Introduction
Cardiac arrest (CA) is a very dangerous state of
shock and is a common cause of mortality and disability because the
whole body suffers from severe ischemia, including the cerebellum,
thalamus, hippocampus, and cortex (1,2).
Even if CA patients receive timely restoration of spontaneous
circulation (ROSC), ischemia still may occur leading to an
ischemia/reperfusion (I/R) injury, which includes apoptosis and
necrosis of neurons and activation of multiple pathogenic
mechanisms (1). The poor outcomes
in CA patients due to these distinct pathologies is termed post-CA
syndrome (2). The probability that
CA patients will recover after ROSC is increasing because of
increasing public knowledge of cardiopulmonary resuscitation (CPR),
significant improvements in the construction of emergency networks,
and rapid progress in rescue technology. Early diagnosis and proper
evaluation of the severity of brain damage is necessary for good
prognoses in CA patients. However, the exact etiology of I/R injury
remains poorly understood in CA, and the cerebral pathology of I/R
injury and the molecular mechanisms of I/R after CA-ROSC remains to
be elucidated.
Thousands of messenger RNAs (mRNAs) are continuously
transcribed and guide the translation of various proteins in the
adult mammalian brain during pathological states. The
transcriptional and translational machinery is comprised of a
variety of molecules, such as DNA methylases, transcription
factors, ribosomes, and RNA polymerases. Recently, noncoding RNAs
have been shown to potently regulate gene expression at the levels
of transcription and translation (3). These noncoding RNAs participate in
cellular, developmental, and disease processes by regulating RNA
and protein levels, imprinting, enhancer function, and
transcription (4–7). Long noncoding RNAs (lncRNAs) are a
relatively new class of RNA molecules that are longer than 200
nucleotides and much less well characterized than microRNAs.
Cerebral lncRNA profiles have been shown to be markedly altered in
many pathological states, to play important roles in the
pathophysiology of central nervous system and brain-related
diseases and disorders, and to have profound effects on disease
outcomes (8–11). Notably, studies have also shown
that lncRNAs are associated with I/R brain injury (11,12).
However, cerebral lncRNA profiles after CA-ROSC,
including a complete list of lncRNAs expressed in the brain, have
yet to be elucidated. In addition, the potential roles and effects
of these lncRNAs on mRNA expression during the early stage of
reperfusion have yet to be determined. Undoubtedly, understanding
lncRNA and mRNA network profiles in post-CA brains could inform the
development of therapeutics to prevent secondary neuronal death
caused by CA. In this study, we used a rat model to explore lncRNA
and mRNA network profiles in cortical pathophysiological processes
to further understand the etiology and molecular mechanisms of
cerebral pathology during an I/R injury.
Materials and methods
Animals
Male Wistar rats (15–16 months old and 305.5–400.3
g) were obtained from the Experimental Animal Center, Sun Yat-Sen
University, Guangdong Province (Guangzhou, China). Animals were
housed in a pathogen-free laboratory at 20–22°C with a 10 h light
and 14 h dark cycle. The animal studies were approved by the
Institutional Animal Care and Use Committee of Sun Yat-sen
University and the procedures used were in accordance with the
Animal Research Reporting for in vivo Experiments Guidelines
on animal research (13).
Induction of ventricular fibrillation
(VF)
After fasting overnight (with access to water), 14
healthy male Wistar rats were anesthetized by intraperitoneal
pentobarbital injection (30 mg/kg; Sigma, Natick, MA, USA). VF was
induced as described previously (14,15).
VF was successfully induced by alternating current (50 Hz, 6 V) for
10 sec with external transthoracic needles. After successful
induction of VF, the ventilator was disconnected. Eight min after
VF, manual chest compressions were performed on the rats along with
mechanical ventilation. Rats were maintained for 24 h after
successful ROSC (CA, n=14). Seven sham-operated rats served as the
blank control (BC, n=7). 5 rats in model group and 4 in sham group
were anesthetized by intraperitoneal pentobarbital injection (40
mg/kg; Sigma), and were sacrificed by decapitation at ROSC 2 h, the
cerebral cortex tissue were harvested for RNA analysis. The rest
rats were survival to 24 h after ROSC and their neurological
deficits scores (NDS) were assessed from 0 (no observed
neurological deficit) to 500 (death or brain death) (14,15)
by 2 investigators who were blinded to the treatment. All of the
other animals were sacrificed at 24 h after ROSC; the brains were
removed and were fixed using paraffin for Nissl and TdT-mediated
dUTP-biotin nick end labeling (TUNEL) staining, the apoptotic cells
were counted randomly 6 fields in sequence slice in each group. All
rats were anesthetized by intraperitoneal pentobarbital injection
(40 mg/kg) and were sacrificed by decapitation.
Tissue harvest and RNA extraction
The rat brains were harvested and the cerebral
cortices were dissected quickly on ice. Samples of cerebral cortex
tissue were quick-frozen in liquid nitrogen and stored at −80°C for
future use. Total RNAs were extracted with TRIzol reagent,
quantified (NanoDrop ND-2000; Thermo Fisher Scientific, Inc.,
Pittsburgh, PA, USA), and analyzed for integrity (Agilent
Bioanalyzer 2100; Agilent Technologies, Inc., Santa Clara, CA,
USA). RNA labeling and microarray hybridization were performed
according to standard protocols provided by the manufacturer.
LncRNA and mRNA microarray expression
profiling
Total RNAs (100 ng) were labeled with the mRNA
Complete Labeling and Hyb kit and hybridized on OE Biotech Rat
lncRNA Microarray v2.0 8×60K (both Agilent Technologies, Inc.).
This microarray contained 40,367 probes for 30,367 rat mRNAs and
10,000 rat lncRNAs derived from authoritative databases, including
Ensembl, RefSeq, Ultra-conserved region encoding LncRNA (UCR), and
lncRNAdb.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
To validate the differentially expressed lncRNAs,
quantitative RT-PCR was performed using a two-step reaction, which
included RT and PCR, according to the manufacturer's instructions.
Each RT reaction consisted of 0.5 µg of RNA, 2 µl of PrimerScript
Buffer, 0.5 µl of oligo dT, 0.5 µl of random 6-mers, and 0.5 µl of
PrimerScript RT Enzyme Mix I (Takara Bio, Inc., Otsu, Japan) in a
total volume of 10 µl. RT reactions were performed in a
GeneAmp® PCR System 9700 (Applied Biosystems; Thermo
Fisher Scientific, Inc.) for 15 min at 37°C followed by heat
inactivation for 5 sec at 85°C. The 10 µl RT reaction mix was then
diluted 10-fold in nuclease-free water and stored at −20°C. RT-qPCR
was performed with 10 µl of a PCR reaction mixture that contained 1
µl of cDNA, 5 µl of 2X LightCycler® 480 SYBR-Green I
Master Mix, 0.2 µl of forward primer, 0.2 µl of reverse primer, and
3.6 µl of nuclease-free water in a LightCycler® 480 II
Real-time PCR Instrument (both Roche Applied Science, Rotkreuz,
Switzerland). Reactions were incubated in a 384-well optical plate
(Roche) at 95°C for 10 min followed by 40 cycles of 95°C for 10 sec
and 60°C for 30 sec. Each sample was run in triplicate and a
melting curve analysis was performed at the end of each PCR cycle
to specifically validate generation of the expected PCR product.
The expression levels of lncRNAs were normalized to
glyceraldehyde-3-phosphate dehydrogenase expression and were
calculated using the 2−ΔΔCq method. Primers were
synthesized by Generay Biotech (Generay, Shanghai, China) based on
mRNA sequences in the NCBI database and the sequences are given in
Table I.
 | Table I.Primers for validation of
differentially expressed long noncoding RNAs. |
Table I.
Primers for validation of
differentially expressed long noncoding RNAs.
| Gene symbol | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| TCONS_00097275 |
GCCACCAGGACTGATAAT |
TGTGTGTGTGTGTGTACG |
| TCONS_00077647 |
ATGGTGATTCGCACTTGT |
TCCTTTAGCTCAGACTGGC |
| TCONS_00132873 |
GTTCAGGCAAGCTCCAAA |
ATCCGTGTCTGGAACATT |
| TCONS_00066427 |
CAGGTCTTCTTTACCACCGA |
GAGAGCAGGCAGAAGTGA |
| NR_002597.1 |
GCTGTGAGATGCAGGACAA |
AAGGTGGCTGCTGTATATT |
| TCONS_00079198 |
GAAGGAGTGTGGAACTCAAC |
CCTCTGCCATTAGACTTTGT |
| GAPDH |
GCGAGATCCCGCTAACATCA |
CTCGTGGTTCACACCCATCA |
Analyses of microarray results and
predictions of lncRNA functions
After differentially expressed mRNA and lncRNA data
from microarrays were acquired, raw signals were log2
transformed. Differentially expressed mRNAs and lncRNAs were
screened for absolute values of fold change (FC) >2 and P-values
<0.05 (Student's t-test). The differentially expressed mRNAs
were submitted to the DAVID database (http://david.abcc.ncifcrf.gov) and were classified
into different annotation groups by Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) analyses. Distinguishable
gene expression patterns were displayed by hierarchical
clustering.
