising
target for the treatment of EC, which warrants further
development.
In conclusion, the present study revealed that
siRNA-mediated galecin-3 silencing in the Eca109 EC cell line
resulted in the inhibition of cell proliferation, migration and
invasion, as well as an increase of cell apoptosis. While the exact
mechanism of the effects of galectin-3 on human EC cells and other
cancer types require further investigation to, galectin-3 silencing
was indicated to be a promising treatment strategy for EC.
Acknowledgments
The present study was supported by a grant from the
Natural Science Foundation of Shandong Province (no.
ZR2012HM095).
References
|
1
|
Dumic J, Dabelic S and Flögel M:
Galectin-3: An open-ended story. Biochim Biophys Acta.
1760:616–635. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barondes SH, Castronovo V, Cooper DN,
Cummings RD, Drickamer K, Feizi T, Gitt MA, Hirabayashi J, Hughes
C, Kasai K, et al: Galectins: A family of animal
β-galactoside-binding lectins. Cell. 76:597–598. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ochieng J, Fridman R, Nangia-Makker P,
Kleiner DE, Liotta LA, Stetler-Stevenson WG and Raz A: Galectin-3
is a novel substrate for human matrix metalloproteinases-2 and -9.
Biochemistry. 33:14109–14114. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gong HC, Honjo Y, Nangia-Makker P, Hogan
V, Mazurak N, Bresalier RS and Raz A: The NH2 terminus of
galectin-3 governs cellular compartmentalization and functions in
cancer cells. Cancer Res. 59:6239–6245. 1999.
|
|
5
|
Zhang HY, Jin L, Stilling GA, Ruebel KH,
Coonse K, Tanizaki Y, Raz A and Lloyd RV: RUNX1 and RUNX2
upregulate Galectin-3 expression in human pituitary tumors.
Endocrine. 35:101–111. 2009. View Article : Google Scholar
|
|
6
|
Sakaki M, Fukumori T, Fukawa T, Elsamman
E, Shiirevnyamba A, Nakatsuji H and Kanayama HO: Clinical
significance of Galectin-3 in clear cell renal cell carcinoma. J
Med Invest. 57:152–157. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jia W, Kidoya H, Yamakawa D, Naito H and
Takakura N: Galectin-3 accelerates M2 macrophage infiltration and
angio-genesis in tumors. Am J Pathol. 182:1821–1831. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu SW, Yu L, Zhou L, Cheng ZN and Tao YS:
Expression of Gal-3 and CD82/KAI1 proteins in non-small cell lung
cancer and their clinical significance. Chinese Journal of
Oncology. 35:124–128. 2013.In Chinese.
|
|
9
|
Ochieng J, Furtak V and Lukyanov P:
Extracellular functions of galectin-3. Glycoconj J. 19:527–535.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu F, Finley RL Jr, Raz A and Kim HR:
Galectin-3 translocates to the perinuclear membranes and inhibits
cytochrome c release from the mitochondria. A role for synexin in
galectin-3 translocation. J Biol Chem. 277:15819–15827. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takenaka Y, Fukumori T, Yoshii T, Oka N,
Inohara H, Kim HR, Bresalier RS and Raz A: Nuclear export of
phosphorylated galectin-3 regulates its antiapoptotic activity in
response to chemotherapeutic drugs. Mol Cell Biol. 24:4395–4406.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Patterson RJ, Wang W and Wang JL:
Understanding the biochemical activities of galectin-1 and
galectin-3 in the nucleus. Glycoconj J. 19:499–506. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shimura T, Takenaka Y, Tsutsumi S, Hogan
V, Kikuchi A and Raz A: Galectin-3, a novel binding partner of
beta-catenin. Cancer Res. 64:6363–6367. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Braeuer RR, Shoshan E, Kamiya T and
Bar-Eli M: The sweet and bitter sides of galectins in melanoma
progression. Pigment Cell Melanoma Res. 25:592–601. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lotan R, Ito H, Yasui W, Yokozaki H, Lotan
D and Tahara E: Expression of a 31-kDa lactose-binding lectin in
normal human gastric mucosa and in primary and metastatic gastric
carcinomas. Int J Cancer. 56:474–480. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Inohara H, Honjo Y, Yoshii T, Akahani S,
Yoshida J, Hattori K, Okamoto S, Sawada T, Raz A and Kubo T:
Expression of galectin-3 in fine-needle aspirates as a diagnostic
marker differentiating benign from malignant thyroid neoplasms.
