Introduction
Glutamate, the principal excitatory neurotransmitter
in the brain, is stored in the vesicles of the presynaptic terminal
via the membrane-bound vesicular glutamate transporter (VGLUT)
(1). Glutamate is released into
the synaptic cleft following the fusion of a vesicle with the cell
membrane (2); once vesicles are
bound to cell membrane VGLUTs, they are emptied of glutamate, which
is released into the cell (3).
Therefore, the expression level of VGLUTs reflects the amount of
glutamate in vesicles (4,5). Three isoforms of VGLUTs have been
isolated to date, with VGLUT1 and VGLUT2 being expressed primarily
in the hippocampus and cerebral cortex (6). Knockdown of VGLUT1 in mice has
resulted in depression-like behavior and impaired long-term
recognition memory; however, short-term memory was retained in the
novel object recognition test, as well as spatial memory as
observed in the Morris water maze test (7). Knockout of VGLUT2 attenuated spatial
learning and memory as well as synaptic plasticity in the mouse
hippocampus (8).
Glutamate levels, as well as VGLUT1 and VGLUT2
expression, are known to be altered during the aging process
(9–13), as well as in Alzheimer's disease
(14,15). However, there have been conflicting
results regarding the alterations in VGLUT1 and VGLUT2 in the aged
hippocampus. For example, VGLUT1 levels are relatively resistant to
aging processes, whereas VGLUT2 are more susceptible to the aging
process. In an aging model using senescence-accelerated mice
(SAMP8), VGLUT1 expression was maintained at several ages compared
with age-matched control groups (13). However, in the normal aging model
using Wistar rats, VGLUT1 expression was decreased in the
hippocampus from PM18 (9). By
contrast, VGLUT2 decreased significantly at PM12 in Wistar rats
(9) and SAMP8 (13).
Mongolian gerbils are ideal experimental models for
studies on aging and epilepsy as they have relatively short
lifespans, a homogenous genetic background and are easy to perform
behavioral tests on (16,17). The average lifespan of half the
population was 110 and 139 weeks in male and female gerbils,
respectively (18). Therefore, in
the present study, the changes in hippocampal VGLUT1 and VGLUT2
immunoreactivity and protein levels were investigated in Mongolian
gerbils of different ages to estimate the changes of glutamate
transporting systems in the aged gerbils and to elucidate the
association between VGLUT and aging processes in the
hippocampus.
Materials and methods
Experimental animals
Male Mongolian gerbils (n=5-; 3-month-old; 50–60 g)
were purchased from Japan SLC, Inc. (Shizuoka, Japan). They were
housed under standard conditions at a constant temperature (22°C)
and humidity (60%), with a 12-h light/dark cycle, and free access
to food and water. The handling and care of the animals conformed
to guidelines compliant with the current international laws and
policies [National Institutes of Health (NIH) Guide for the Care
and Use of Laboratory Animals; NIH Publication no. 85-23, 1985,
revised 1996] (19). Ethical
approval was obtained from the Institutional Animal Care and Use
Committee of Kangwon National University (Chuncheon, Gangwon,
Republic of Korea) for all animal procedures in the present study
(no. KW-160802-2). Animals were firstly divided into 3 groups, and
were then further divided into 5 subgroups (n=10 in each subgroup):
Adolescent [postnatal month (PM) 1], adult (PM6 and PM12) and aged
(PM18 and PM24) groups. All experiments were conducted with an
effort to minimize the number of animals required and the suffering
caused by the procedures employed in the present study.
Tissue processing
For histology experiments, the animals (n=5/group)
at PM1, 6, 12, 18 and 24 were anesthetized with 1 g/kg urethane by
intraperitoneal injection (Sigma-Aldrich; Merck KGaA), then
perfused transcardially with 0.1 M of phosphate-buffered saline
(PBS; pH 7.4) followed by 4% paraformaldehyde in 0.1 M PBS (pH
7.4). Brains were then removed and postfixed in the same fixative
for 12 h at 25°C prior to cryopreservation via overnight storage in
30% sucrose at 4°C. Serial coronal brain sections (30 µm) were
generated using a cryostat (Leica Microsystems GmbH, Wetzlar,
Germany) at −25°C and maintained in 6-well plates containing PBS
until further processing.