Statistical analysis
All statistical analyses were performed using the
SPSS software 13.0 (SPSS, Inc., Chicago, IL, USΑ). Data were
presented as mean ± SD or proportions as appropriate. Unpaired
student t-test were performed to compare the differences of NDS,
apoptotic cells, and validating lncRNA expressions between two
groups, and correlations were calculated using Spearman's
correlation coefficient, which is the Pearson's correlation
coefficient of the indexed ranks of the two data sets. P<0.05 by
two-tailed t-test was considered to indicate a statistically
significant difference.
Results
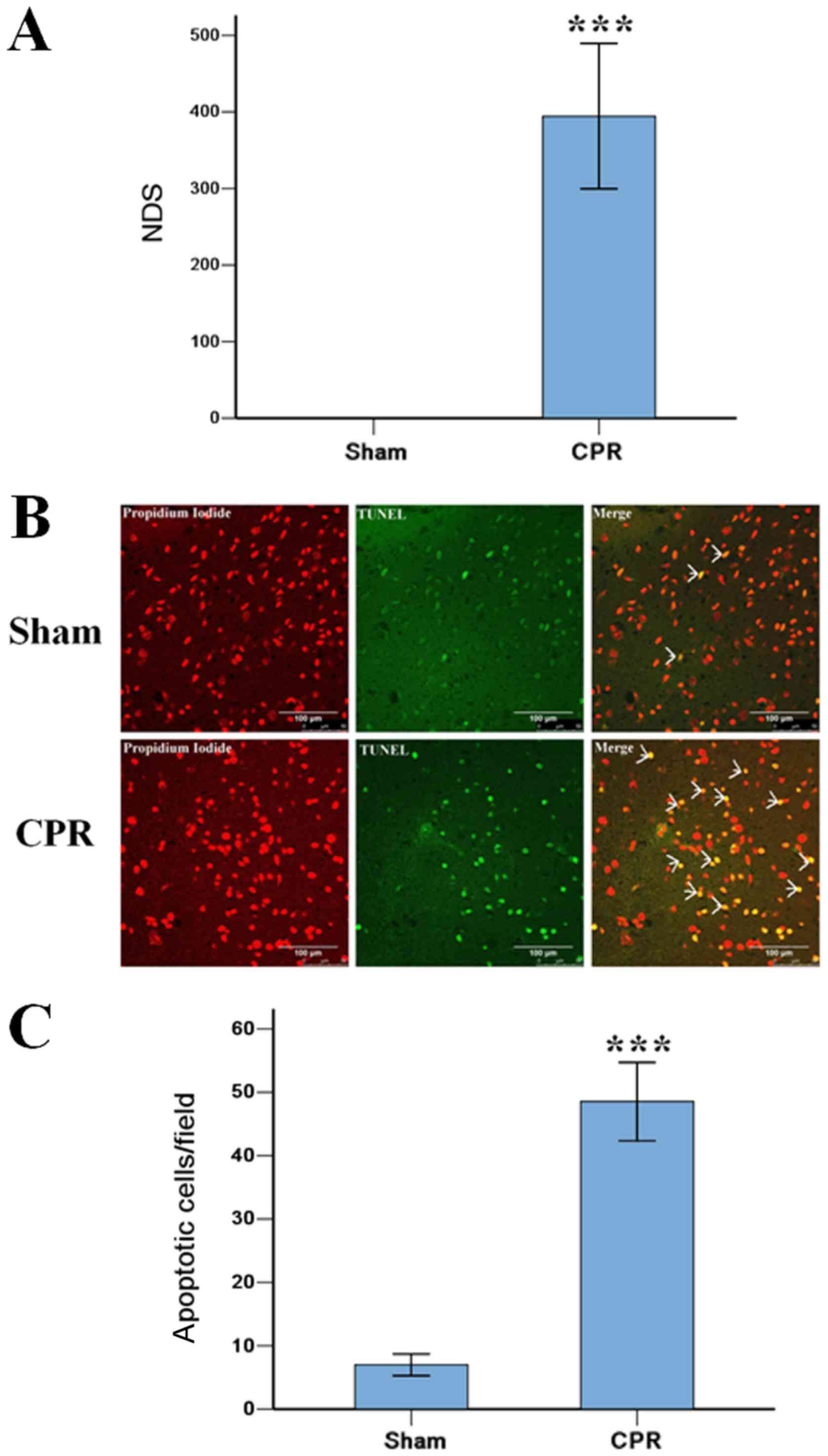
The severity of brain damage of rats
after ROSC 24 h
The induction of VF caused serious injury to the
brain in rats after ROSC 24 h in CPR group, 3 rats were dead within
24 h, and the other 6 rats were survival to 24 h. All rats in Sham
group were alive to the end of the experiment. The NDS score in CPR
group were 394±95, significantly higher than 0±0 in Sham group
(P<0.001). The TUNEL staining showed that the induction of VF
significantly increased the amount of apoptotic cells in the
cortex, compared with that in Sham group (Fig. 1).
Changes in lncRNA and mRNA expressions
in the CA and BC groups
From the lncRNA microarray, we found 58 lncRNA
transcripts that were differentially expressed. Thirty-seven of the
lncRNA transcripts were upregulated and 21 of the lncRNA
transcripts were downregulated in the CA group when compared with
the BC group (Table II). The
TCONS_00045076 (probe CUST_12461 _PI429484123) and TCONS_00087195
(probe CUST_12461_PI429484123) lncRNAs were the most upregulated
and downregulated transcripts in the CA group, respectively.
According to absolute FC values (abs), the differentially expressed
lncRNA transcripts were divided into two groups: five transcripts
with FC values between 5 and 10 (highly dysregulated lncRNAs) and
53 transcripts with FC values between 2 and 5 (less dysregulated
lncRNAs).
| Table II.Differentially expressed long
noncoding RNAs in cardiac arrest-return of spontaneous
circulation. |
Table II.
Differentially expressed long
noncoding RNAs in cardiac arrest-return of spontaneous
circulation.
