Introduction
Green tea and coffee are traditionally consumed as
dietary beverages all over the world. It is generally recognized
that polyphenolic compounds in green tea have potentially
beneficial effects on human health including anti-inflammation,
anti-oxidation and prevention of tumor development (1,2). In
addition, polyphenolic compounds in coffee are proposed to
potentiate human health benefits such as prevention of type 2
diabetes mellitus and Parkinson's disease (3). A major flavonoid of green tea is
(−)-epigallocatechin gallate (EGCG), which is considered as one of
the most active polyphenolic molecules relating to the health
promoting properties of this beverage (4). On the other hand, chlorogenic acid
(CGA) is a main phenolic compound in coffee (5). Regarding the beneficial properties of
green tea and coffee for bone disorders, accumulating evidence
indicates that consumption of green tea prevents both age-related
bone loss and fracture in elderly people (6), and that coffee consumption reduces
the risk of osteoporosis and osteoporotic fracture (3). However, the details underlying the
beneficial effects of green tea and coffee on bone remain
unclear.
Bone metabolism is mainly regulated by two types of
functional cells, osteoblasts and osteoclasts (7). The former cells are responsible for
bone formation, and the latter cells are for bone resorption. In
the adult skeletal system, bone mass is continuously maintained by
resorption of old bone and subsequent formation of new bone, so
called bone remodeling. Disorder of this important process is
considered to cause metabolic bone diseases, including osteoporosis
and fracture healing distress. It is currently recognized that
various humoral factors including cytokines, hormones, growth
factors and prostaglandins (PGs) play important roles as bone
remodeling mediators (8). As for
the EGCG-effects on bone metabolism, it has been shown that EGCG
attenuates bone resorption through the inhibition of osteoclast
formation, resulting in suppression of osteoclastogenesis (5). In osteoblasts, EGCG reportedly
increases alkaline phosphatase activity followed by increment of
osteoblastic bone formation (5).
On the other hand, it has been reported that CGA suppresses
osteoclast-mediated bone resorption by downregulation of receptor
activator of nuclear factor-κB (RANK) ligand-mediating effects
(9). In vivo studies in rat
model, CGA increases mineralization in the tibia and improve
mechanical properties of the femoral diaphysis (10). We have previously reported that
EGCG suppresses the interleukin-6 (IL-6) synthesis stimulated by
platelet-derived growth factor-BB, basic fibroblast growth factor
or endothelin-1 in osteoblast-like MC3T3-E1 cells (11–13).
Additionally, we recently demonstrated that EGCG and CGA enhance
the tumor necrosis factor-α (TNF-α)-stimulated IL-6 synthesis in
these cells (14). However, the
exact mechanism behind the effects of EGCG and CGA on bone
remodeling has not yet been elucidated.
Osteoprotegerin (OPG) is a cytokine which has an
inhibitory effect on osteoclast functions. OPG is classified into a
member of the TNF receptor family along with RANK (15). OPG, which is produced in
osteoblasts and secreted, binds to RANK ligand (RANKL) as a decoy
receptor and prevents RANKL from binding to RANK, resulting in the
suppression of bone resorption through the reduction of both
osteoclastgenesis and osteoclast activity (16). OPG-deficient mice and
RANKL-overexpressing transgenic mice reportedly suffer from severe
osteoporosis (17,18). Therefore, it is generally
recognized that RANK/RANKL/OPG axis is a major regulatory system
for bone remodeling (19). On the
other hand, PGs are lipid mediators with various functions and play
crucial roles in the pathophysiological responses of skeletal
tissue (20). Among them,
prostaglandin F2α (PGF2α) is a potent
stimulator of bone resorption and promotes osteoclast formation
(20). In addition, accumulating
evidence indicates that PGF2α also takes part in the
process of bone formation (21).
We have previously shown that PGF2α stimulates the
synthesis of OPG through the activation of p44/p42
mitogen-activated protein (MAP) kinase, p38 MAP kinase and stress
activated protein kinase/c-Jun N-terminal kinase (SAPK/JNK)
in osteoblast-like MC3T3-E1 cell (22).
In the present study, we investigated the effects of
EGCG or CGA on the PGF2α-stimulated OPG synthesis in
osteoblast-like MC3T3-E1 cells. We herein show that EGCG but not
CGA upregulates the PGF2α-stimulated OPG synthesis in
osteoblasts.
Materials and methods
Materials
EGCG, CGA and PGF2α were purchased from
Sigma-Aldrich (St. Louis, MO, USA). A mouse OPG enzyme-linked
immunosorbent assay (ELISA) kit was obtained from R&D Systems,
Inc. (Minneapolis, MN, USA). Phospho-specific p44/p42 MAP kinase
antibodies, p44/p42 MAP kinase antibodies, phospho-specific p38 MAP
kinase antibodies, p38 MAP kinase antibodies, phospho-specific
SAPK/JNK antibodies and SAPK/JNK antibodies were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA).
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibodies were
obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
The enhanced chemiluminescence (ECL) Western Blotting Detection
system was obtained from GE Healthcare Life Sciences (Little
Chalfont, UK). Other materials and chemicals were obtained from
commercial sources. EGCG was dissolved in dimethyl sulfoxide. CGA
and PGF2α were dissolved in ethanol. The maximum
concentration of dimethyl sulfoxide or ethanol was 0.1%, which did
not affect either the assay for OPG or western blot analysis.
Cell culture
Cloned osteoblast-like MC3T3-E1 cells that have been
derived from newborn mouse calvaria (23) were maintained as previously
described (24). Briefly, the
cells were cultured in α-minimum essential medium (α-MEM)
containing 10% fetal bovine serum (FBS) at 37°C in a humidified
atmosphere of 5% CO2/95% air. The cells were seeded into
35-mm diameter dishes (5×104 cells/dish) or 90-mm
diameter dishes (2×105 cells/dish) in α-MEM containing
10% FBS. After 5 days, the medium was exchanged for α-MEM
containing 0.3% FBS. The cells were used for experiments after 48
h.