Immunohistochemistry
To ensure that the immunohistochemical data were
comparable between groups, sections were carefully processed under
parallel conditions. Tissue sections located 90 µm apart from each
other were selected from within an area between 1.4–2.0 mm
posterior to the bregma, as defined by a gerbil atlas (20). Five sections, (30 µm) from each of
the tissue sections collected, located 90 µm apart from each other
were sequentially incubated with 0.3% hydrogen peroxide
(H2O2) in PBS for 30 min and 10% normal goat
serum (cat no. S1000; Vector Laboratories, Inc., Burlingame, CA,
USA) in 0.05 M PBS for 30 min at 25°C. Sections were then incubated
with the guinea pig anti-VGLUT1 antibody (dilution 1:5,000; cat no.
AB5905; Chemicon, Temecula, CA, USA) and guinea pig anti-VGLUT2
(dilution 1:10,000; cat no. AB2251; Chemicon) overnight at room
temperature. Sections were then incubated with a biotinylated goat
anti-guinea pig IgG (1:200; cat no. BA-7000; Vector Laboratories,
Inc.) for 2 h at room temperature, followed by a
streptavidin-peroxidase complex (ABC kit; cat no. PK-6100; Vector
Laboratories, Inc.) for 1 h at room temperature. Immunostaining was
visualized via detection with DAB in 0.1 M Tris-HCl buffer (pH
7.2). Sections were then dehydrated in graded ethanol (70, 80, 90,
95, 100, 100 or 70–100%) and mounted on gelatin-coated slides in
Canada balsam (Kanto Chemical Co., Inc., Tokyo, Japan).
In order to establish the specificity of the VGLUT1
and VGLUT2 antibodies, the procedure included the omission of the
VGLUT1 and VGLUT2 antibodies, goat anti-guinea pig IgG, and the
substitution of normal goat serum for the primary antibody. In
addition, for positive control test, immunohistochemistry was
conducted with VGLUT1 and VGLUT2 antibodies in the cerebellum of
gerbils as VGLUT1 and VGLUT2 are specifically detected in
cerebellum (21).
Analysis of the hippocampal CA1 and CA2/3 regions,
and dentate gyrus was performed using an ImageJ software v.1.5
(NIH, Bethesda, MD, USA). Digital images of the mid-point of each
region were captured using a BX51 light microscope (Olympus
Corporation, Tokyo, Japan) equipped with a digital camera (DP72;
Olympus Corporation) connected to a computer monitor. Images were
calibrated into an array of 512×512 pixels corresponding to a
tissue area of 1,200×900 µm (magnification, ×100). Each pixel
resolution was 256 gray levels, and the area was divided into the
strata oriens, pyramidale and radiatum. The intensity of VGLUT1 and
VGLUT2 immunoreactivity was evaluated by relative optical density
(ROD), which was obtained following transformation of the mean gray
level using the formula: ROD=log (256/mean gray level). The ROD of
background staining was determined in unlabeled portions of the
sections using Photoshop CC 2015 software (Adobe Systems, Inc., San
Jose, CA, USA); this value was subtracted to correct for
nonspecific staining, using ImageJ v.1.50 software (NIH). Data are
expressed as a percentage of the PM1 group values (set to
100%).
Western blot analysis
To confirm the age-associated changes of VGLUT1 and
VGLUT2 in the hippocampus, animals at PM1, 6, 12, 18, and 24
(n=5/subgroup) were sacrificed and used for western blot analysis.
Following euthanasia and the removal of brains, tissues were cut to
500 µm thick sections using a vibratome (Leica Microsystems GmbH)
and the hippocampus was dissected out the using a surgical blade.