| No. | lncRNA | FC (abs) | Regulation | BC | CA | Chromosome no. |
|---|
| 1 | TCONS_00045076 | 8.07 | Up |
2.60±2.54 |
5.61±0.94 | 14 |
| 2 | TCONS_00097275 | 8.00 | Up |
2.31±1.87 |
5.31±0.78 | 4 |
| 3 | TCONS_00097694 | 6.75 | Up |
2.59±2.36 |
5.34±1.05 | 4 |
| 4 | TCONS_00058731 | 6.27 | Up |
2.67±1.53 |
5.32±0.36 | 17 |
| 5 |
ENSRNOT00000031100 | 5.38 | Up |
4.59±1.48 |
7.02±0.59 | 15 |
| 6 | TCONS_00045077 | 4.47 | Up |
4.13±1.60 |
6.29±0.94 | Un_random |
| 7 | XR_146410.1 | 4.09 | Up |
4.78±1.13 |
6.81±0.96 | Un |
| 8 | TCONS_00064719 | 4.06 | Up |
4.55±1.23 |
6.57±0.89 | Un_random |
| 9 | XR_146719.1 | 3.70 | Up |
4.27±1.27 |
6.15±0.68 | Un |
| 10 |
ENSRNOT00000058957 | 3.69 | Up |
7.88±0.38 |
9.76±0.58 | X |
| 11 |
ENSRNOT00000030838 | 3.36 | Up |
8.37±0.22 |
10.12±0.53 | 3 |
| 12 | XR_146012.1 | 3.34 | Up |
7.34±0.83 |
9.08±0.59 | 5 |
| 13 | XR_146595.1 | 3.08 | Up |
3.62±0.76 |
5.24±0.47 | 14 |
| 14 | TCONS_00040687 | 3.08 | Up |
4.27±0.71 |
5.90±0.59 | 13 |
| 15 | XR_147150.1 | 3.03 | Up |
5.74±0.80 |
7.34±0.73 | Un |
| 16 | XR_146408.1 | 2.85 | Up |
5.48±0.89 |
6.99±0.61 | Un |
| 17 | TCONS_00062712 | 2.84 | Up |
3.29±1.11 |
4.80±0.27 | 18 |
| 18 | TCONS_00113107 | 2.76 | Up |
4.69±0.55 |
6.16±0.45 | 6 |
| 19 | TCONS_00122410 | 2.74 | Up |
3.52±0.40 |
4.98±0.99 | 7 |
| 20 | TCONS_00046288 | 2.68 | Up |
3.01±0.26 |
4.44±0.58 | 15 |
| 21 | TCONS_00009008 | 2.59 | Up |
3.97±0.60 |
5.34±0.61 | 1 |
| 22 | TCONS_00008734 | 2.48 | Up |
3.22±0.99 |
4.53±0.48 | 1 |
| 23 | TCONS_00066427 | 2.30 | Up |
9.68±0.75 |
10.88±0.59 | 2 |
| 24 | TCONS_00041399 | 2.28 | Up |
2.92±0.60 |
4.11±0.53 | 14 |
| 25 |
ENSRNOT00000065794 | 2.25 | Up |
3.81±0.64 |
4.98±0.47 | 8 |
| 26 | TCONS_00062192 | 2.24 | Up |
3.52±0.32 |
4.69±0.28 | 18 |
| 27 | TCONS_00051727 | 2.20 | Up |
3.46±0.86 |
4.60±0.27 | 16 |
| 28 | TCONS_00048046 | 2.20 | Up |
4.43±0.70 |
5.57±0.49 | 15 |
| 29 | FR144720 | 2.16 | Up |
3.39±0.51 |
4.50±0.59 | 1 |
| 30 |
ENSRNOT00000074021 | 2.14 | Up |
5.38±0.50 |
6.48±0.61 | 2 |
| 31 | TCONS_00132873 | 2.11 | Up |
8.84±0.69 |
9.91±0.63 | 12 |
| 32 | TCONS_00065841 | 2.10 | Up |
3.61±0.79 |
4.68±0.47 | 19 |
| 33 | TCONS_00054776 | 2.08 | Up |
4.69±0.39 |
5.75±0.35 | 16 |
| 34 | TCONS_00054777 | 2.08 | Up |
3.50±0.77 |
4.56±0.24 | 16 |
| 35 | uc.280+ | 2.07 | Up |
6.95±0.69 |
8.01±0.39 | 3 |
| 36 |
ENSRNOT00000040695 | 2.04 | Up |
6.57±0.09 |
7.60±0.32 | X_random |
| 37 | uc.129+ | 2.02 | Up |
6.72±0.44 |
7.73±0.55 | 2 |
| 38 | TCONS_00087195 | 3.74 | Down |
5.15±0.49 |
3.25±0.55 | 3 |
| 39 | TCONS_00129724 | 3.49 | Down |
4.77±0.85 |
2.96±0.57 | 8 |
| 40 | TCONS_00077647 | 3.27 | Down |
7.78±1.02 |
6.07±0.68 | 20 |
| 41 | TCONS_00072164 | 3.16 | Down |
4.96±0.34 |
3.30±1.31 | 2 |
| 42 | NR_002597.1 | 2.92 | Down |
8.97±1.07 |
7.42±0.48 | 20 |
| 43 | TCONS_00079198 | 2.73 | Down |
6.78±0.85 |
5.33±0.59 | 20 |
| 44 | TCONS_00046110 | 2.69 | Down |
6.54±0.62 |
5.11±0.99 | 15 |
| 45 | TCONS_00016640 | 2.64 | Down |
5.00±0.28 |
3.60±0.45 | 1 |
| 46 | TCONS_00106864 | 2.38 | Down |
5.60±0.27 |
4.35±0.81 | 5 |
| 47 | TCONS_00098087 | 2.37 | Down |
4.94±0.43 |
3.69±0.08 | 4 |
| 48 | TCONS_00138507 | 2.36 | Down |
5.87±0.45 |
4.63±0.95 | X |
| 49 | NR_037614.1 | 2.35 | Down |
5.90±0.22 |
4.66±0.94 | 8 |
| 50 | XR_145894.1 | 2.34 | Down |
5.56±0.72 |
4.34±0.38 | 3 |
| 51 | TCONS_00132190 | 2.23 | Down |
7.32±0.84 |
6.17±0.58 | 9 |
| 52 | TCONS_00040199 | 2.22 | Down |
5.07±0.58 |
3.92±0.68 | 13 |
| 53 | TCONS_00040514 | 2.16 | Down |
4.91±0.52 |
3.80±0.76 | 13 |
| 54 | XR_145922.1 | 2.14 | Down |
8.05±0.69 |
6.95±0.43 | 4 |
| 55 | TCONS_00062765 | 2.10 | Down |
5.11±0.73 |
4.04±0.53 | 18 |
| 56 | TCONS_00021035 | 2.05 | Down |
5.72±0.19 |
4.69±0.53 | 10 |
| 57 | TCONS_00066438 | 2.04 | Down |
4.18±0.21 |
3.15±0.68 | n/a |
| 58 | TCONS_00122316 | 2.00 | Down |
3.84±0.39 |
2.83±0.71 | 7 |
Using the same criteria that we used for the
lncRNAs, we found 258 differentially expressed mRNA transcripts.
177 of the transcripts in the CA group were upregulated and 81 of
the transcripts in the CA group were downregulated when compared
with the BC group. The NM_001106299 (A_43_P16887) and NM_001033691
(A_44_P1039994) mRNAs were the most upregulated and downregulated
transcripts in the CA group, respectively. Some of the
differentially expressed mRNAs (FC>4) are listed in Table III; the un-named mRNAs are listed
by their probe names, such as A_44_P809486.
| Table III.Differentially expressed mRNAs in
cardiac arrest-return of spontaneous circulation. |
Table III.
Differentially expressed mRNAs in
cardiac arrest-return of spontaneous circulation.
| No. | mRNA | FC (abs) | Regulation | BC | CA | Chromosome no. |
|---|
| 1 | Ahsp | 38.62 | Up |
1.98±3.43 |
7.25±2.12 | 1 |
| 2 | Ptges | 18.56 | Up |
3.27±3.51 |
7.48±0.88 | 3 |
| 3 | Egr4 | 13.98 | Up |
2.45±2.89 |
6.25±1.23 | 4 |
| 4 | Six3 | 11.17 | Up |
2.93±3.11 |
6.41±0.95 | 6 |
| 5 | Apold1 | 10.84 | Up |
4.44±2.56 |
7.87±0.94 | 4 |
| 6 | Gch1 | 10.10 | Up |
3.21±2.34 |
6.55±1.17 | 15 |
| 7 | Msantd1 |
9.23 | Up |
2.40±2.07 |
5.60±0.75 | 14 |
| 8 | A_64_P010891 |
7.52 | Up |
2.83±1.98 |
5.74±1.51 | 17 |
| 9 | Trh |
7.39 | Up |
2.30±1.37 |
5.19±0.71 | 4 |
| 10 | Nr4a1 |
6.64 | Up |
8.15±1.55 |
10.88±0.58 | 7 |
| 11 | Gdf15 |
5.72 | Up |
3.55±1.63 |
6.06±0.44 | 16 |
| 12 | Ptgs2 |
5.33 | Up |
5.71±1.31 |
8.13±0.93 | 13 |
| 13 | LOC688972 |
5.31 | Up |
3.60±0.45 |
6.01±1.69 | 19 |
| 14 | A_64_P058988 |
4.83 | Up |
3.79±0.86 |
6.07±1.28 | 19 |
| 15 | Grin3b |
4.72 | Up |
3.47±1.69 |
5.71±0.65 | 7 |
| 16 | A_44_P386579 |
4.69 | Up |
3.68±1.83 |
5.90±0.66 | 18 |
| 17 | Gadd45g |
4.66 | Up |
7.14±1.76 |
9.36±0.60 | 17 |
| 18 | A_64_P065301 |
4.65 | Up |
4.64±1.82 |
6.86±0.89 | Un |
| 19 | A_64_P134263 |
4.63 | Up |
3.99±1.67 |
6.20±0.79 | Un |
| 20 | Tacr1 |
4.44 | Up |
2.80±1.62 |
4.95±0.51 | 4 |
| 21 | Kcnv2 |
4.43 | Up |
4.63±1.57 |
6.78±0.35 | 1 |
| 22 | Cenpf |
4.40 | Up |
2.55±1.61 |
4.68±0.99 | 13 |
| 23 | Mt1a |
4.29 | Up |
9.24±0.57 |
11.34±0.55 | 19 |
| 24 | LOC287167 |
4.29 | Up |
5.25±1.57 |
7.35±0.86 | 10 |
| 25 | Tmem252 |
4.26 | Up |
5.34±1.72 |
7.43±0.26 | 1 |
| 26 | LOC100362110 |
4.17 | Up |
5.25±1.39 |
7.31±0.25 | 13 |
| 27 | LOC685488 |
4.12 | Up |
4.87±1.36 |
6.91±0.79 | 19 |
| 28 | Nr4a2 |
4.07 | Up |
7.36±0.95 |
9.39±1.07 | 3 |
| 29 | Irf7 | 12.31 | Down |
8.27±1.66 |
4.65±2.63 | 1 |
| 30 | RT1-A2 |
9.38 | Down |
10.74±1.30 |
7.51±2.07 | 20 |
| 31 | RT1-M6-1 |
7.73 | Down |
5.98±0.29 |
3.03±2.30 | 20 |
| 32 | A_44_P884971 |
6.99 | Down |
5.53±0.39 |
2.72±2.23 | 9 |
| 33 | Olr59 |
6.30 | Down |
5.11±0.49 |
2.46±2.07 | 1 |
| 34 | A_64_P033885 |
5.90 | Down |
5.63±1.04 |
3.07±1.92 | 18 |
| 35 | Enthd1 |
5.80 | Down |
4.83±0.71 |
2.30±1.98 | 7 |
| 36 | L2hgdh |
5.20 | Down |
6.05±0.50 |
3.67±1.22 | 6 |
| 37 | Osr1 |
4.95 | Down |
7.59±0.36 |
5.28±1.68 | 6 |
| 38 | Lrrc17 |
4.93 | Down |
5.20±0.79 |
2.90±1.67 | 4 |
| 39 | Tnfsf10 |
4.83 | Down |
6.66±1.00 |
4.39±0.69 | 2 |
| 40 | Adam33 |
4.46 | Down |
5.04±0.37 |
2.88±1.00 | 3 |
| 41 | Ltbp2 |
4.19 | Down |
5.32±0.87 |
3.25±0.74 | 6 |
| 42 | RGD1563091 |
4.03 | Down |
5.68±0.80 |
3.66±0.93 | 4 |
The microarray data were uploaded to the GEO
database (GEO no. GSE108342; https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE108342).