Assay for OPG
The cultured cells were pretreated with various
doses of EGCG or CGA for 60 min, and then stimulated by 10 µM of
PGF2α or vehicle in 1 ml of α-MEM containing 0.3% FBS
for the indicated periods. The conditioned medium was collected at
the end of incubation, and the OPG concentration was then measured
using a mouse OPG ELISA kit according to the manufacturer's
protocol.
Real-time (RT)-PCR
The cultured cells were pretreated with 10 µM of
EGCG or vehicle for 60 min, and then stimulated by 10 µM of
PGF2α or vehicle in α-MEM containing 0.3% FBS for 6 h.
Total RNA was isolated and transcribed into complementary DNA using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Carlsbad, CA, USA) and OmniScript Reverse Transcriptase kit
(Qiagen, Inc., Valencia, CA, USA), respectively. RT-PCR was
performed using a LightCycler system (version 3.5; Roche
Diagnostics, Basel, Switzerland) in capillaries and FastStart DNA
Master SYBR Green I provided with the kit (Roche Diagnostics).
Sense and antisense primers for mouse OPG mRNA were purchased from
Takara Bio, Inc. (primer set ID: MA026526; Otsu, Japan), while
mouse GAPDH mRNA primers were synthesized based on the report of
Simpson et al (25). The
amplified products were determined using a melting curve analysis.
The OPG mRNA levels were normalized to those of GAPDH mRNA.
Western blot analysis
The cultured cells were pretreated with various
doses of EGCG for the indicated periods, and then stimulated by 10
µM of PGF2α or vehicle in α-MEM containing 0.3% FBS for
the indicated periods. The cells were washed twice with
phosphate-buffered saline and then lysed, homogenized and sonicated
in a lysis buffer containing 62.5 mM Tris/HCl (pH 6.8), 2% sodium
dodecyl sulfate (SDS), 50 mM dithiothreitol and 10% glycerol.
SDS-polyacrylamide gel electrophoresis (PAGE) was performed using
the method of Laemmli (26) in 10%
polyacrylamide gels. The proteins were fractionated and transferred
onto an Immun-Blot polyvinylidene difluoride (PVDF) membranes
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). The membranes were
blocked with 5% fat-free dry milk in Tris-buffered saline-Tween
(TBS-T; 20 mM Tris-HCl, pH 7.6, 137 mM NaCl, 0.1% Tween-20) for 2 h
prior to incubation with primary antibodies. A Western blot
analysis was performed as described previously (27) using antibodies against
phospho-specific p44/p42 MAP kinase, p44/p42 MAP kinase,
phospho-specific p38 MAP kinase, p38 MAP kinase, phospho-specific
SAPK/JNK, SAPK/JNK or GAPDH as primary antibodies at a dilution of
1:1,000 in 5% milk in TBS-T overnight at 4°C. Goat anti-rabbit IgG
horseradish peroxidase-labeled antibodies (KPL, Inc., Gaithersburg,
MD, USA) were used as secondary antibodies at a dilution of 1:1,000
in 5% milk in TBS-T for 1 h at room temperature. The peroxidase
activity on the PVDF sheet was visualized on X-ray film by means of
the ECL Western Blotting Detection system.
Determinations of the absorbance for
ELISA and the densitometric analysis for western blotting
The absorbance of the ELISA samples was measured at
450 nm with the EL 340 Bio Kinetic Reader (Bio-Tek Instruments,
Inc., Winooski, VT, USA). Densitometric analysis was performed
using a scanner and image analysis software package (ImageJ version
1.48; National Institutes of Health, Bethesda, MD, USA). The
phosphorylated protein levels were calculated as follows: the
background-subtracted signal intensity of each phosphorylation
signal was respectively normalized to the total protein signal and
plotted as the fold increase in comparison with that of the control
cells without treatment or stimulation.
Statistical analysis
The data were analyzed by an ANOVA followed by
Bonferroni method for multiple comparisons between pairs, and a
p-value <0.05 was considered to indicate statistically
significant difference. All data are presented as the mean ±
standard error of the mean (SEM) of triplicate determinations from
three independent cell preparations.
Results
Effects of EGCG or CGA on the
PGF2α-stimulated OPG release in MC3T3-E1 cells
In our previous study (22), we have shown that PGF2α
stimulates OPG synthesis in osteoblast-like MC3T3-E1 cells.
Therefore, we first examined the effect of EGCG on the
PGF2α-stimulated OPG release in MC3T3-E1 cells. EGCG
significantly enhanced the PGF2α-stimulated OPG release
time-dependently up to 48 h (Fig.