Hippocampal tissues were homogenized in 50 mM PBS (pH 7.4) at 4°C
for 30 sec containing 0.1 mM ethylene glycol bis (2-aminoethyl
ether)-N,N,N,N tetraacetic acid (pH 8.0), 0.2% nonidet P-40, 10 mM
ethylenediamine tetraacetic acid (pH 8.0), 15 mM sodium
pyrophosphate, 100 mM β-glycerophosphate, 50 mM NaF, 150 mM NaCl, 2
mM sodium orthovanadate, 1 mM phenylmethylsulfonyl fluoride and 1
mM dithiothreitol (DTT). Following centrifugation for 5 min at
16,000 × g at 4°C, the protein level was determined in the
supernatants using a Micro Bicinchoninic Acid protein assay kit
with bovine serum albumin as the standard according to the
manufacturer's instructions (Pierce; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). Aliquots containing 20 µg of total protein
were boiled in loading buffer containing 150 mM Tris (pH 6.8), 3 mM
DTT, 6% SDS, 0.3% bromophenol blue and 30% glycerol. Then, each
aliquot was loaded onto a 12% polyacrylamide gel. Following
electrophoresis, proteins were transferred to nitrocellulose
membranes (Pall Corporation, East Hills, NY, USA). The membranes
were then incubated with 5% non-fat dry milk in PBS containing 0.1%
Tween-20 for 45 min at 25°C, followed by incubation with guinea pig
anti-VGLUT1 (dilution 1:10,000; cat no. AB5905; EMD Millipore,
Billerica, MA, USA) and guinea pig anti-VGUT2 (dilution 1:20,000;
cat no. AB2251; EMD Millipore) for 12 h at 4°C. Then, the membrane