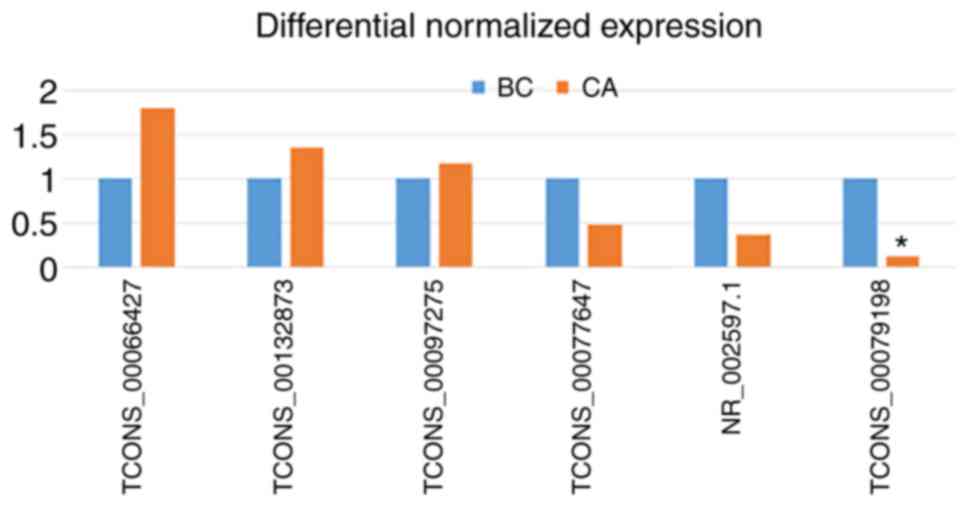
Validation of lncRNA microarray
To validate the lncRNA microarray results, three
upregulated and two downregulated lncRNAs were randomly selected
from the 58 differentially expressed lncRNAs and their expressions
were analyzed in five CA tissues and in four BC tissues using
RT-qPCR. The RT-qPCR results were consistent with the microarray
results, and RT-qPCR demonstrated that the TCONS_00079198 lncRNA
transcript was differentially expressed (P<0.05 using the 2-ΔΔCt
method; Fig. 2). RT-qPCR analysis
of the other lncRNAs transcripts demonstrated that the differential
expressions were consistent with the microarray data, but were not
significant (P>0.05).
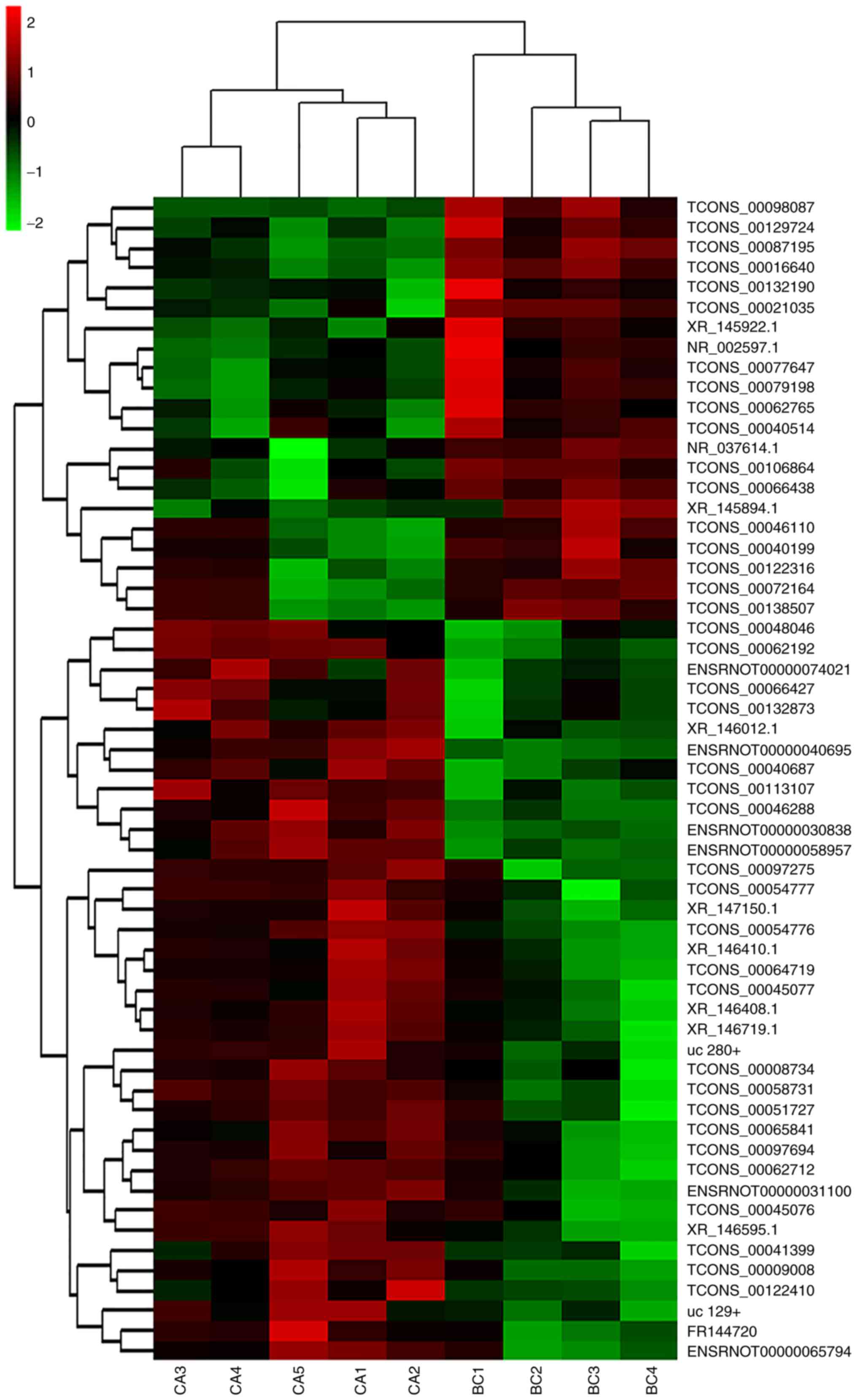
Hierarchical clustering analysis
Differentially expressed lncRNA data were used to
generate a heat map analysis of unsupervised hierarchical
clustering. The differentially expressed lncRNAs clearly segregated
into BC and CA clusters (Fig. 3)
indicating that there was a significant difference in the lncRNA
expression profiles between the CA and BC groups.