1A). The PGF2α-stimulated OPG release was
significantly amplified by EGCG in a dose-dependent manner in the
range between 10 and 30 µM. The maximum effect of EGCG on the
release of OPG was observed at 30 µM, which caused an approximate
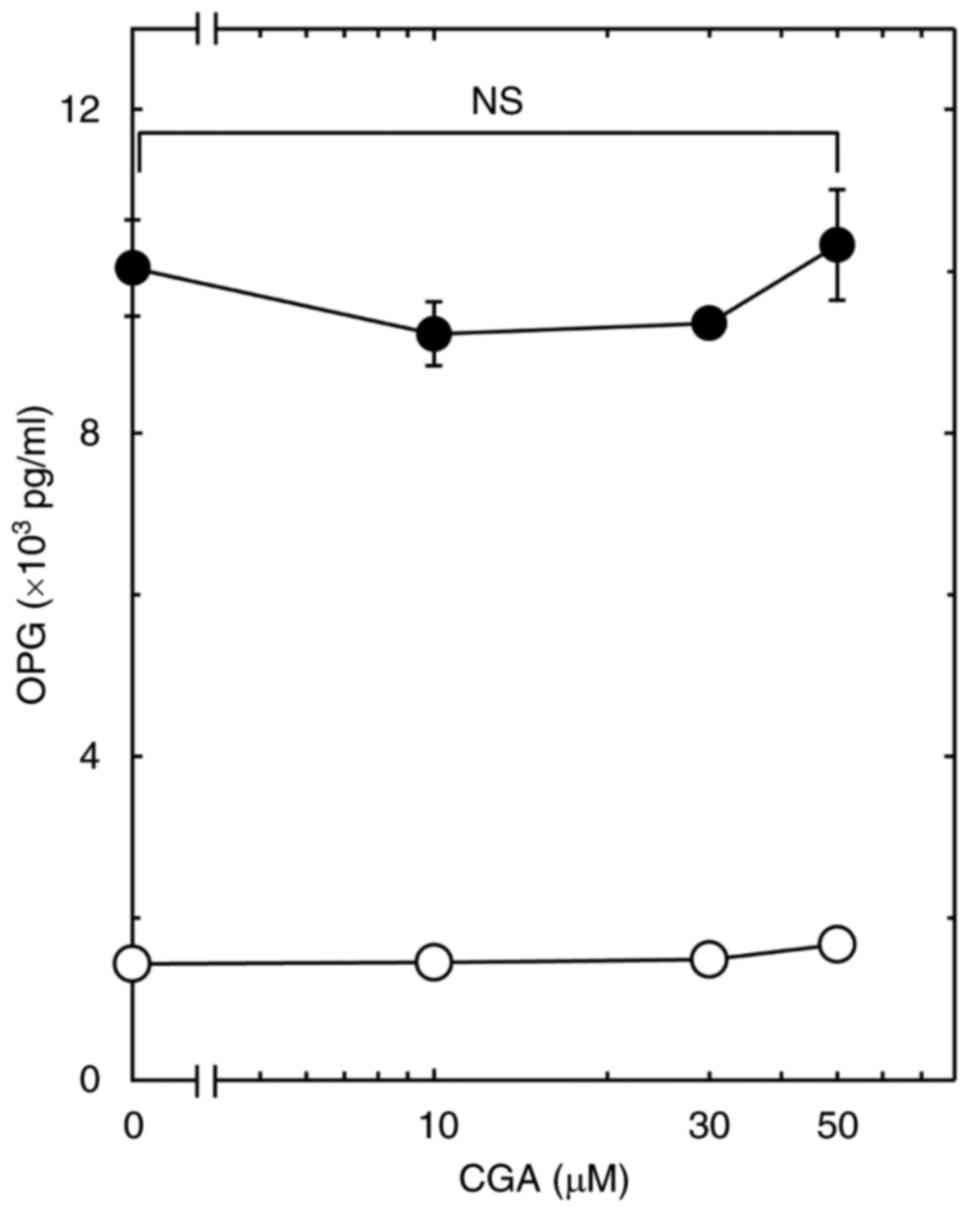
75% increase in the PGF2α-effect (Fig. 1B). On the contrary, CGA had little
effect on the PGF2α-stimulated OPG release up to 50 µM
(Fig. 2).
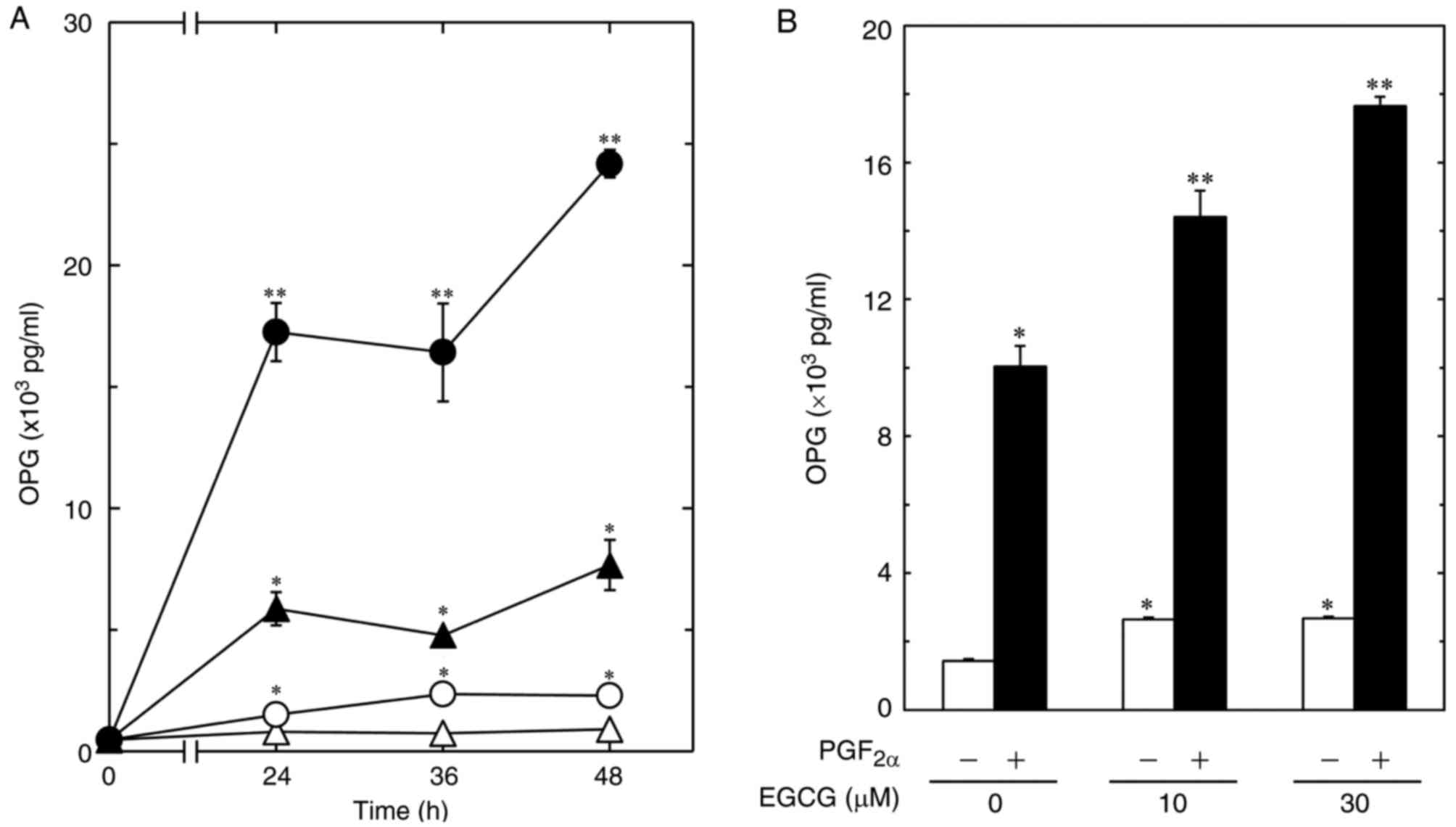 | Figure 1.Effect of EGCG on the
PGF2α-stimulated OPG release in MC3T3-E1 cells. (A) The
cultured cells were pretreated with 30 µM of EGCG (●, ○) or vehicle
(▲, Δ) for 60 min, and then stimulated by 10 µM of PGF2α
(closed symbols) or vehicle (open symbols) for the indicated
periods. (B) The cultured cells were pretreated with various doses
of EGCG for 60 min, and then stimulated by 10 µM of
PGF2α (closed bars) or vehicle (open bars) for 48 h. The