was incubated with peroxidase-conjugated anti-guinea pig antibody
(1:400; cat no. BA-7000; Vector Laboratories, Inc.). Visualization
was performed using an enhanced luminol-based chemiluminescent kit
(Pierce; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The blots were scanned and densitometry
was performed for the quantification of ROD of each band using
Scion Image software (version 4.0.3; Scion Corp., Frederick, MD,
USA). These data were normalized against β-actin (dilution 1:500;
cat no. ab8229; Abcam, Cambridge, UK).
Statistical analysis
The data are presented as the mean ± standard error
mean of triplicate measurement. Differences among the groups were
statistically analyzed by one-way analysis of variance followed by
a Bonferroni's post hoc test, using GraphPad Prism v5.01 software
(GraphPad Software, Inc., La Jolla, CA). P<0.05 was considered
to indicate a statistically significant difference.
Results
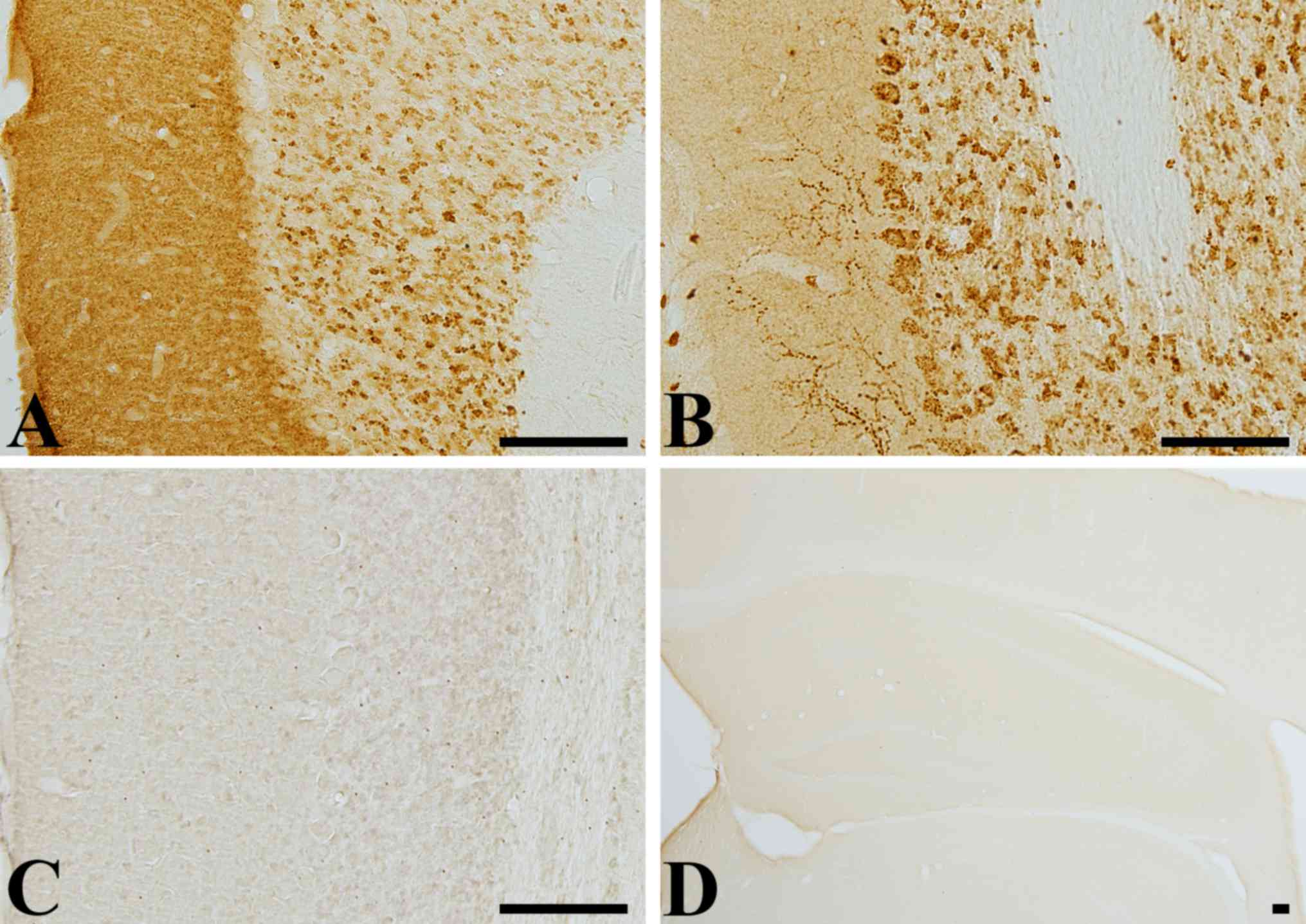
Antibody specificity
VGLUT1 immunoreactivity was identified in the
molecular and granular layer of cerebellum, while VGLUT2
immunoreactivity was mainly detected in the Purkinje cell and
granular layer of cerebellum. In the molecular layer, VGLUT1
immunoreactivity was diffusely observed, while VGLUT2
immunoreactivity was found in some coarse fibers (Fig. 1A and B). Negative control test with
pre-immune serum did not show any marked staining of VGLUT1 and
VGLUT2 in the cerebellum (Fig. 1C and
D).