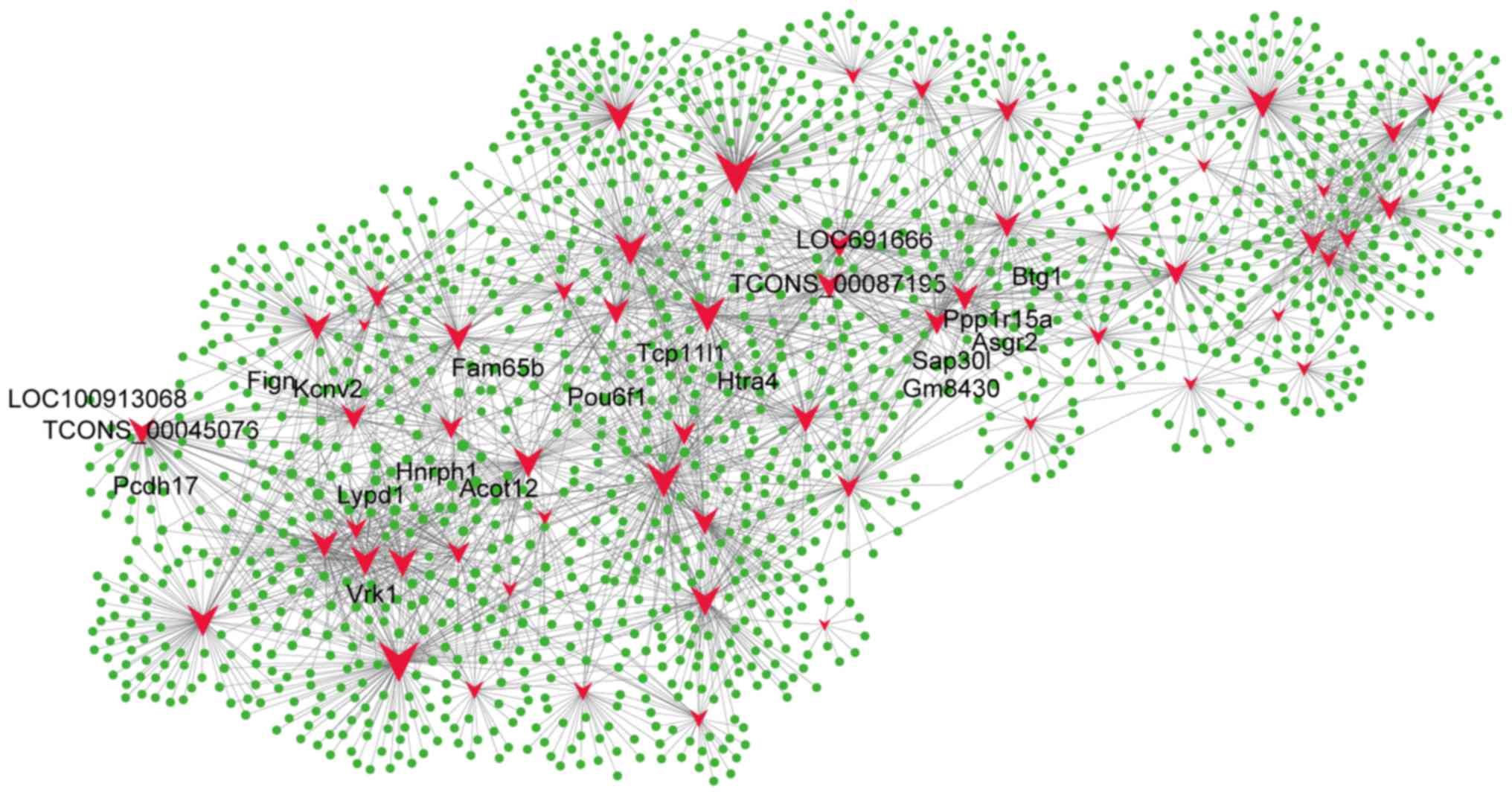
LncRNA and mRNA coexpression
profiles
To compare lncRNA profiles with their corresponding
mRNA network profiles, we calculated the Pearson correlation for
the expression value of each lncRNA with the expression value of
each mRNA for paired CA and BC samples (Fig. 4). Differentially expressed lncRNAs
were coexpressed with thousands of mRNAs. For instance,
TCONS_00087195 was coexpressed with 1,402 mRNA transcripts, and
TCONS_00045076 was coexpressed with 1,293 mRNA transcripts. The top
five mRNAs that correlated with TCONS_00087195 were Tcp11l1,
Ppp1r15a, Asgr2, Htra4, and LOC691666, and the top five mRNAs that
correlated with TCONS_00045076 were Lypd1, Fign, LOC100913068,
Kcnv2, and Pcdh17. Of the 58 lncRNAs, 40 are predicted to regulate
Btg1, 36 are predicted to regulate Acot12, 36 are predicted to
regulate Fam65b, 41 are predicted to regulate Gm8430, 38 are
predicted to regulate Hnrph1, 40 are predicted to regulate Pou6f1,
40 are predicted to regulate Sap30l, 36 are predicted to regulate
Vrk1, and 38 are predicted to regulate Zfp454 (Fig. 4). These data indicate that
differentially expressed lncRNAs potentially regulate mRNA
expression and form a complex lncRNA-mRNA interaction network
during CA.
Predictions of lncRNA functions during
CA pathology
We found that each differentially expressed lncRNA
associated with numerous differentially expressed mRNAs involved in
multiple processes, and our data indicated that these lncRNA-mRNA
networks play potentially important roles in pathological processes
during CA. Thus, we made functional predictions based on GO and
KEGG pathway annotations of differentially expressed lncRNAs and
data on the corresponding coexpressed mRNAs. The differentially
expressed lncRNAs could be grouped into numerous signaling pathways
and processes (Figs. 5 and
6). The relationships between
differentially expressed lncRNAs and their functional annotations
were predicted by the top 200 and top 500 GO pathway annotations
sorted by q-value, frequency counting, and statistical function
annotation according to component, function, and pathway process,
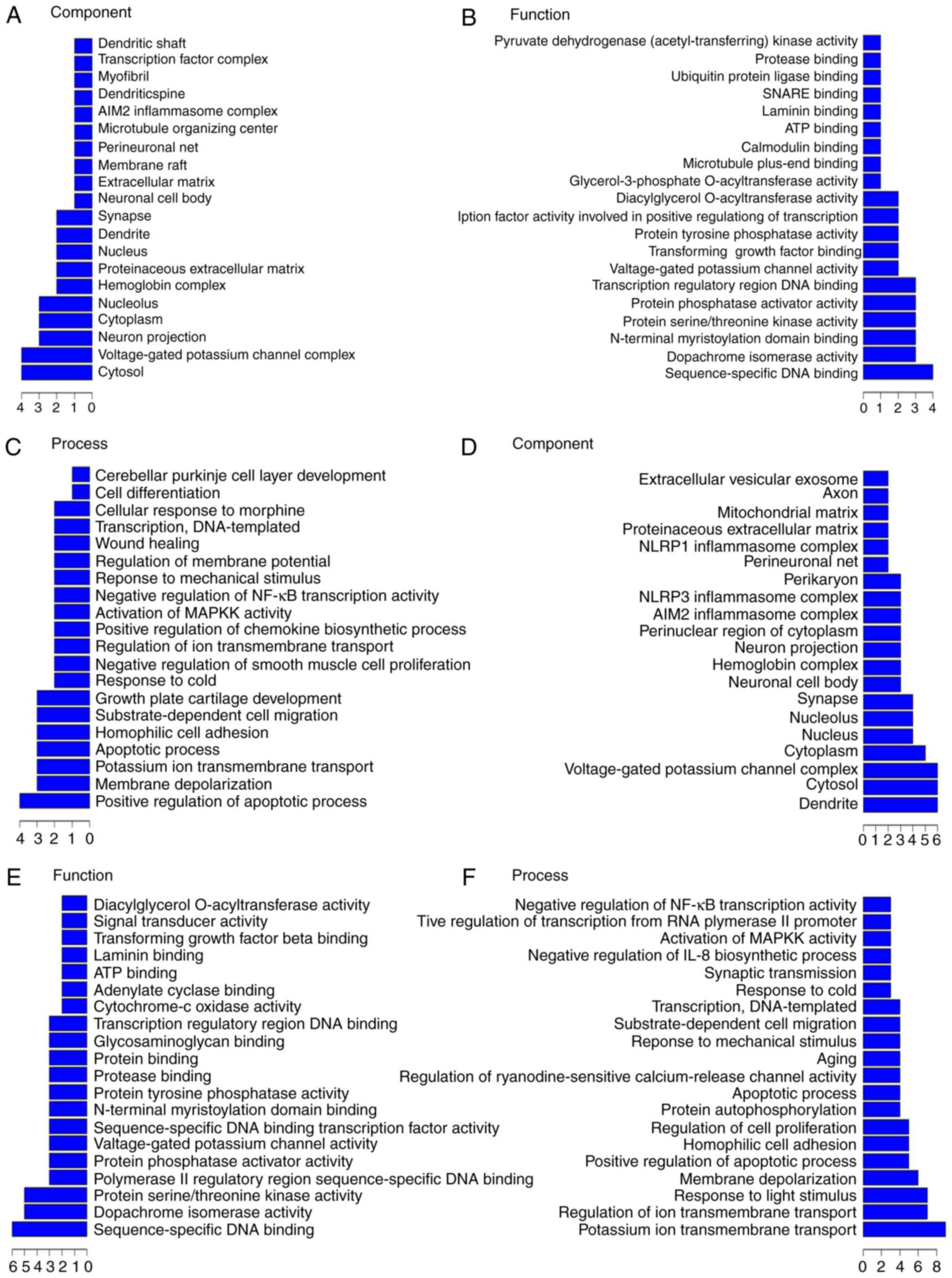
respectively. For example, in the top 200 GO pathway enrichments
(Fig. 5A), the most frequent
predictions were cytosol, voltage-gated potassium channel complex,
neuron projection, cytoplasm, and nucleolus with regards to the
pathways annotated by components. With regards to the pathways
annotated by function, the most frequent predictions were
sequence-specific DNA binding, dopachrome isomerase activity,
protein serine/threonine kinase activity, N-terminal myristoylation
domain binding, protein phosphatase activator activity, and
transcription regulatory region DNA binding. With regards to the
pathways annotated by process, the most frequent predictions were
positive regulation of apoptotic process, membrane depolarization,
potassium ion transmembrane transport, apoptotic process,
homophilic cell adhesion, substrate-dependent cell migration,
growth plate cartilage development, and response to cold. In the
top 500 GO pathway enrichments, (Fig.
