Introduction
Lung cancer is one of the most common malignancies
and has a significant socioeconomic impact on patients and their
families (1). In western
countries, the mortality rate of lung cancer is 15% and the
worldwide mortality rate for patients with lung cancer is 86%
(2). The high mortality of lung
cancer is mainly attributable to the lack of effective therapeutic
methods and the difficulty of obtaining an early diagnosis. Thus,
the development of effective therapeutic targets is urgently
required.
Differentially expressed genes (DEGs) have been
reported to have important roles in lung cancer, and their
identification may aid in the elucidation of its underlying
molecular mechanisms as well as the discovery of novel biomarkers
and treatments (3). Numerous
genes, including p53 (3,4),
EGFR (5,6), kRAS (7), PIK3CA (8) and EML4 (9), are known to be associated with lung
cancer, while others have remained elusive. Futhermore,
SEMA5A and -6A were identified as potential
therapeutic targets for lung cancer (10–12).
Although tremendous efforts have been made to discover novel
targets for lung cancer treatments, the current knowledge is
insufficient and requires expansion.
In the present study, DEGs between lung cancer and
normal lung tissues were identified. Protein-protein interaction
(PPI) and transcription factor (TF) regulatory networks were
constructed and key target genes were screened. Through the
identification of key genes, the possible underlying molecular
mechanisms as well as potential candidate biomarkers and treatment
targets for lung cancer were explored.
Materials and methods
Affymetrix microarray data
The gene expression profile dataset no. GSE3268
deposited in the Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo/) by Wachi et
al (13) based on the GPL96
platform (HG-U133A; Affymetrix Human Genome U133A Array), was
subjected to bioinformatics analysis in the present study. The
dataset contained a total of 10 chips, including five squamous cell
lung cancer tissues and five paired adjacent normal lung tissues
obtained from patients with squamous cell lung cancer.
Furthermore, the gene expression profile dataset
GSE19804 based on the platform GPL570 (HG-U133_Plus_2; Affymetrix
Human Genome U133 Plus 2.0 Array), which was deposited in the GEO
database by Lu et al (14),
was used. The dataset contained 120 chips, including 60 samples of
non-small cell lung cancer tissues and 60 samples of paired normal
lung tissues from female Taiwanese patients.
Identification of DEGs
The raw data were pre-processed using the Affy
package (15) in R language. DEGs
of GSE3268 (DEG1) and GSE19804 (DEG2) between normal groups and
disease groups were respectively analyzed using the limma package
in R (16). Fold changes (FCs) in
the expression of individual genes were calculated and DEGs with
P<0.05 and |log FC| >1 were considered to be significant.
DEG1 and DEG2 were then combined and the pooled dataset was
referred to as the overlapping DEGs in the present study.
Gene ontology (GO) and pathway enrichment
analysis of DEGs
GO analysis is a commonly used approach for
functional studies of large-scale transcriptomic data (17). The Kyoto Encyclopedia of Genes and
Genomes (KEGG) pathway database (18) contains information on networks of
molecules or genes. The Database for Annotation, Visualization and
Integrated Discovery (DAVID) (19)
was used to systematically extract biological information from the
large number of genes. GO functions and KEGG pathways of the
overlapping DEGs were analyzed using DAVID 6.7 with P<0.05.
Construction of PPI network and screening
of modules
The Search Tool for the Retrieval of Interacting
Genes (STRING) (20) database was
used to retrieve the predicted interactions for the DEGs; version
9.1 of STRING covers 1,133 completely sequenced species. All
associations obtained in STRING are provided with a confidence
score, which represents a rough estimate of the likelihood of a
given association to describe a functional linkage between two
proteins (21). The overlapping
DEGs with a confidence score >0.4 were selected to construct the
PPI network using Cytoscape software (version 3.0; http://cytoscape.org/) (22). Cytoscape allows for the
visualization of complex networks and their integration to any type
of attribute data. The MCODE (23)
plugin in Cytoscape was used to divide the PPI into modules. GO
functional analysis of genes in the modules was performed using the
BinGo 2.44 plugin in Cytoscape (24) with a threshold of P<0.05 using
the hypergeometric test.
Transcriptional regulatory network
construction
The University of California at Santa Cruz (UCSC)
database (http://genome.ucsc.edu) contains
information on TF binding sites and the regulated genes (25). Using information collected from the
UCSC database, DEGs were matched with their associated TFs. The TF
regulatory network then was constructed using Cytoscape software
(26).