OPG concentrations in the culture medium were determined using
ELISA. Each value represents the mean ± SEM of triplicate
determinations from three independent cell preparations.
*P<0.05, compared to the value of control. **P<0.05, compared
to the value of PGF2α alone. EGCG, (−)-epigallocatechin
gallate; PGF2α, prostaglandin F2α; OPG,
osteoprotegerin; ELISA, enzyme-linked immunosorbent assay. |
Effect of EGCG on the PGF2α-induced
expression levels of OPG mRNA in MC3T3-E1 cells
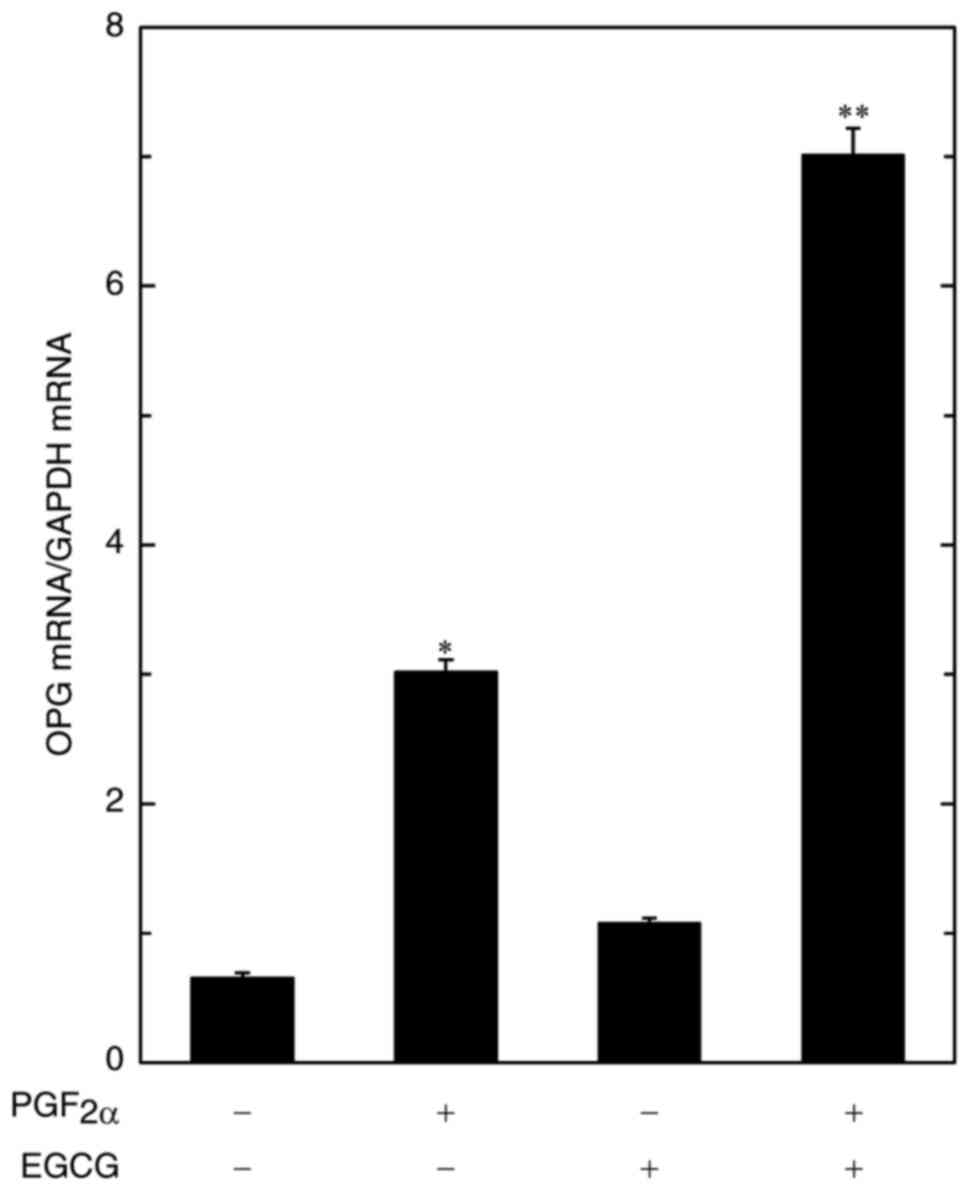
In order to investigate whether the amplification by
EGCG of the PGF2α-stimulated OPG release is mediated via
transcriptional events in osteoblast-like MC3T3-E1 cells, we
examined the effect of EGCG on the PGF2α-induced
expression levels of OPG mRNA. EGCG at 10 µM significantly
strengthened the PGF2α-induced OPG mRNA expression
(Fig. 3).