5B), the most frequent component predictions were dendrite,
cytosol, voltage-gated potassium channel complex, cytoplasm,
nucleus, nucleolus, synapse, neuronal cell body, hemoglobin
complex, neuron projection, perinuclear region of cytoplasm, AIM2
inflammasome complex, NLRP3 inflammasome complex, and perikaryon.
The most frequent function predictions were sequence-specific DNA
binding, dopachrome isomerase activity, protein serine/threonine
kinase activity, protein phosphatase activator activity, and
voltage-gated potassium channel activity. The most frequent process
predictions were potassium ion transmembrane transport, regulation
of ion transmembrane transport, response to light stimulus,
membrane depolarization, positive regulation of apoptotic process,
homophilic cell adhesion, regulation of cell proliferation, protein
autophosphorylation, apoptotic process, positive regulation of
ryanodine-sensitive calcium-release channel activity, aging,
response to mechanical stimulus, substrate-dependent cell
migration, transcription, DNA-templated, and response to cold.
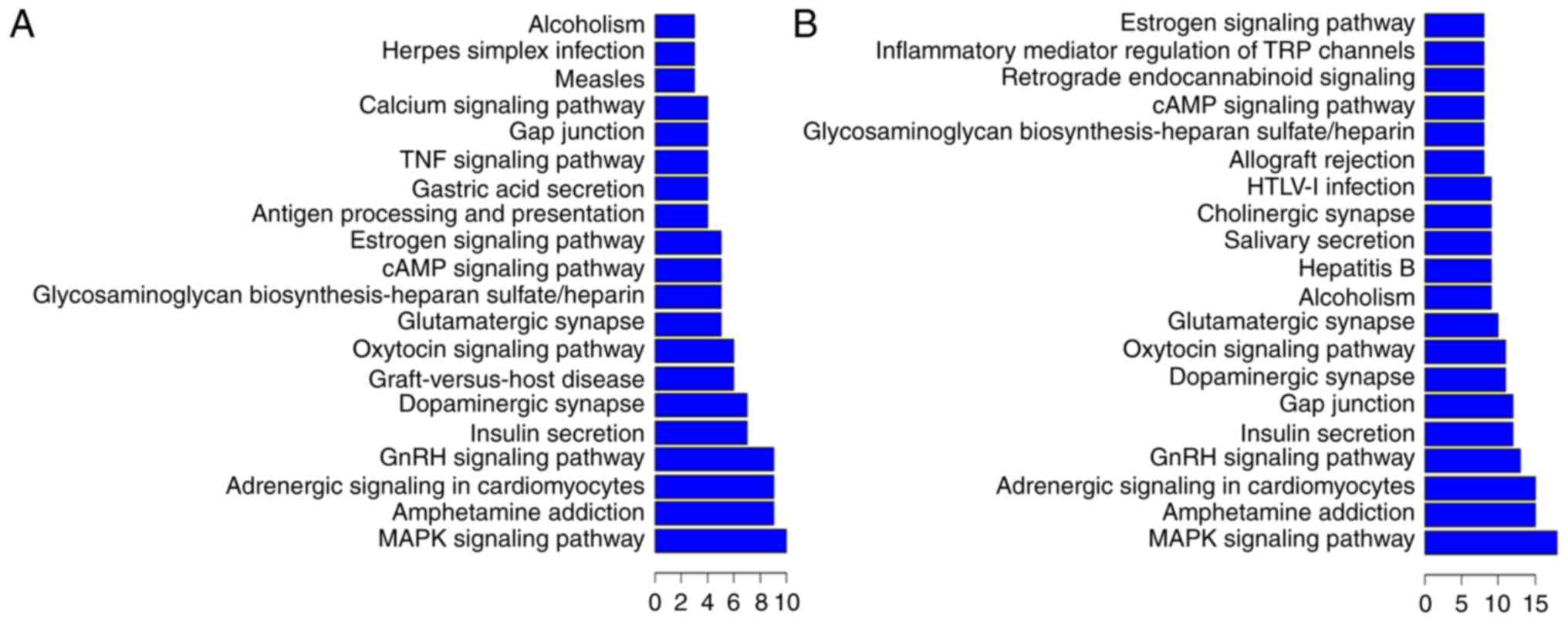
KEGG pathway annotations of differentially expressed
lncRNAs and their corresponding coexpressed mRNAs were performed to
predict lncRNA functions during CA pathology (Fig. 6). In the top 200 KEGG pathway
enrichments (Fig. 6A), the most
frequent predictions were the MAPK signaling pathway, amphetamine
addiction, adrenergic signaling in cardiomyocytes, the GnRH
signaling pathway, insulin secretion, dopaminergic synapse,
glutamatergic synapse, the cAMP signaling pathway, the estrogen
signaling pathway, antigen processing and presentation, gastric
acid secretion, the TNF signaling pathway, gap junction, and the
calcium signaling pathway. In the top 500 KEGG pathway enrichments
(Fig. 6B), the most frequent
predictions were the MAPK signaling pathway, amphetamine addiction,
adrenergic signaling in cardiomyocytes, the GnRH signaling pathway,
insulin secretion, gap junction, dopaminergic synapse, the oxytocin
signaling pathway, and the glutamatergic synapse. In summary, the
most common KEGG pathways and GO pathways involved in CA were
signaling pathways, apoptotic processes, inflammation, and synaptic
processes. For example, in the KEGG enrichment, for the most
downregulated lncRNA TCONS_00087195, the most frequently predicted
functions involved the Ras signaling pathway, amphetamine
addiction, and dopaminergic synapse. For the second-most
downregulated lncRNA TCONS_00129724, the most frequently predicted
functions involved the TNF signaling pathway, the NF-κ B signaling
pathway, and the Jak-STAT signaling pathway. For the third-most
downregulated lncRNA TCONS_00077647, the most frequently predicted
functions involved the RIG-I-like receptor signaling pathway, the
MAPK signaling pathway, and cytosolic DNA sensing. However, for the
most upregulated lncRNA TCONS_00045076, the most frequently
predicted function was protein processing in the dopaminergic and
glutamatergic synapse. For the second-most upregulated lncRNA
TCONS_00097275, the most frequently predicted functions involved
protein processing in the dopaminergic synapse, the estrogen
signaling pathway, and TNF signaling. For the third-most
upregulated lncRNA TCONS_00097694, the most predicted functions
involved the glutamatergic synapse, the dopaminergic synapse, and
cAMP signaling. As expected, one lncRNA can participate in multiple
KEGG pathways and both upregulated and downregulated lncRNAs could
be involved in some of the same processes, such as the TNF
signaling pathway and the dopaminergic synapse, indicating that
lncRNAs play complex roles in CA pathology.
Discussion
In the present study, we investigated lncRNA and
mRNA profiles in a CA-ROSC rat model during an early stage of
reperfusion using high-throughput lncRNA and mRNA microarrays. The
lncRNA microarray showed that 58 lncRNAs were differentially
expressed in the CA brain; 37 of the lncRNAs were upregulated and
21 of the lncRNAs were downregulated when compared with the control
samples. The mRNA microarray showed that 258 mRNA transcripts were
differentially expressed in the CA brain; 177 of the mRNAs were
upregulated and 81 of the mRNAs were downregulated. We investigated
the relationships between these lncRNAs and mRNAs and found that
each differentially expressed lncRNA was coexpressed with thousands
of mRNAs. Furthermore, we predicted the functions of these
differentially expressed lncRNAs with their corresponding
coexpressed mRNAs and found that hundreds of pathway annotations
were enriched in both GO and KEGG analyses. The predicted functions
of lncRNAs were primarily related to protein voltage-gated
potassium channel complexes, serine/threonine kinase activity,
protein phosphatase activator activity, the MAPK signaling pathway,
and apoptotic processes.