Results
GO and pathway enrichment analysis of
DEGs
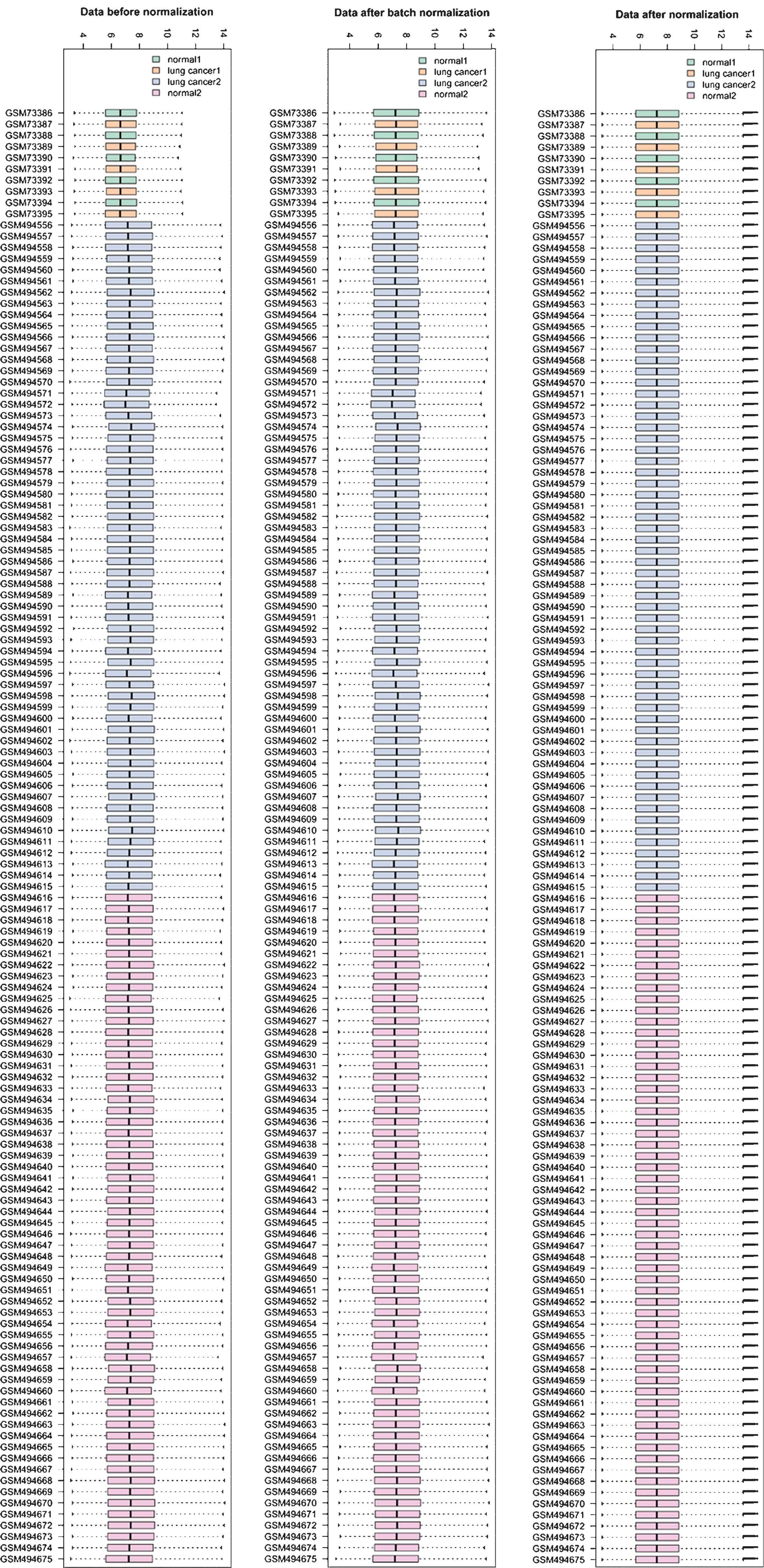
From the GEO datasets, information on the expression
of 8,172 genes was obtained. The normalized results showed that the
expression median after normalization was in a straight line
(Fig. 1). A total of 466 DEGs,
including 156 upregulated and 310 downregulated genes, were
selected.