Effects of EGCG on the
PGF2α-stimulated phosphorylation of p44/p42 MAP kinase, p38 MAP
kinase and SAPK/JNK in MC3T3-E1 cells
As for the intracellular signaling of
PGF2α in OPG synthesis, we have recently demonstrated
that p44/p42 MAP kinase, p38 MAP kinase and SAPK/JNK act as
positive regulators in osteoblast-like MC3T3-E1 cells (22). Therefore, we next examined the
effects of EGCG on the PGF2α-stimulated phosphorylation
of p44/p42 MAP kinase, p38 MAP kinase and SAPK/JNK in these cells.
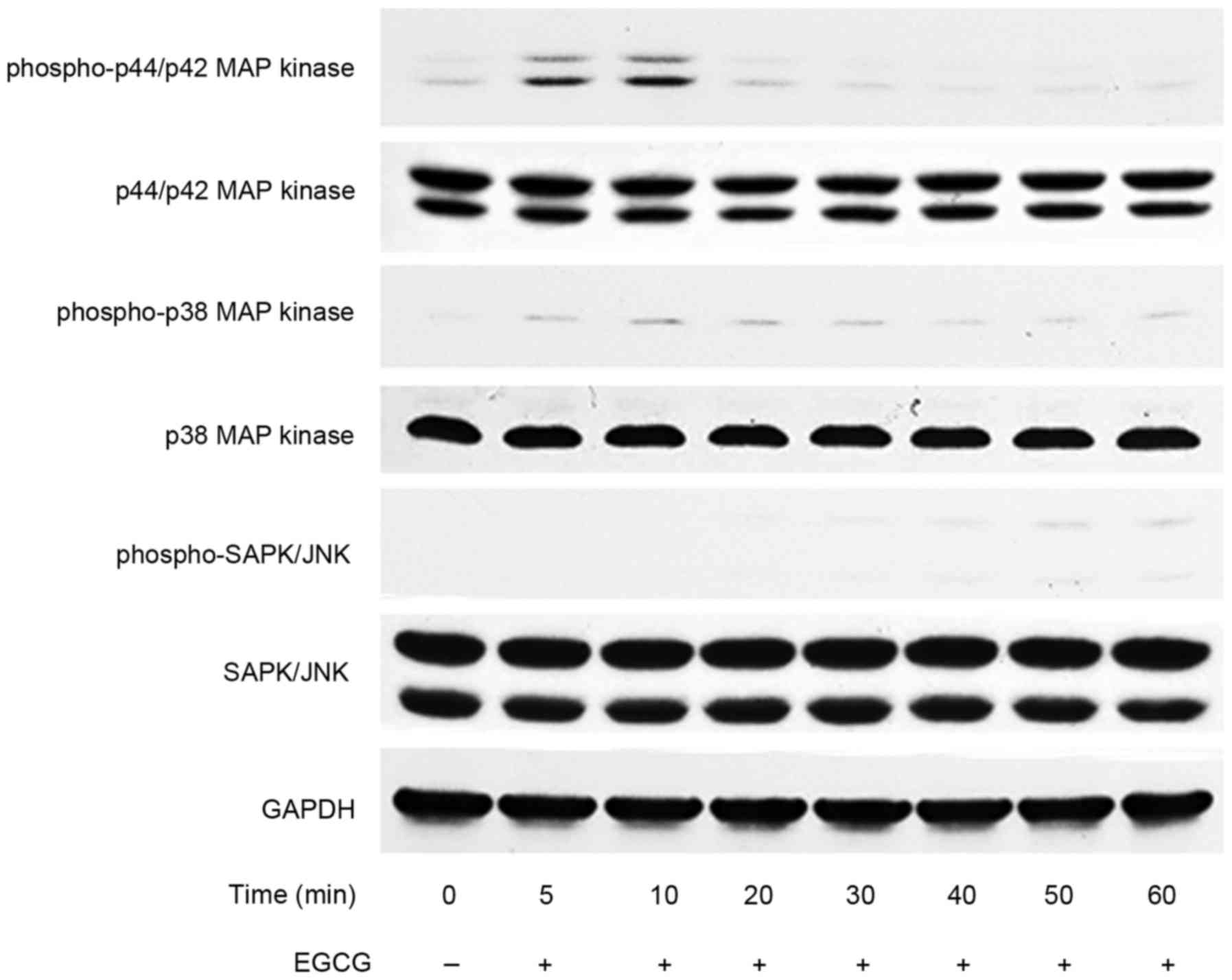
However, EGCG failed to strengthen the PGF2α-induced
phosphorylation of p44/p42 MAP kinase (Fig. 4A), p38 MAP kinase (Fig. 4B) or SAPK/JNK (Fig. 4C) over the range 10 to 30 µM. EGCG
by itself markedly induced the phosphorylation of p44/p42 MAP
kinase up to 10 min, and the status decreased thereafter, whereas
EGCG hardly affected the phosphorylation status of p38 MAP kinase
or SAPK/JNK within 60 min (Fig.