Previous studies have shown that ischemia leads to
extensive changes in cerebral lncRNA expression in rodents
(12,16), but the cerebral lncRNA expression
profile and the potential roles of lncRNAs during the early stages
of reperfusion remain unknown. The present study is the first that
synchronously examined genome-wide lncRNA and mRNA expression
patterns and their network profiles in a CA-ROSC model and that
discovered extensive alterations in expression of lncRNAs and their
corresponding mRNAs as a result of CA-ROSC.
In the present study, we found that CA-ROSC markedly
altered expression of mRNAs involved in vital metabolic pathways,
such as inflammatory and apoptotic pathways. In addition, we found
that lncRNAs coregulate expression of the differentially expressed
mRNAs. Some of the differentially mRNAs identified in our study
have also been identified in other ischemia studies (Table IV). To gain insight into the
potential roles of the differentially expressed lncRNAs and mRNAs,
we performed GO and KEGG pathway analyses to predict their
biological functions during CA-ROSC. We found that the
differentially expressed lncRNAs may be involved in protein
voltage-gated potassium channel complexes, serine/threonine kinase
activity, protein phosphatase activator activity, the MAPK
signaling pathway, and apoptotic processes. These proteins and
kinases are key enzymes in metabolic, inflammatory, and apoptotic
pathways, which have been shown to be involved in I/R injuries
(17–20). These results indicated that lncRNAs
in the cerebral region may participate in pathological processes
after CA-ROSC by promoting post-ischemic pathologies, including
ionic imbalance, receptor dysfunction, and the inflammatory
response during the early stages of reperfusion.
| Table IV.A brief summary of the literature on
differentially expressed mRNAs in cardiac arrest-return of
spontaneous circulation and ischemia. |
Table IV.
A brief summary of the literature on
differentially expressed mRNAs in cardiac arrest-return of
spontaneous circulation and ischemia.
| Author, year | mRNA | Animal | Literature
title | (Refs.) |
|---|
| Norman et
al, 2011 | Tumor necrosis
factor | Adult male C57BL/6
mice | Cardiopulmonary
arrest and resuscitation disrupts cholinergic anti-inflammatory
processes: A role for cholinergic α7 nicotinic receptors. | (35) |
| Hu et al,
2013 |
| Male Wistar rats
(15–16 months) | Ulinastatin
attenuates oxidation, inflammation and neural apoptosis in the
cerebral cortex of adult rats with ventricular fibrillation after
cardiopulmonary resuscitation. | (36) |
| Kaneko and
Kibayashi, 2012 |
| ddY mice and
Swiss-Webster mice | Mild hypothermia
facilitates the expression of cold-inducible RNA-binding protein
and heat shock protein 70.1 in mouse brain. | (37) |
| Anju et al,
2011 | Glutamate
decarboxylase mRNA | Wistar neonatal
rats (4-days) | Decreased GABAB
receptor function in the cerebellum and brain stem of hypoxic
neonatal rats: Role of glucose, oxygen and epinephrine
resuscitation. | (38) |
| Tyree et al,
2006 | Early Growth
Response mRNA | Newborn
piglets | Impact of room air
resuscitation on early growth response gene-1 in a neonatal piglet
model of cerebral hypoxic ischemia. | (39) |
| Vincze et
al, 2010 | Transforming growth
factors-β1, −2 and −3 | Rat | Distribution of
mRNAs encoding transforming growth factors-β1, −2, and −3 in the
intact rat brain and after experimentally induced focal
ischemia. | (40) |
| Zhu et al,
2001 | Transforming growth
factor-β1 | Rat | β2-adrenoceptor
stimulation enhances latent transforming growth factor-β-binding
protein-1 and transforming growth factor-β1 expression in rat
hippocampus after transient forebrain ischemia. | (41) |
For the downregulated lncRNAs, the most frequent
predicted functions enriched in the KEGG analysis involved the Ras
signaling pathway, amphetamine addiction, the NF-κ B signaling
pathway, the Jak-STAT signaling pathway, the RIG-I-like receptor
signaling pathway, the MAPK signaling pathway, and the cytosolic
DNA sensing pathway. Previous studies have shown that some of these
pathways, such as the Ras signaling pathway (21), the NF-κ B signaling pathway
(22,23), the Jak-STAT signaling pathway
(24), the RIG-I-like receptor
signaling pathway (25), the
cytosolic DNA sensing pathway (25), and the MAPK signaling pathway
(26,27) are involved in CA-ROSC and I/R
injury. In addition, the cocaine- and amphetamine-regulated
transcript (CART), which codes for a neuropeptide involved in
amphetamine addiction has also been shown to play a neuroprotective
role in cerebral ischemia and reperfusion (I/R) injury (28,29).
The most frequent predicted functions of the
upregulated lncRNAs involve protein processing during glutamatergic
synapsis (30), the estrogen
signaling pathway (26), and the
cAMP signaling pathway (31,32),
which play vital roles in cerebral ischemia and reperfusion injury.
In summary, our data indicated that the lncRNA-mRNA network
interactions have potentially complex roles in cerebral I/R injury.
Both upregulated and downregulated lncRNAs were predicted to
participate in pathological processes during cerebral I/R injury,
such as dopaminergic synapsis and the TNF signaling pathway
(33,34), indicating that lncRNAs work
synergistically during I/R injury.
There were several limitations in our study. First,
we only investigated differentially expressed lncRNAs and mRNAs in
cerebral cortex tissue. Second, the time for CA sample collection
lasted only eight min. However, the aberrant expressions of lncRNAs
and mRNAs after CA-ROSC may have spatial and temporal patterns that
we did not evaluate in this study. The samples used in this study
were limited (five samples in the CA group and four samples in the
BC group). More samples and future studies are required to verify
these results.
In this study, we found that CA-ROSC led to
extensive alterations in lncRNA and mRNA expression patterns, and
these alterations may affect the mRNA transcription and protein
translation of vital pathways during I/R injury. Thus, lncRNAs may
be promising novel targets for the development of therapeutics to
reduce cerebral damage. Future studies should determine whether
modulating specific lncRNAs can prevent post-CA pathophysiological
events and/or promote plasticity and regeneration.
Acknowledgements
Not applicable.
Funding
The present study was supported by funding from The
National Nature Science Foundation of China (grant nos. 81272021
and 81571867), The Science and Technology Foundation of Guangdong
Province, China (grant nos. 2012B061700046 and 2012B031800286), and
The Research Program for Colleges and Universities in Guangzhou
(grant no. 2012C054).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available in the Genome Expression Omnibus (GEO)
repository, www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE108342.
Authors' contributions
CH and ZL conceived the idea for the study. CH, RL,
MY, XZ and XLi contributed to the design, and performed the animal
experiments and harvested samples. XLia and HW contributed to
sample measurement procedures. ZZ and QL performed data analysis.
CH, MY and RL wrote the manuscript, and CH and XZ revised the
manuscript.
Ethics approval and consent to
participate
The animal studies were approved by the
Institutional Animal Care and Use Committee of Sun Yat-sen
University (Guangdong, China) and the procedures used were in
accordance with the Animal Research Reporting for In Vivo
Experiments guidelines on animal research.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Writing Group Members, Mozaffarian D,
Benjamin EJ, Go AS, Arnett DK, Blaha MJ, Cushman M, Das SR, de
Ferranti S, Després JP, et al: Executive summary: Heart disease and
stroke statistics-2016 update: A report from the american heart
association. Circulation. 133:447–454. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Neumar RW, Nolan JP, Adrie C, Aibiki M,
Berg RA, Böttiger BW, Callaway C, Clark RS, Geocadin RG, Jauch EC,
et al: Post-cardiac arrest syndrome: Epidemiology, pathophysiology,
treatment, and prognostication. A consensus statement from the
International Liaison Committee on Resuscitation (American Heart
Association, Australian and New Zealand Council on Resuscitation,
European Resuscitation Council, Heart and Stroke Foundation of
Canada, InterAmerican heart foundation, Resuscitation Council of
Asia, and the Resuscitation Council of Southern Africa); the
American Heart Association Emergency Cardiovascular Care Committee;
the Council on Cardiovascular Surgery and Anesthesia; the Council
on Cardiopulmonary, Perioperative, and Critical care; the Council
on Clinical Cardiology; and the Stroke Council. Circulation.
118:2452–2483. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Khalil AM, Guttman M, Huarte M, Garber M,
Raj A, Rivea Morales D, Thomas K, Presser A, Bernstein BE, van
Oudenaarden A, et al: Many human large intergenic noncoding RNAs
associate with chromatin-modifying complexes and affect gene
expression. Proc Natl Acad Sci USA. 106:pp. 11667–11672. 2009;
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cabili MN, Trapnell C, Goff L, Koziol M,
Tazon-Vega B, Regev A and Rinn JL: Integrative annotation of human
large intergenic noncoding RNAs reveals global properties and
specific subclasses. Genes Dev. 25:1915–1927. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leung A, Trac C, Jin W, Lanting L, Akbany
A, Sætrom P, Schones DE and Natarajan R: Novel long noncoding RNAs
are regulated by angiotensin II in vascular smooth muscle cells.