Results of GO analysis showed that the upregulated
DEGs were significantly enriched in biological processes, including
collagen metabolic processes, multicellular organismal
macromolecule metabolic processes and nuclear division (Table I); the downregulated DEGs were
significantly enriched in biological processes, including response
to wounding, immune response, defense response and inflammatory
response (Table I).
 | Table IGO and pathway analysis of the
differentially expressed genes. |
Table I
GO and pathway analysis of the
differentially expressed genes.
| Expression | Category | Term/gene and
function | Count | P-value |
|---|
| Upregulated | KEGG_PATHWAY | hsa04110 - Cell
cycle | 12 |
6.94×10−7 |
| KEGG_PATHWAY | hsa04512 -
ECM-receptor interaction | 10 |
1.50×10−6 |
| KEGG_PATHWAY | hsa04510 - Focal
adhesion | 10 |
1.42×10−3 |
| KEGG_PATHWAY | hsa04115 - p53
signaling pathway | 6 |
2.14×10−3 |
| KEGG_PATHWAY | hsa00240 -
Pyrimidine metabolism | 5 |
3.93×10−2 |
| GOTERM_BP_FAT | GO:0032963 -
Collagen metabolic process | 9 |
2.10×10−10 |
| GOTERM_BP_FAT | GO:0044259 -
Multicellular organismal macromolecule metabolic process | 9 |
5.19×10−10 |
| GOTERM_BP_FAT | GO:0000280 -
Nuclear division | 17 |
5.79×10−10 |
| GOTERM_BP_FAT | GO:0007067 -
Mitosis | 17 |
5.79×10−10 |
| GOTERM_BP_FAT | GO:0000278 -
Mitotic cell cycle | 21 |
7.04×10−10 |
| GOTERM_BP_FAT | GO:0000087 - M
phase of mitotic cell cycle | 17 |
7.55×10−10 |
| GOTERM_CC_FAT | GO:0005576 -
Extracellular region | 53 |
1.41×10−10 |
| GOTERM_CC_FAT | GO:0005578 -
Proteinaceous extracellular matrix | 19 |
7.80×10−9 |
| GOTERM_CC_FAT | GO:0031012 -
Extracellular matrix | 19 |
2.50×10−8 |
| GOTERM_CC_FAT | GO:0044421 -
Extracellular region part | 30 |
2.27×10−7 |
| GOTERM_CC_FAT | GO:0005819 -
Spindle | 12 |
4.55×10−7 |
| GOTERM_MF_FAT | GO:0004222 -
Metalloendopeptidase activity | 9 |
9.37×10−6 |
| GOTERM_MF_FAT | GO:0048407 -
Platelet-derived growth factor binding | 4 |
1.53×10−4 |
| GOTERM_MF_FAT | GO:0004175 -
Endopeptidase activity | 13 |
3.80×10−4 |
| GOTERM_MF_FAT | GO:0004857 - Enzyme
inhibitor activity | 11 |
3.81×10−4 |
| Downregulated | KEGG_PATHWAY | hsa04060 -
Cytokine-cytokine receptor interaction | 20 |
6.99×10−5 |
| KEGG_PATHWAY | hsa04610 -
Complement and coagulation cascades | 8 |
2.47×10−3 |
| KEGG_PATHWAY | hsa04062 -
Chemokine signaling pathway | 13 |
4.53×10−3 |
| KEGG_PATHWAY | hsa04650 - Natural
killer cell mediated cytotoxicity | 10 |
9.69×10−3 |
| KEGG_PATHWAY | hsa04614 -
Renin-angiotensin system | 4 |
1.01×10−2 |
| GOTERM_BP_FAT | GO:0009611 -
Response to wounding | 48 |
2.23×10−17 |
| GOTERM_BP_FAT | GO:0006952 -
Defense response | 46 |
1.66×10−13 |
| GOTERM_BP_FAT | GO:0006954 -
Inflammatory response | 33 |
2.92×10−13 |
| GOTERM_BP_FAT | GO:0006955 - Immune
response | 43 |
4.20×10−10 |
| GOTERM_BP_FAT | GO:0048545 -
Response to steroid hormone stimulus | 21 |
3.81×10−9 |
| GOTERM_CC_FAT | GO:0005615 -
Extracellular space | 55 |
2.36×10−18 |
| GOTERM_CC_FAT | GO:0044421 -
Extracellular region part | 64 |
2.03×10−17 |
| GOTERM_CC_FAT | GO:0005576 -
Extracellular region | 93 |
3.37×10−15 |
| GOTERM_CC_FAT | GO:0005886 - Plasma
membrane | 131 |
2.25×10−12 |
| GOTERM_CC_FAT | GO:0005887 -
Integral to plasma membrane | 61 |
1.99×10−11 |
| GOTERM_MF_FAT | GO:0019838 - Growth
factor binding | 16 |
2.01×10−9 |
| GOTERM_MF_FAT | GO:0030246 -
Carbohydrate binding | 27 |
7.86×10−9 |
| GOTERM_MF_FAT | GO:0019955 -
Cytokine binding | 13 |
1.54×10−6 |
| GOTERM_MF_FAT | GO:0005509 -
Calcium ion binding | 39 |
1.04×10−5 |
| GOTERM_MF_FAT | GO:0030247 -
Polysaccharide binding | 14 |
1.11×10−5 |
Pathway analysis showed that the upregulated DEGs
were significantly enriched in cell cycle, extracellular matrix -
receptor interaction and the p53 signaling pathway (Table I); the downregulated DEGs were
significantly enriched in cytokine receptor interaction, complement
and coagulation cascades as well as chemokine signaling pathways
(Table I).