5).
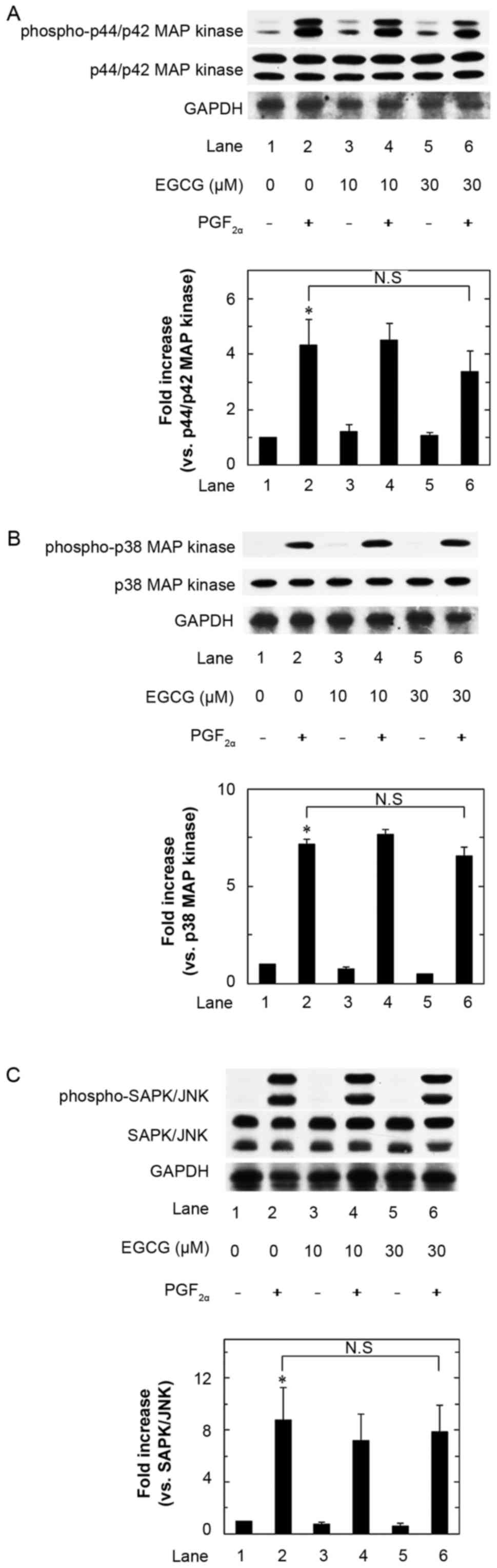 | Figure 4.Effects of EGCG on the
PGF2α-stimulated phosphorylation of p44/p42 MAP kinase
(A), p38 MAP kinase (B) and SAPK/JNK (C) in MC3T3-E1 cells. The
cultured cells were pretreated with various doses of EGCG for 60
min, and then stimulated by 10 µM of PGF2α or vehicle
for 20 min (A and C) or 10 min (B). The cell extracts were then
subjected to SDS-PAGE with subsequent Western blot analysis with
antibodies against phospho-specific p44/p42 MAP kinase, p44/p42 MAP
kinase, phospho-specific p38 MAP kinase, p38 MAP kinase,
phospho-specific SAPK/JNK or SAPK/JNK. The histogram shows
quantitative representation of the levels of
PGF2α-stimulated phosphorylation obtained from a laser
densitometric analysis of three independent experiments. Each value
represents the mean ± SEM of triplicate determinations from three
independent cell preparations. *P<0.05, compared to the value of
control (lane 1). N.S designates no significant difference between
the indicated pairs. EGCG, (−)-epigallocatechin gallate;
PGF2α, prostaglandin F2α; MAPK,
mitogen-activated protein kinase; SAPK/JNK, stress activated
protein kinase/c-Jun N-terminal kinase |
Discussion
In the present study, we demonstrated that EGCG,
most abundant catechin in green tea, significantly enhanced the
PGF2α-stimulated OPG release in osteoblast-like MC3T3-E1
cells. In addition, we showed that EGCG increased the OPG mRNA
expression levels induced by PGF2α. Based on these
findings, it is most likely that the amplification by EGCG of the
PGF2α-stimulated OPG release is mediated through a
transcriptional event. To the best of our knowledge, this is
probably the first report demonstrating the amplification of
PGF2α-stimulated OPG synthesis by EGCG in osteoblast
lineage. By contrast, CGA, which is contained abundantly in coffee,
did not affect the PGF2α-induced OPG release in MC3T3-E1
cells. We have previously shown that CGA as well as EGCG enhances
the TNF-α-stimulated IL-6 synthesis in these cells (14). Thus, it is likely that the effects
of CGA on the IL-6 synthesis in osteoblasts would be different from
EGCG in each stimulator or the responsive output.
In addition, we next investigated the intracellular
signaling mechanisms underlying the effect of EGCG on the
PGF2α-stimulated OPG synthesis in osteoblast-like
MC3T3-E1 cells. It is generally established that the MAP kinase
superfamily plays a pivotal role in a variety of cellular functions
including proliferation, differentiation and survival (28). Three major MAP kinases including
p44/p42 MAP kinase, p38 MAP kinase and SAPK/JNK, are generally
recognized as central elements used by mammalian cells to
transducer diverse messages (29).