Cancer. 85:2475–2484. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rice TW, Rusch VW, Apperson-Hansen C,
Allen MS, Chen LQ, Hunter JG, Kesler KA, Law S, Lerut TE, Reed CE,
et al: Worldwide esophageal cancer collaboration. Dis Esophagus.
22:1–8. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liang N, Song X, Xie J, Xu D, Liu F, Yu X,
Tian Y, Liu Z, Qiao L and Zhang J: Effect of galectin-3 on the
behavior of Eca-109 human esophageal cancer cells. Mol Med Rep.
11:896–902. 2015.
|
|
20
|
Du YY, Zhao LM, Chen L, Sang MX, Li J, Ma
M and Liu JF: The tumor-suppressive function of miR-1 by targeting
LASP1 and TAGLN2 in esophageal squamous cell carcinoma. J
Gastroenterol Hepatol. Epub ahead of print. 2015. View Article : Google Scholar
|
|
21
|
Li S, Qin X, Li Y, Zhang X, Niu R, Zhang
H, Cui A, An W and Wang X: MiR-133a suppresses the migration and
invasion of esophageal cancer cells by targeting the EMT regulator
SOX4. Am J Transl Res. 7:1390–1403. 2015.PubMed/NCBI
|
|
22
|
Zhou W, Yue H, Li C, Chen H and Yuan Y:
Protein arginine methyltransferase 1 promoted the growth and
migration of cancer cells in esophageal squamous cell carcinoma.
Tumour Biol. 2015.
|
|
23
|
Pei Y, Wang P, Liu H, He F and Ming L:
FOXQ1 promotes esophageal cancer proliferation and metastasis by
negatively modulating CDH1. Biomed Pharmacother. 74:89–94. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li J, Zhu SC, Li SG, Zhao Y, Xu JR and
Song CY: TKTL1 promotes cell proliferation and metastasis in
esophageal squamous cell carcinoma. Biomed Pharmacother. 74:71–76.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hood JD and Cheresh DA: Role of integrins
in cell invasion and migration. Nat Rev Cancer. 2:91–100. 2002.
View Article : Google Scholar
|
|
27
|
Liu FT and Rabinovich GA: Galectins as
modulators of tumour progression. Nat Rev Cancer. 5:29–41. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kobayashi T, Shimura T, Yajima T, Kubo N,
Araki K, Tsutsumi S, Suzuki H, Kuwano H and Raz A: Transient gene
silencing of galectin-3 suppresses pancreatic cancer cell migration
and invasion through degradation of β-catenin. Int J Cancer.
129:2775–2786. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Honjo Y, Nangia-Makker P, Inohara H and
Raz A: Down-regulation of Galectin-3 suppresses tumorigenicity of
human breast carcinoma cells. Clin Cancer Res. 7:661–668.
2001.PubMed/NCBI
|
|
30
|
Bresalier RS, Mazurek N, Sternberg LR,
Byrd JC, Yunker CK, Nangia-Makker P and Raz A: Metastasis of human
colon cancer is altered by modifying expression of the
beta-galactoside-binding protein galectin-3. Gastroenterology.
115:287–296. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hittelet A, Camby I, Nagy N, Legendre H,
Bronckart Y, Decaestecker C, Kaltner H, Nifant'ev NE, Bovin NV,
Pector JC, et al: Binding sites for lewis antigens are expressed by
human colon cancer cells and negatively affect their migration. Lab
Invest. 83:777–787. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Debray C, Vereecken P, Belot N, Teillard
P, Brion JP, Pandolfo M and Pochet R: Multifaceted role of
galectin-3 on human glioblastoma cell motility. Biochem Biophys Res
Commun. 325:1393–1398. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Glinsky VV, Glinsky GV, Glinskii OV,
Huxley VH, Turk JR, Mossine VV, Deutscher SL, Pienta KJ and Quinn
TP: Intravascular metastatic cancer cell homotypic aggregation at
the sites of primary attachment to the endothelium. Cancer Res.