Construction of PPI network and screening
of module
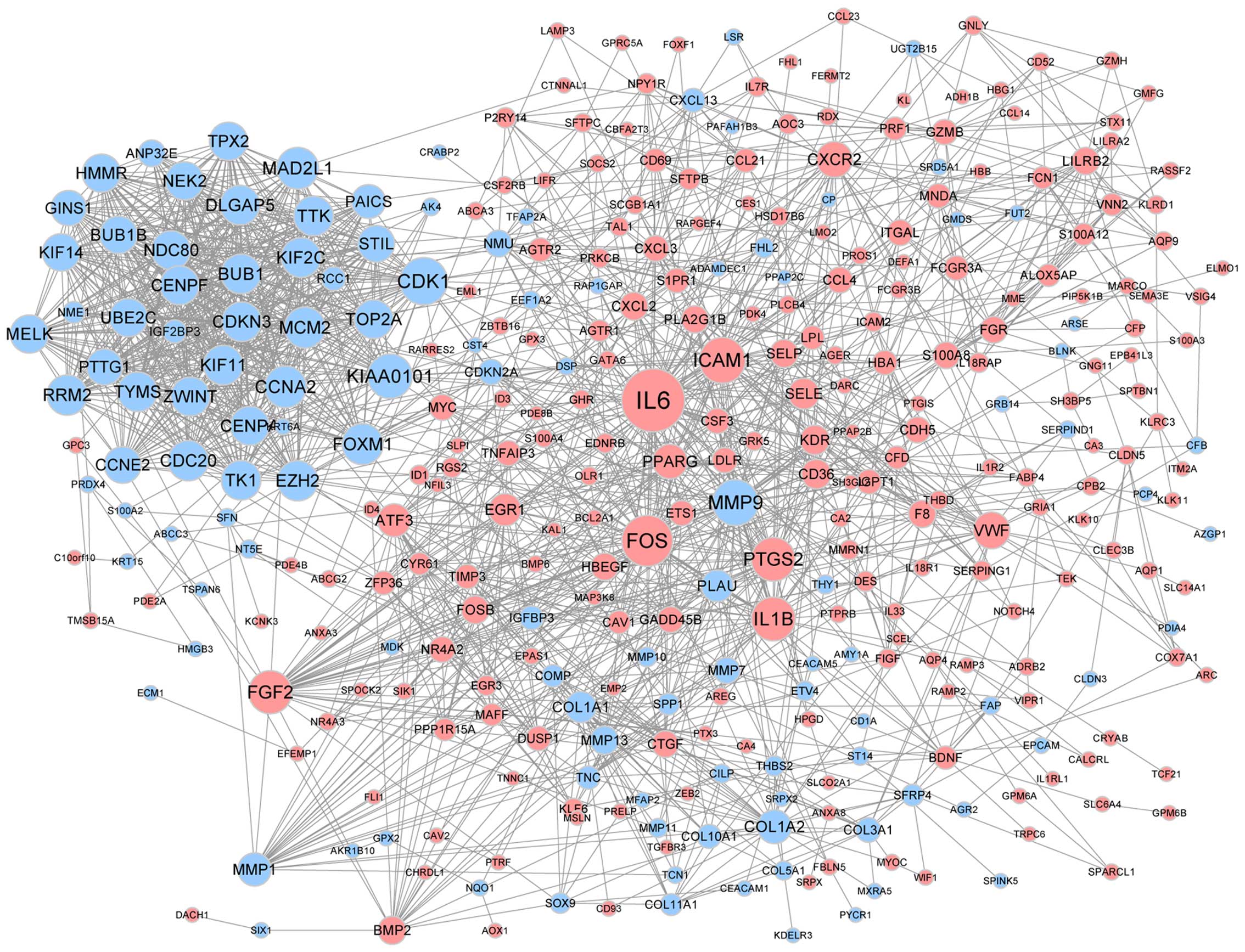
The PPI network was constructed based on the
predicted interactions of the identified DEGs (Fig. 2). Genes of IL-6,
FOSB, CDK1, MMP9 and ICAM1 were found
to have a high degree of interaction in lung cancer. A sub-network
containing 34 nodes and 547 edges was screened from the PPI
network, such as PTTG1 (Fig.