We have previously shown that PGF2α stimulates OPG
synthesis via activation of p44/p42 MAP kinase, p38 MAP kinase and
SAPK/JNK in osteoblast-like MC3T3-E1 cells (22). However, we found that EGCG hardly
affected the PGF2α-induced phosphorylation of p44/p42
MAP kinase, p38 MAP kinase or SAPK/JNK in these cells. It is firmly
established that MAP kinases are activated through the
phosphorylation of threonine and tyrosine residues induced by the
responsible dual specificity kinase, also known as MAP kinase
kinase (30,31). Thus, it seems unlikely that the
modulations of these MAP kinases activities are involved in the
enhancement by EGCG of the PGF2α-stimulated OPG
synthesis in osteoblast-like MC3T3-E1 cells. Regarding IL-6
synthesis, it is likely that EGCG might act at a point downstream
of the MAP kinases or another target, directing the amplification
of IL-6 synthesis. Indeed, we found that EGCG by itself markedly
induced the phosphorylation of p44/p42 MAP kinase up to 10 min, and
the status decreased thereafter, whereas EGCG hardly affected the
phosphorylation status of p38 MAP kinase or SAPK/JNK within 60 min.
It seems likely that EGCG affects the cell function of
osteoblast-like MC3T3-E1 cells through the p44/p42 MAP
kinase-mediating signaling. In addition, in order to investigate
the PGF2α-effects on OPG synthesis in EGCG-affected
osteoblasts compared with those in naive osteoblasts,
osteoblast-like MC3T3-E1 cells were pretreated with EGCG for 60 min
before PGF2α stimulation. Further investigations are
necessary to elucidate the exact mechanism of EGCG in osteoblasts.
Taking our findings into account as a whole, it is most likely that
not CGA but EGCG enhances the PGF2α-stimulated OPG
synthesis in osteoblast-like MC3T3-E1 cells, and that the
amplifying effect of EGCG on the IL-6 synthesis is independent from
the activation of p44/p42 MAP kinase, p38 MAP kinase or
SAPK/JNK.
EGCG is a natural polyphenol abundantly found in
green tea, and possesses numerous favorable effects on the health
of human being through the actions of anti-oxidation (6). Accumulating evidence indicate that
green tea consumption prevents both age-related bone loss and
fracture in elderly people (6). It
is well known that PGF2α has multiple effects on bone
metabolism, and that PGF2α acts as a mediator of bone
remodeling, resulting in the regulation of bone turnover (21). Osteoblast-producing OPG functions
as a decoy receptor of RANKL, and blocks RANKL-RANK interaction
which is critical for the osteoclastgenesis and the activation of
osteoclasts (16,32). Based on our present findings
demonstrating the upregulation by EGCG of OPG synthesis induced by
PGF2α in osteoblasts, it is possible that EGCG directs
the bone metabolism toward the increase of formation but the
suppression of resorption. Therefore, the effect of EGCG showing
here might provide a novel potential aspect for the favorable
actions of EGCG-containing beverages in bone health for the elderly
peoples. Further investigations would be needed to clarify the
detailed mechanism of EGCG underlying the OPG synthesis in
osteoblasts.
In conclusion, our results strongly suggest that
EGCG but not CGA enhances the PGF2α-stimulated OPG
synthesis in osteoblasts.
Acknowledgements
We are very grateful to Yumiko Kurokawa for her
skillful technical assistance. This investigation was supported in
part by a Grant-in-Aid for Scientific Research (26462289, 15K10487)
from the Ministry of Education, a Grant-in-Aid for Scientific
Research (H25-Aging-General-004) from the Ministry of Health,
Labour and Welfare, and the Research Funding for Longevity Sciences
(25–4, 26–12)
from National Center for Geriatrics and Gerontology (NCGG),
Japan.
References
|
1
|
Thielecke F and Boschmann M: The potential
role of green tea catechins in the prevention of the metabolic
syndrome-a review. Phytochemistry. 70:11–24. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shimizu M, Adachi S, Masuda M, Kozawa O
and Moriwaki H: Cancer chemoprevention with green tea catechins by
targeting receptor tyrosine kinases. Mol Nutr Food Res. 55:832–843.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Higdon JV and Frei B: Coffee and health: A
review of recent human research. Crit Rev Food Sci Nutr.
46:101–123. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Harborne JB and Williams CA: Advances in
flavonoid research since 1992. Phytochemistry. 55:481–504. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
George SE, Ramalakshmi K and Rao LJ Mohan:
A perception on health benefits of coffee. Crit Rev Food Sci Nutr.
48:464–486. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shen CL, Yeh JK, Cao JJ and Wang JS: Green
tea and bone metabolism. Nutr Res. 29:437–456. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Karsenty G and Oury F: Biology without
walls: The novel endocrinology of bone. Annu Rev Physiol.
74:87–105. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Parfitt AM: Targeted and nontargeted bone
remodeling: Relationship to basic multicellular unit origination
and progression. Bone. 30:5–7. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kwak SC, Lee C, Kim JY, Oh HM, So HS, Lee
MS, Rho MC and Oh J: Chlorogenic acid inhibits osteoclast
differentiation and bone resorption by down-regulation of receptor
activator of nuclear factor kappa-B ligand-induced nuclear factor
of activated T cells c1 expression. Biol Pharm Bull. 36:1779–1786.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Folwarczna J, Pytlik M, Zych M, Cegiela U,
Nowinska B, Kaczmarczyk-Sedlak L, Sliwinski L, Trzeciak H and
Trzeciak HI: Effects of caffeic and chlorogenic acids on the rat
skeletal system. Eur Rev Med Pharmacol Sci. 19:682–693.