63:3805–3811. 2003.PubMed/NCBI
|
|
34
|
O'Driscoll L, Linehan R, Liang YH, Joyce
H, Oglesby I and Clynes M: Galectin-3 expression alters adhesion,
motility and invasion in a lung cell line (DLKP), in vitro.
Anticancer Res. 22:3117–3125. 2002.
|
|
35
|
Junking M, Wongkham C, Sripa B,
Sawanyawisuth K, Araki N and Wongkham S: Decreased expression of
galectin-3 is associated with metastatic potential of liver
fluke-associated cholangiocarcinoma. Eur J Cancer. 44:619–626.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Endo K, Kohnoe S, Tsujita E, Watanabe A,
Nakashima H, Baba H and Maehara Y: Galectin-3 expression is a
potent prognostic marker in colorectal cancer. Anticancer Res.
25:3117–3121. 2005.PubMed/NCBI
|
|
37
|
Shi Y, He B, Kuchenbecker KM, You L, Xu Z,
Mikami I, Yagui-Beltran A, Clement G, Lin YC, Okamoto J, et al:
Inhibition of Wnt-2 and galectin-3 synergistically destabilizes
beta-catenin and induces apoptosis in human colorectal cancer
cells. Int J Cancer. 121:1175–1181. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zaia Povegliano L, Oshima CT, de Oliveira
Lima F, Andrade Scherholz PL and Manoukian Forones N:
Immunoexpression of galectin-3 in colorectal cancer and its
relationship with survival. J Gastrointest Cancer. 42:217–221.
2011. View Article : Google Scholar
|
|
39
|
Lotz MM, Andrews CW Jr, Korzelius CA, Lee
EC, Steele GD Jr, Clarke A and Mercurio AM: Decreased expression of
Mac-2 (carbohydrate binding protein 35) and loss of its nuclear
localization are associated with the neoplastic progression of
colon carcinoma. Proc Natl Acad Sci USA. 90:3466–3470. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu L, Sakai T, Sano N and Fukui K:
Nucling mediates apoptosis by inhibiting expression of galectin-3
through interference with nuclear factor kappaB signaling. Biochem
J. 380:31–41. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dumic J, Lauc G and Flögel M: Expression
of galectin-3 in cells exposed to stress-roles of jun and
NF-kappaB. Cell Physiol Biochem. 10:149–158. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Baichwal VR and Baeuerle PA: Activate
NF-kappaB or die? Curr Biol. 7:R94–R96. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Baeuerle PA and Henkel T: Function and
activation of NF-kappaB in the immune system. Annu Rev Immunol.
12:141–179. 1994. View Article : Google Scholar
|
|
44
|
Lin CI, Whang EE, Abramson MA, Donner DB,
Bertagnolli MM, Moore FD Jr and Ruan DT: Galectin-3 regulates
apoptosis and doxorubicin chemoresistance in papillary thyroid
cancer cells. Biochem Biophys Res Commun. 379:626–631. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Oka N, Nakahara S, Takenaka Y, Fukumori T,
Hogan V, Kanayama HO, Yanagawa T and Raz A: Galectin-3 inhibits
tumor necrosis factor-related apoptosis-inducing ligand-induced
apoptosis by activating Akt in human bladder carcinoma cells.
Cancer Res. 65:7546–7553. 2005.PubMed/NCBI
|
|
46
|
Choi JH, Chun KH, Raz A and Lotan R:
Inhibition of N-(4-hydroxyphenyl) retinamide-induced apoptosis in
breast cancer cells by galectin-3. Cancer Biol Ther. 3:447–452.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tepsiri N, Chaturat L, Sripa B, Namwat W,
Wongkham S, Bhudhisawasdi V and Tassaneeyakul W: Drug sensitivity
and drug resistance profiles of human intrahepatic
cholangiocar-cinoma cell lines. World J Gastroenterol.
11:2748–2753. 2005. View Article : Google Scholar : PubMed/NCBI
|