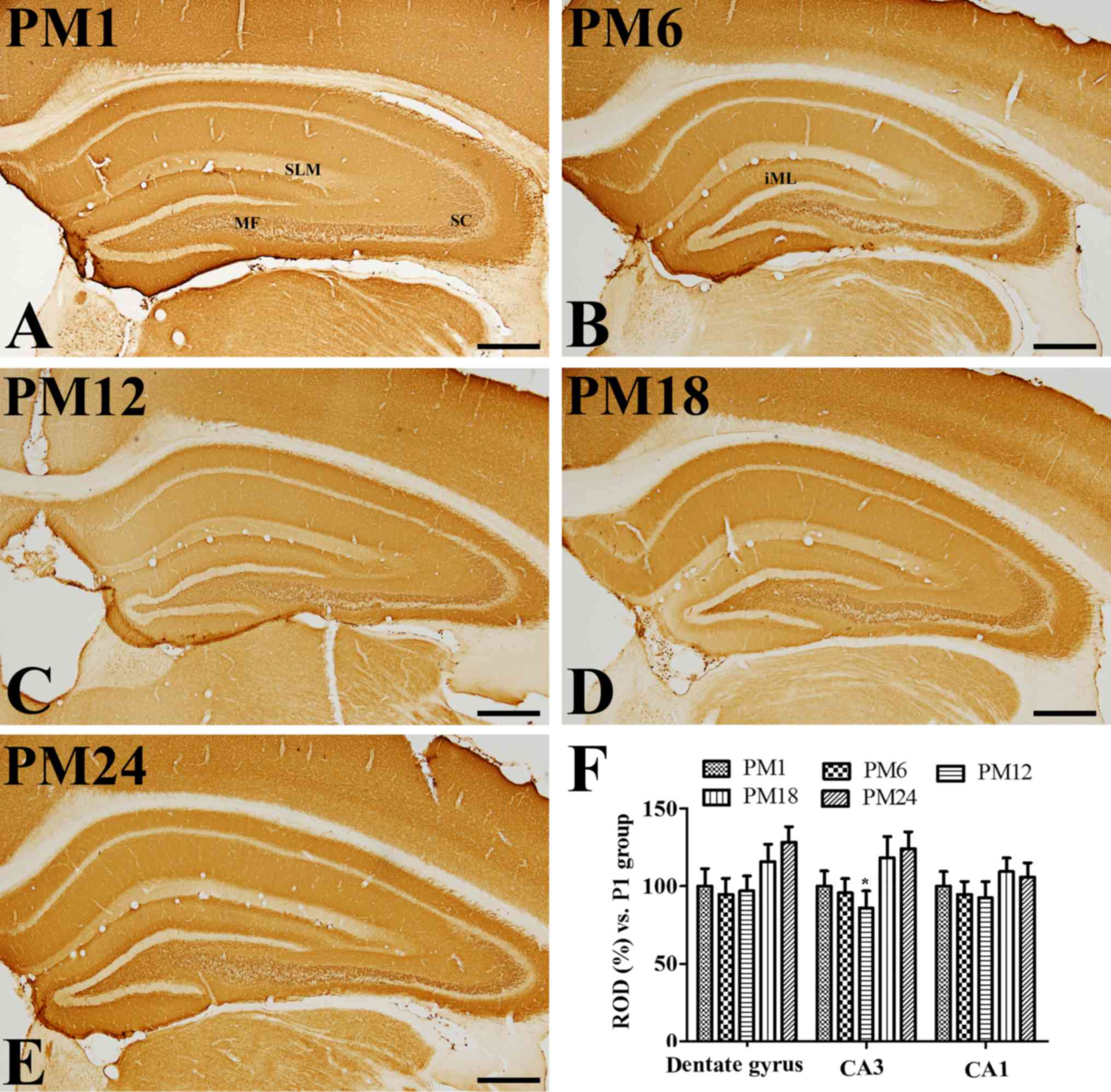
Age-associated changes in VGLUT1
immunoreactivity
In all groups, VGLUT1 immunoreactivity was observed
in the mossy fibers of the dentate gyrus, as well as the Schaffer
collaterals and stratum lacunosum-moleculare in the hippocampal CA1
region (Fig. 2A-E). There were
some variations in the VGLUT1 expression pattern in the inner
molecular layer of the dentate gyrus; VGLUT1 immunoreactivity was
clearly visible in the PM6 and PM24 groups (Fig. 2B and E). VGLUT1 immunoreactivity
was altered in the dentate gyrus, however, not in the hippocampal
CA1-3 regions, with increasing age. VGLUT1 immunoreactivity was
markedly increased in the dentate gyrus of the PM18 and PM24 groups
when compared with the PM1 group (Fig.
2F).