2015.PubMed/NCBI
|
|
11
|
Takai S, Matsushima-Nishiwaki R, Adachi S,
Natsume H, Minamitani C, Mizutani J, Otsuka T, Tokuda H and Kozawa
O: (−)-Epigallocatechin gallate reduces platelet-derived growth
factor-BB-stimulated interleukin-6 synthesis in osteoblasts:
Suppression of SAPK/JNK. Mediators Inflamm. 2008:2918082008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tokuda H, Takai S, Hanai Y,
Matsushima-Nishiwaki R, Yamauchi J, Harada A, Hosoi T, Ohta T and
Kozawa O: (−)-Epigallocatechin gallate inhibits basic fibroblast
growth factor-stimulated interleukin-6 synthesis in osteoblasts.
Horm Metab Res. 40:674–678. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tokuda H, Takai S, Hanai Y,
Matsushima-Nishiwaki R, Hosoi T, Harada A, Ohta T and Kozawa O:
(−)-Epigallocatechin gallate suppresses endothelin-1-induced
interleukin-6 synthesis in osteoblasts: inhibition of p44/p42 MAP
kinase activation. FEBS Lett. 581:1311–1316. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamamoto N, Tokuda H, Kuroyanagi G,
Kainuma S, Ohguchi R, Fujita K, Matsushima-Nishiwaki R, Kozawa O
and Otsuka T: Amplification by (−)-epigallocatechin gallate and
chlorogenic acid of TNF-α-stimulated interleukin-6 synthesis in
osteoblasts. Int J Mol Med. 36:1707–1712. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kostenuik PJ and Shalhoub V:
Osteoprotegerin: A physiological and pharmacological inhibitor of
bone resorption. Curr Pharm Des. 7:613–635. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Simonet WS, Lacey DL, Dunstan CR, Kelley
M, Chang MS, Lüthy R, Nguyen HQ, Wooden S, Bennett L, Boone T, et
al: Osteoprotegerin: A novel secreted protein involved in the
regulation of bone density. Cell. 89:309–319. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mizuno A, Amizuka N, Irie K, Murakami A,
Fujise N, Kanno T, Sato Y, Nakagawa N, Yasuda H, Mochizuki S, et
al: Severe osteoporosis in mice lacking osteoclastogenesis
inhibitory factor/osteoprotegerin. Biochem Biophys Res Commun.
247:610–615. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mizuno A, Kanno T, Hoshi M, Shibata O,
Yano K, Fujise N, Kinosaki M, Yamaguchi K, Tsuda E, Murakami A, et
al: Transgenic mice overexpressing soluble osteoclast
differentiation factor (sODF) exhibit severe osteoporosis. J Bone
Miner Metab. 20:337–344. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tat S Kwan, Padrines M, Théoleyre S,
Heymann D and Fortun Y: IL-6, RANKL, TNF-α/IL-1: Interrelations in
bone resorption pathophysiology. Cytokine Growth Factor Rev.
15:49–60. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hikiji H, Takato T, Shimizu T and Ishii S:
The roles of prostanoids, leukotrienes, and platelet-activating
factor in bone metabolism and disease. Prog Lipid Res. 47:107–126.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Agas D, Marchetti L, Hurley MM and
Sabbieti MG: Prostaglandin F2α: A bone remodeling mediator. J Cell
Physiol. 228:25–29. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kuroyanagi G, Tokuda H,
Matsushima-Nishiwaki R, Kondo A, Mizutani J, Kozawa O and Otsuka T:
Resveratrol suppresses prostaglandin F2α-induced osteoprotegerin
synthesis in osteoblasts: Inhibition of the MAP kinase signaling.
Arch Biochem Biophys. 542:39–45. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sudo H, Kodama HA, Amagai Y, Yamamoto S
and Kasai S: In vivo differentiation and calcification in a new
clonal osteogenic cell line derived from newborn mouse calvaria. J
Cell Biol. 96:191–198. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kozawa O, Tokuda H, Miwa M, Kotoyori J and
Oiso Y: Cross-talk regulation between cyclic AMP production and
phosphoinositide hydrolysis induced by prostaglandin E2 in
osteoblast-like cells. Exp Cell Res. 198:130–134. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Simpson DA, Feeney S, Boyle C and Stitt
AW: Retinal VEGF mRNA measured by SYBR green I fluorescence: A
versatile approach to quantitative PCR. Mol Vis. 6:178–183.
2000.PubMed/NCBI
|
|
26
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kato K, Ito H, Hasegawa K, Inaguma Y,
Kozawa O and Asano T: Modulation of the stress-induced synthesis of
hsp27 and alpha B-crystallin by cyclic AMP in C6 rat glioma cells.
J Neurochem. 66:946–950. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kyriakis JM and Avruch J: Mammalian
mitogen-activated protein kinase signal transduction pathways
activated by stress and inflammation. Physiol Rev. 81:807–869.
2001.PubMed/NCBI
|
|
29
|
Widmann C, Gibson S, Jarpe MB and Johnson
GL: Mitogen-activated protein kinase: Conservation of a
three-kinase module from yeast to human. Physiol Rev. 79:143–180.
1999.PubMed/NCBI
|
|
30
|
Dhanasekaran N and Reddy E Premkumar:
Signaling by dual specificity kinases. Oncogene. 17:1447–1455.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin A, Minden A, Martinetto H, Claret FX,
Lange-Carter C, Mercurio F, Johnson GL and Karin M: Identification
of a dual specificity kinase that activates the Jun kinases and
p38-Mpk2. Science. 14:286–290. 1995. View Article : Google Scholar
|
|
32
|
Greenfield EM, Bi Y and Miyauchi A:
Regulation of osteoclast activity. Life Sci. 65:1087–1102. 1999.
View Article : Google Scholar : PubMed/NCBI
|