3). The DEGs in the sub-net were significantly enriched in
biological processes, such as the cell cycle, and pathway analysis
showed that they were significantly enriched in cell cycle and
oocyte meiosis (Table II).
| Table IIGO and pathway analysis of genes in
sub-network. |
Table II
GO and pathway analysis of genes in
sub-network.
| Category | Term/gene and
function | Count | P-value |
|---|
| KEGG_PATHWAY | hsa04110 - Cell
cycle | 10 |
1.09×10−11 |
| KEGG_PATHWAY | hsa04114- Oocyte
meiosis | 6 |
1.09×10−5 |
| KEGG_PATHWAY | hsa04914 -
Progesterone-mediated oocyte maturation | 4 |
1.83×10−3 |
| KEGG_PATHWAY | hsa04115 - p53
signaling pathway | 3 |
1.65×10−3 |
| KEGG_PATHWAY | hsa00240 -
Pyrimidine metabolism | 3 |
3.10×10−2 |
| GOTERM_BP_FAT | GO:0000278 -
Mitotic cell cycle | 19 |
7.13×10−21 |
| GOTERM_BP_FAT | GO:0007049 - Cell
cycle | 22 |
1.65×10−19 |
| GOTERM_BP_FAT | GO:0000280 -
Nuclear division | 16 |
2.14×10−19 |
| GOTERM_BP_FAT | GO:0007067 -
Mitosis | 16 |
2.14×10−19 |
| GOTERM_BP_FAT | GO:0000087 - M
phase of mitotic cell cycle | 16 |
2.82×10−19 |
| GOTERM_CC_FAT | GO:0005819 -
Spindle | 12 |
9.20×10−15 |
| GOTERM_CC_FAT | GO:0000777 -
Condensed chromosome kinetochore | 8 |
3.94×10−11 |
| GOTERM_CC_FAT | GO:0015630 -
Microtubule cytoskeleton | 14 |
5.31×10−11 |
| GOTERM_CC_FAT | GO:0000779 -
Condensed chromosome, centromeric region | 8 |
1.01×10−10 |
| GOTERM_CC_FAT | GO:0000922 -
Spindle pole | 7 |
1.01×10−10 |
| GOTERM_MF_FAT | GO:0005524 -
Adenosine triphosphate binding | 15 |
4.89×10−7 |
| GOTERM_MF_FAT | GO:0032559 - Adenyl
ribonucleotide binding | 15 |
5.78×10−7 |
| GOTERM_MF_FAT | GO:0030554 - Adenyl
nucleotide binding | 15 |
1.10×10−6 |
| GOTERM_MF_FAT | GO:0001883 - Purine
nucleoside binding | 15 |
1.32×10−6 |
| GOTERM_MF_FAT | GO:0001882 -
Nucleoside binding | 15 |
1.44×10−6 |
TF-target gene regulatory network
analysis
Associations between 44 TFs and their 47 target DEGs
were collected from the TF regulatory network (Fig. 4). TFs of FOSB and
LMO2, which exhibited a high degree of interaction, were
selected from this network. Furthermore, the results also showed
that MMP9 was the target of FOSB and PTTG1 was
the target of LMO2.
Discussion
Lung cancer is the leading cause of
cancer-associated mortality; however, the underlying molecular
mechanisms of its development and progression have remained to be
fully elucidated (1). The present
study used a bioinformatics approach to predict the potential
therapeutic targets and explore the possible molecular mechanisms
for lung cancer. A total of 466 DEGs between tumorous and normal
tissues was identified, among which 310 genes were downregulated
and 156 were upregulated. By constructing a PPI network and a TF
regulatory network, key genes, including IL6, MMP9
and PTTG1, were identified.
IL-6 is a multifunctional cytokine that was
characterized as a regulator of immune and inflammatory responses
(27,28). It is involved in the regulation of
cell proliferation, survival and metabolism, and IL-6 signaling has
an important role in tumorigenesis (29). Chung et al (30) found that IL-6 activated PI3K, which
promoted apoptosis in human prostate cancer cell lines.
Furthermore, studies have shown that IL-6 inhibited the growth of
numerous types of cancer, including lung (31), breast (32) and prostate cancer (33). In the present study, IL-6
was shown to be downregulated in squamous cell and non-small cell
lung cancer, and GO analysis showed that IL-6 was
significantly enriched in biological processes, including defense
response, inflammatory response, immune response and regulation of
cell proliferation, which was consistent with a previous study
(29). Combined with the above
studies, it is indicated that IL-6 may be a diagnostic
biomarker and therapeutic target in lung cancer.