Age-associated changes in VGLUT2
immunoreactivity
In all groups, VGLUT2 immunoreactivity was observed
in the granule cell layer and outer molecular layer of the dentate
gyrus. In addition, VGLUT2 immunoreactivity was also detected in
the stratum pyramidale, Schaffer collaterals and stratum
lacunosum-moleculare in the hippocampal CA1-3 (Fig. 3A-E). VGLUT2 immunoreactivity
gradually decreased in the dentate gyrus with age; however, VGLUT2
immunoreactivity in the PM18 group increased significantly in the
CA1-3 regions when compared with the PM1 and PM12 groups (Fig. 3F). In addition, VGLUT2
immunoreactivity was also markedly detected in the non-pyramidal
cells of the stratum oriens and radiatum of the CA1-3 regions
(Fig. 3D). However, VGLUT2
immunoreactivity in the PM24 group was significantly decreased in
the hippocampal CA1-3 regions when compared with the PM1 group
(Fig. 3E and F).
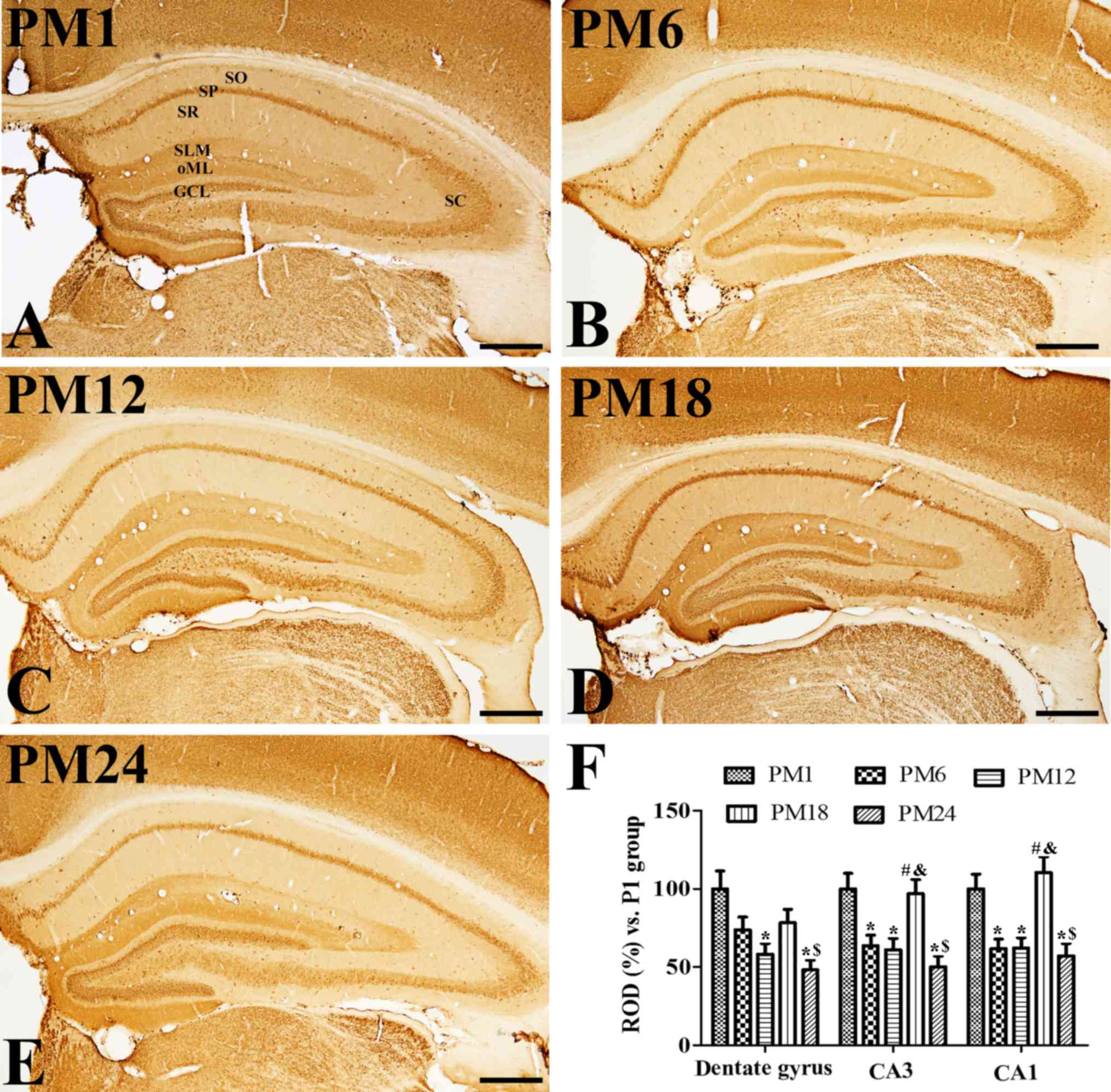 | Figure 3.Immunohistochemistry for VGLUT2 in the
hippocampus of the (A) adolescent PM1, adult (B) PM6 and (C) PM12,
and aged (D) PM18 and (E) PM24 groups. VGLUT2 immunoreactivity was
observed in the granule cell layer (indicated by GCL) and the outer
molecular layer (indicated by oML), stratum pyramidale (indicated