Circ Res. 113:266–278. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bonasio R and Shiekhattar R: Regulation of
transcription by long noncoding RNAs. Annu Rev Genet. 48:433–455.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dong X, Yu LG, Sun R, Cheng YN, Cao H,
Yang KM, Dong YN, Wu Y and Guo XL: Inhibition of PTEN expression
and activity by angiotensin II induces proliferation and migration
of vascular smooth muscle cells. J Cell Biochem. 114:174–182. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qureshi IA and Mehler MF: Long non-coding
RNAs: Novel targets for nervous system disease diagnosis and
therapy. Neurotherapeutics. 10:632–646. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yin KJ, Hamblin M and Chen YE: Non-coding
RNAs in cerebral endothelial pathophysiology: Emerging roles in
stroke. Neurochem Int. 77:9–16. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Antoniou D, Stergiopoulos A and Politis
PK: Recent advances in the involvement of long non-coding RNAs in
neural stem cell biology and brain pathophysiology. Front Physiol.
5:1552014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dharap A, Pokrzywa C and Vemuganti R:
Increased binding of stroke-induced long non-coding RNAs to the
transcriptional corepressors Sin3A and coREST. ASN Neuro.
5:283–289. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kilkenny C, Browne WJ, Cuthill IC, Emerson
M and Altman DG: Improving bioscience research reporting: The
ARRIVE guidelines for reporting animal research. Osteoarthritis
Cartilage. 20:256–260. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin JY, Liao XX, Li H, Wei HY, Liu R, Hu
CL, Huang GQ, Dai G and Li X: Model of cardiac arrest in rats by
transcutaneous electrical epicardium stimulation. Resuscitation.
81:1197–1204. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chun-Lin H, Jie W, Xiao-Xing L, Xing L,
Yu-Jie L, Hong Z, Xiao-Li J and Gui-Fu W: Effects of therapeutic
hypothermia on coagulopathy and microcirculation after
cardiopulmonary resuscitation in rabbits. Am J Emerg Med.
29:1103–1110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dharap A, Nakka VP and Vemuganti R: Effect
of focal ischemia on long noncoding RNAs. Stroke. 43:2800–2802.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xiang Y, Zhao H, Wang J, Zhang L, Liu A
and Chen Y: Inflammatory mechanisms involved in brain injury
following cardiac arrest and cardiopulmonary resuscitation. Biomed
Rep. 5:11–17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Uchino H, Ogihara Y, Fukui H, Chijiiwa M,
Sekine S, Hara N and Elmér E: Brain injury following cardiac
arrest: Pathophysiology for neurocritical care. J Intensive Care.
4:312016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Quillinan N, Herson PS and Traystman RJ:
Neuropathophysiology of brain injury. Anesthesiol Clin. 34:453–464.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Quillinan N, Grewal H, Deng G, Shimizu K,
Yonchek JC, Strnad F, Traystman RJ and Herson PS: Region-specific
role for GluN2B-containing NMDA receptors in injury to Purkinje
cells and CA1 neurons following global cerebral ischemia.
Neuroscience. 284:555–565. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yao C, Zhang J, Chen F and Lin Y:
Neuroprotectin D1 attenuates brain damage induced by transient
middle cerebral artery occlusion in rats through TRPC6/CREB
pathways. Mol Med Rep. 8:543–550. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei X, Zhang B, Zhang Y, Li H, Cheng L,
Zhao X, Yin J and Wang G: Hydrogen sulfide inhalation improves
neurological outcome via NF-κB-mediated inflammatory pathway in a
rat model of cardiac arrest and resuscitation. Cell Physiol
Biochem. 36:1527–1538. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tang ZX, Chen GX, Liang MY, Rong J, Yao
JP, Yang X and Wu ZK: Selective antegrade cerebral perfusion
attenuating the TLR4/NF-κB pathway during deep hypothermia
circulatory arrest in a pig model. Cardiology. 128:243–250. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ottani A, Neri L, Canalini F, Calevro A,
Rossi R, Cappelli G, Ballestri M, Giuliani D and Guarini S:
Protective effects of the melanocortin analog NDP-α-MSH in rats
undergoing cardiac arrest. Eur J Pharmacol. 745:108–116. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang H, Wang G, Zhang L, Zhang J, Zhang J,
Wang Q and Billiar TR: ADAR1 suppresses the activation of cytosolic
RNA-sensing signaling pathways to protect the liver from
ischemia/reperfusion injury. Sci Rep. 6:202482016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lebesgue D, Chevaleyre V, Zukin RS and
Etgen AM: Estradiol rescues neurons from global ischemia-induced
cell death: Multiple cellular pathways of neuroprotection.
Steroids. 74:555–561. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jover-Mengual T, Zukin RS and Etgen AM:
MAPK signaling is critical to estradiol protection of CA1 neurons
in global ischemia. Endocrinology. 148:1131–1143. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Qiu B, Liu J, Zhu WG and Zhu S:
Cocaine- and amphetamine-regulated transcript facilitates the
neurite outgrowth in cortical neurons after oxygen and glucose
deprivation through PTN-dependent pathway. Neuroscience.
277:103–110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bin J, Wang Q, Zhuo YY, Xu JP and Zhang
HT: Piperphentonamine (PPTA) attenuated cerebral ischemia-induced
memory deficits via neuroprotection associated with anti-apoptotic
activity. Metab Brain Dis. 27:495–505. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tjepkema-Cloostermans MC, Hindriks R,
Hofmeijer J and van Putten MJ: Generalized periodic discharges
after acute cerebral ischemia: Reflection of selective synaptic
failure? Clin Neurophysiol. 125:255–262. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Steinberg SF, Alcott S, Pak E, Hu D,
Protas L, Möise NS, Robinson RB and Rosen MR: beta(1)-Receptors
increase cAMP and induce abnormal Ca(i) cycling in the German
shepherd sudden death model. Am J Physiol Heart Circ Physiol.
282:H1181–H1188. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li P, Gu T, Wang C, Zhang G and Shi E:
Neuregulin 1 attenuates neuronal apoptosis induced by deep
hypothermic circulatory arrest through ErbB4 signaling in rats. J
Cardiovasc Pharmacol. 66:551–557. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang W, Zhao L, Bai F, Zhang T, Dong H and
Liu L: The protective effect of dopamine against OGD/R
injury-induced cell death in HT22 mouse hippocampal cells. Environ
Toxicol Pharmacol. 42:176–182. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sui B, Li Y and Ma L: Postconditioning
improvement effects of ulinastatin on brain injury following
cardiopulmonary resuscitation. Exp Ther Med. 8:1301–1307. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Norman GJ, Morris JS, Karelina K, Weil ZM,
Zhang N, Al-Abed Y, Brothers HM, Wenk GL, Pavlov VA, Tracey KJ and
Devries AC: Cardiopulmonary arrest and resuscitation disrupts
cholinergic anti-inflammatory processes: A role for cholinergic α7
nicotinic receptors. J Neurosci. 31:3446–3452. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hu CL, Xia JM, Cai J, Li X, Liao XX, Li H,
Zhan H, Dai G and Jing XL: Ulinastatin attenuates oxidation,
inflammation and neural apoptosis in the cerebral cortex of adult
rats with ventricular fibrillation after cardiopulmonary
resuscitation. Clinics (Sao Paulo). 68:1231–1238. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kaneko T and Kibayashi K: Mild hypothermia
facilitates the expression of cold-inducible RNA-binding protein
and heat shock protein 70.1 in mouse brain. Brain Res.
1466:128–136. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Anju TR, Jayanarayanan S and Paulose CS:
Decreased GABAB receptor function in the cerebellum and brain stem
of hypoxic neonatal rats: Role of glucose, oxygen and epinephrine
resuscitation. J Biomed Sci. 18:312011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tyree MM, Dalgard C and O'Neill JT: Impact
of room air resuscitation on early growth response gene-1 in a
neonatal piglet model of cerebral hypoxic ischemia. Pediatric Res.
59:423–427. 2006. View Article : Google Scholar
|
|
40
|
Vincze C, Pál G, Wappler EA, Szabó ER,
Nagy ZG, Lovas G and Dobolyi A: Distribution of mRNAs encoding
transforming growth factors-beta1, −2, and −3 in the intact rat
brain and after experimentally induced focal ischemia. J Comp
Neurol. 518:3752–3770. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhu Y, Culmsee C, Roth-Eichhorn S and
Krieglstein J: Beta(2)-adrenoceptor stimulation enhances latent
transforming growth factor-beta-binding protein-1 and transforming
growth factor-beta1 expression in rat hippocampus after transient
forebrain ischemia. Neuroscience. 107:593–602. 2001. View Article : Google Scholar : PubMed/NCBI
|