MMP9 has a key role in cell migration,
proliferation, differentiation, angiogenesis, apoptosis and host
defense (34). Dysregulatoin of
MMPs has been implicated in numerous diseases, including chronic
ulcers and cancer (35–37). Downregulation of MMPs has been
shown to inhibit metastasis, while upregulation of MMPs led to
enhanced cancer cell invasion (37). In the present study, MMP9
was overexpressed and regulated by FOSB in lung cancer
tissues. Kim et al (38)
found that FOSB was downregulated in pancreatic cancer and
promoted tumor progression. Kataoka et al (39) found that FOSB gene
expression in cancer stroma is a independent prognostic indicator
for patients with epithelial ovarian cancer receiving standard
therapy. Combined with the above studies, the present study
indicated that MMP9 may have important roles in the
progression of lung cancer, and that it may be utilized as a
therapeutic target.
PTTG1 has tumorigenic activity and is highly
expressed in various tumor types (40). Studies have shown that PTTG1
was overexpressed in esophageal cancer and associated with
endocrine therapy resistance in breast cancer (41,42).
Yoon et al (40) showed
that the PTTG1 oncogene promoted tumor malignancy via
epithelial-to-mesenchymal expansion of the cancer stem cell
population. Hamid et al (43) found that PTTG1 promoted
tumorigenesis in human embryonic kidney cells. A study by Li et
al (44) indicated that
PTTG1 promoted migration and invasion of human non-small
cell lung cancer cells. Panguluri et al (45) showed that PTTG1 was an
important target gene for ovarian cancer therapy. In the present
study, PTTG1 was found to be overexpressed in lung cancer
tissues and regulated by LMO2. LMO2 is an important regulator in
determining cell fate and controlling cell growth and
differentiation (46). Nakata
et al (47) found that
LMO2 was a novel predictive biomarker with the potential to
enhance the accuracy of prognoses for pancreatic cancer. Yamada
et al (48) showed that
LMO2 is a key regulator of tumour angiogenesis. Combined with the
above studies, the present study indicated that PTTG1 may
have important roles in the progression of lung cancer and that it
may represent a therapeutic target.
In conclusion, the bioinformatics analysis of the
present study indicated that IL-6, MMP9 and
PTTG1 may have key roles in the progression and development
of lung cancer. They may be used as prognostic biomarkers as well
as specific therapeutic targets for the treatment of lung cancer.
However, molecular biology experiments are required to confirm
these findings.
References
|
1
|
Nugent M, Edney B, Hammerness PG, Dain BJ,
Maurer LH and Rigas JR: Non-small cell lung cancer at the extremes
of age: Impact on diagnosis and treatment. Ann Thorac Surg.
63:193–197. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang SP, Luh KT, Kuo SH and Lin CC:
Chronological observation of epidemiological characteristics of
lung cancer in Taiwan with etiological consideration-a 30-year
consecutive study. Jpn J Clin Oncol. 14:7–19. 1984.PubMed/NCBI
|
|
3
|
Andriani F, Roz E, Caserini R, Conte D,
Pastorino U, Sozzi G and Roz L: Inactivation of both FHIT and p53
cooperate in deregulating proliferation-related pathways in lung
cancer. J Thorac Oncol. 7:631–642. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Toyooka S, Tsuda T and Gazdar AF: The TP53
gene, tobacco exposure and lung cancer. Hum Mutat. 21:229–239.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shigematsu H, Lin L, Takahashi T, Nomura
M, Suzuki M, Wistuba II, Fong KM, Lee H, Toyooka S, Shimizu N, et
al: Clinical and biological features associated with epidermal
growth factor receptor gene mutations in lung cancers. J Natl
Cancer Inst. 97:339–346. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martin P, Kelly CM and Carney D: Epidermal
growth factor receptor-targeted agents for lung cancer. Cancer
Control. 13:129–140. 2006.PubMed/NCBI
|
|
7
|
Eberhard DA, Johnson BE, Amler LC, Goddard
AD, Heldens SL, Herbst RS, Ince WL, Jänne PA, Januario T, Johnson
DH, et al: Mutations in the epidermal growth factor receptor and in
KRAS are predictive and prognostic indicators in patients with
non-small-cell lung cancer treated with chemotherapy alone and in
combination with erlotinib. J Clin Oncol. 23:5900–5909. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yamamoto H, Shigematsu H, Nomura M,
Lockwood WW, Sato M, Okumura N, Soh J, Suzuki M, Wistuba II, Fong
KM, et al: PIK3CA mutations and copy number gains in human lung
cancers. Cancer Res. 68:6913–6921. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wong DW, Leung EL, So KK, Tam IY, Sihoe
AD, Cheng LC, Ho KK, Au JS, Chung LP, Pik Wong M, et al: The
EML4-ALK fusion gene is involved in various histologic types of
lung cancers from nonsmokers with wild-type EGFR and KRAS. Cancer.