by SP), Schaffer collaterals (indicated by SC) and stratum
lacunosum-moleculare (indicated by SLM) of the hippocampus. VGLUT2
immunoreactivity as markedly increased in the stratum radiatum
(indicated by SR) and oriens (indicated by SO) of the CA1 region in
the PM18 group. VGLUT2 immunoreactivity decreased in all layers of
the hippocampus in the PM24 group. Scale bar=500 µm. (F) ROD are
expressed as a percentage of the value of the VGLUT2
immunoreactivity in the PM1 group in the dentate gyrus, hippocampal
CA1 and CA2/3 regions per section of the PM1, PM6, PM12, PM18 and
PM24 groups (n=5/group). Data are presented as the mean ± standard
error mean. *P<0.05 vs. the PM1 group; #P<0.05,
vs. the PM6 group; $P<0.05 vs. the PM12 group;
&P<0.05 vs. the PM18 group. VGLUT, vesicular
glutamate transporter; PM, postnatal month; ROD, relative optical
densities. |
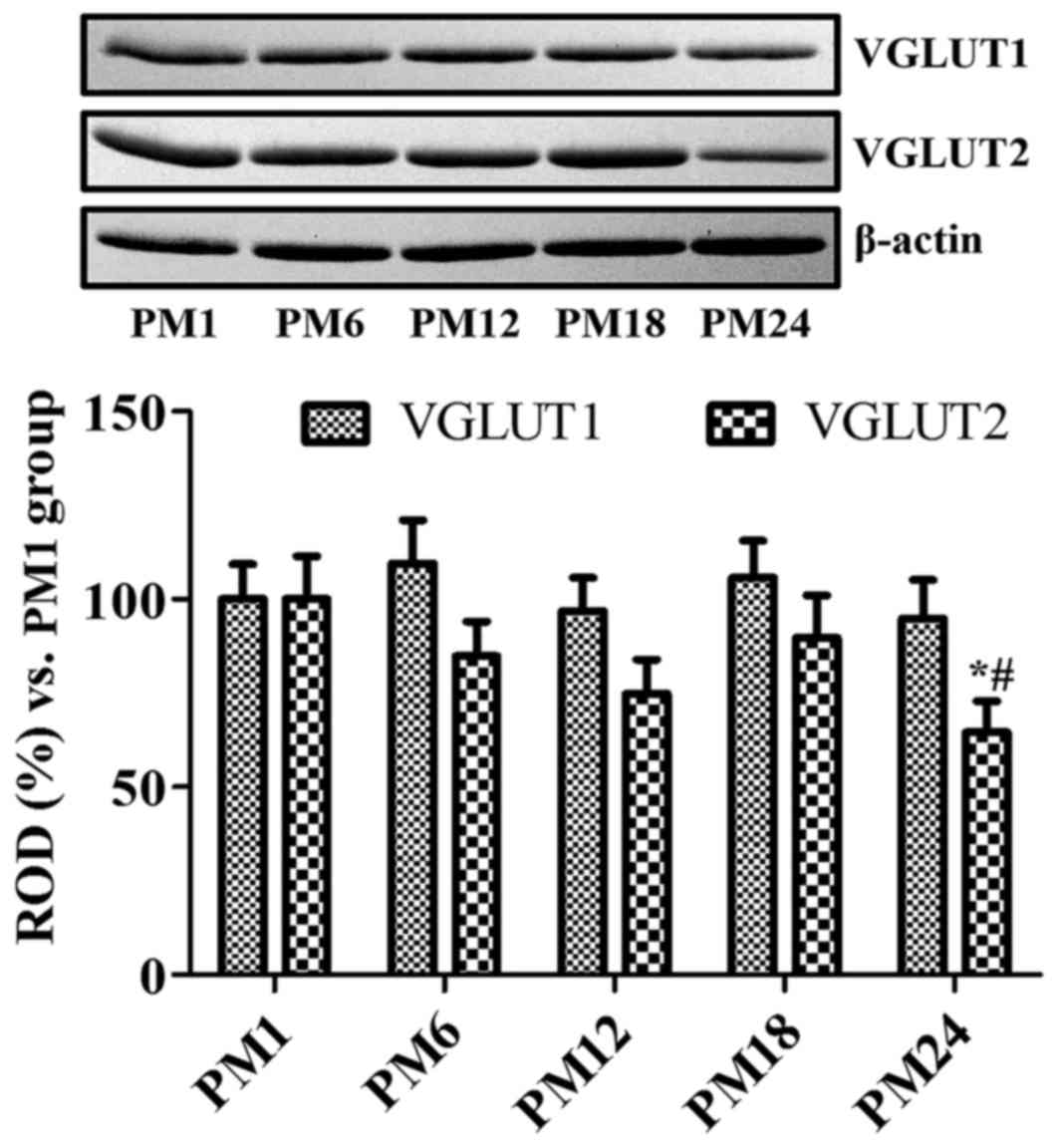
Age-associated changes in VGLUT
protein levels
VGLUT1 protein levels were higher in the PM6 and
PM24 groups when compared with the PM1 group; however,
statistically significant differences were not identified between
the different groups (Fig. 4).
VGLUT2 protein levels increased in the PM18 group when compared