115:1723–1733. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Castro-Rivera E, Ran S, Thorpe P and Minna
JD: Semaphorin 3B (SEMA3B) induces apoptosis in lung and breast
cancer, whereas VEGF165 antagonizes this effect. Proc Natl Acad Sci
USA. 101:11432–11437. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tomizawa Y, Sekido Y, Kondo M, Gao B,
Yokota J, Roche J, Drabkin H, Lerman MI, Gazdar AF, Minna JD, et
al: Inhibition of lung cancer cell growth and induction of
apoptosis after reex-pression of 3p21. 3 candidate tumor suppressor
gene SEMA3B. Proc Natl Acad Sci USA. 98:13954–13959. 2001.
View Article : Google Scholar
|
|
12
|
Brambilla E, Constantin B, Drabkin H and
Roche J: Semaphorin SEMA3F localization in malignant human lung and
cell lines: A suggested role in cell adhesion and cell migration.
Am J Pathol. 156:939–950. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wachi S, Yoneda K and Wu R:
Interactome-transcriptome analysis reveals the high centrality of
genes differentially expressed in lung cancer tissues.
Bioinformatics. 21:4205–4208. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu TP, Tsai MH, Lee JM, Hsu CP, Chen PC,
Lin CW, Shih JY, Yang PC, Hsiao CK, Lai LC, et al: Identification
of a novel biomarker, SEMA5A, for non-small cell lung carcinoma in
nonsmoking women. Cancer Epidemiol Biomarkers Prev. 19:2590–2597.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: Affy-analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Smyth GK: Linear models and empirical
bayes methods for assessing differential expression in microarray
experiments. Stat Appl Genet Mol Biol. 3:2004.
|
|
17
|
Hulsegge I, Kommadath A and Smits MA:
Globaltest and GOEAST: Two different approaches for Gene Ontology
analysis. BMC Proc. 3(Suppl 4): S102009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ogata H, Goto S, Sato K, Fujibuchi W, Bono
H and Kanehisa M: KEGG: Kyoto encyclopedia of genes and genomes.
Nucleic Acids Res. 27:29–34. 1999. View Article : Google Scholar
|
|
19
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization and integrated discovery. Genome Biol. 4:P32003.
View Article : Google Scholar
|
|
20
|
Franceschini A, Szklarczyk D, Frankild S,
Kuhn M, Simonovic M, Roth A, Lin J, Minguez P, Bork P, von Mering
C, et al: STRING v9.1: Protein-protein interaction networks, with
increased coverage and integration. Nucleic Acids Res.
41:D808–D815. 2013. View Article : Google Scholar :
|
|
21
|
Szklarczyk D, Franceschini A, Kuhn M,
Simonovic M, Roth A, Minguez P, Doerks T, Stark M, Muller J, Bork
P, et al: The STRING database in 2011: Functional interaction
networks of proteins, globally integrated and scored. Nucleic Acids
Res. 39:D561–D568. 2011. View Article : Google Scholar :
|
|
22
|
Kohl M, Wiese S and Warscheid B:
Cytoscape: Software for visualization and analysis of biological
networks. Methods Mol Biol. 696:291–303. 2011. View Article : Google Scholar
|
|
23
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maere S, Heymans K and Kuiper M: BiNGO: A
cytoscape plugin to assess overrepresentation of gene ontology
categories in biological networks. Bioinformatics. 21:3448–3449.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wingender E, Dietze P, Karas H and Knüppel
R: TRANSFAC: A database on transcription factors and their DNA
binding sites. Nucleic Acids Res. 24:238–241. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schafer ZT and Brugge JS: IL-6 involvement
in epithelial cancers. J Clin Invest. 117:3660–3663. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kishimoto T: Interleukin-6: From basic
science to medicine-40 years in immunology. Annu Rev Immunol.
23:1–21. 2005. View Article : Google Scholar
|
|
29
|
Hodge DR, Hurt EM and Farrar WL: The role
of IL-6 and STAT3 in inflammation and cancer. Eur J Cancer.
41:2502–2512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chung TD, Yu JJ, Kong TA, Spiotto MT and
Lin JM: Interleukin-6 activates phosphatidylinositol-3 kinase,
which inhibits apoptosis in human prostate cancer cell lines.
Prostate. 42:1–7. 2000. View Article : Google Scholar
|
|
31
|
Takizawa H, Ohtoshi T, Ohta K, Yamashita
N, Hirohata S, Hirai K, Hiramatsu K and Ito K: Growth inhibition of
human lung cancer cell lines by interleukin 6 in vitro: A possible
role in tumor growth via an autocrine mechanism. Cancer Res.
53:4175–4181. 1993.PubMed/NCBI
|
|
32
|
Knüpfer H and Preiss R: Significance of
interleukin-6 (IL-6) in breast cancer (review). Breast Cancer Res
Treat. 102:129–135. 2007. View Article : Google Scholar
|
|
33
|
Giri D, Ozen M and Ittmann M:
Interleukin-6 is an autocrine growth factor in human prostate
cancer. Am J Pathol. 159:2159–2165. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sica A, Allavena P and Mantovani A: Cancer
related inflammation: The macrophage connection. Cancer Lett.
267:204–215. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Benveniste EN: Role of
macrophages/microglia in multiple sclerosis and experimental
allergic encephalomyelitis. J Mol Med (Berl). 75:165–173. 1997.
View Article : Google Scholar
|
|
36
|
Firestein GS: Evolving concepts of
rheumatoid arthritis. Nature. 423:356–361. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Coussens LM, Fingleton B and Matrisian LM:
Matrix metal-loproteinase inhibitors and cancer: Trials and
tribulations. Science. 295:2387–2392. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim JH, Lee JY, Lee KT, Lee JK, Lee KH,
Jang KT, Heo JS, Choi SH and Rhee JC: RGS16 and FosB underexpressed
in pancreatic cancer with lymph node metastasis promote tumor
progression. Tumor Biol. 31:541–548. 2010. View Article : Google Scholar
|
|
39
|
Kataoka F, Tsuda H, Arao T, Nishimura S,
Tanaka H, Nomura H, Chiyoda T, Hirasawa A, Akahane T, Nishio H, et
al: EGRI and FOSB gene expressions in cancer stroma are independent
prognostic indicators for epithelial ovarian cancer receiving
standard therapy. Gene Chromosome Cancer. 51:300–312. 2012.
View Article : Google Scholar
|
|
40
|
Yoon CH, Kim MJ, Lee H, Kim RK, Lim EJ,
Yoo KC, Lee GH, Cui YH, Oh YS, Gye MC, et al: PTTG1 oncogene
promotes tumor malignancy via epithelial to mesenchymal transition
and expansion of cancer stem cell population. J Biol Chem.
287:19516–19527. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shibata Y, Haruki N, Kuwabara Y, Nishiwaki
T, Kato J, Shinoda N, Sato A, Kimura M, Koyama H, Toyama T, et al:
Expression of PTTG (pituitary tumor transforming gene) in
esophageal cancer. Jpn J Clin Oncol. 32:233–237. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ghayad SE, Vendrell JA, Bieche I, Spyratos
F, Dumontet C, Treilleux I, Lidereau R and Cohen PA: Identification
of TACC1, NOV and PTTG1 as new candidate genes associated with
endocrine therapy resistance in breast cancer. J Mol Endocrinol.
42:87–103. 2009. View Article : Google Scholar
|
|
43
|
Hamid T, Malik MT and Kakar SS: Ectopic
expression of PTTG1/securin promotes tumorigenesis in human
embryonic kidney cells. Mol Cancer. 4:32005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li H, Yin C, Zhang B, Sun Y, Shi L, Liu N,
Liang S, Lu S, Liu Y, Zhang J, et al: PTTG1 promotes migration and
invasion of human non-small cell lung cancer cells and is modulated
by miR-186. Carcinogenesis. 34:2145–2155. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Panguluri SK, Yeakel C and Kakar SS: PTTG:
An important target gene for ovarian cancer therapy. J Ovarian Res.
1:62008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ma S, Guan XY, Beh PS, Wong KY, Chan YP,
Yuen HF, Vielkind J and Chan KW: The significance of LMO2
expression in the progression of prostate cancer. J Pathol.
211:278–285. 2007. View Article : Google Scholar
|
|
47
|
Nakata K, Ohuchida K, Nagai E, Hayashi A,
Miyasaka Y, Kayashima T, Yu J, Aishima S, Oda Y, Mizumoto K, et al:
LMO2 is a novel predictive marker for a better prognosis in
pancreatic cancer. Neoplasia. 11:712–719. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yamada Y, Pannell R, Forster A and
Rabbitts TH: The LIM-domain protein Lmo2 is a key regulator of
tumour angiogenesis: A new anti-angiogenesis drug target. Oncogene.
21:1309–1315. 2002. View Article : Google Scholar : PubMed/NCBI
|