with the PM12 group; however, no statistically significant
difference was detected. VGLUT2 protein levels were significantly
decreased in the PM24 group when compared with the PM1 and PM18
groups (Fig. 4).
Discussion
Glutamate is concentrated in synaptic vesicles by
VGLUTs (6), which are specifically
required for exocytic release (22,23).
In the present study, the specificity of VGLUT1 and VGLUT2
antibodies was tested in the cerebellum of gerbils. Localization of
VGLUT1 and VGLUT2 in gerbils was similar to that in the cerebellum
of rats (21,24,25).
The treatment with pre-immune serum completely blocked the
immunohistochemical staining for VGLUT1 and VGLUT2 in the
cerebellum and the hippocampus of gerbils.
VGLUT1 was expressed primarily in the mossy fibers
of the dentate gyrus, as well as the Schaffer collaterals and
stratum lacunosum-moleculare of the hippocampal CA2/3 regions.
VGLUT2 was observed in the granule cell layer and outer molecular
layer of the dentate gyrus, as well as the stratum pyramidale,
Schaffer collaterals and stratum lacunosum-moleculare of the
hippocampal CA1-3 regions. These results are consistent with
previous studies, in regard to the expression of VGLUT1 and 2
glutamatergic terminals (24,26–29).
In the present study, VGLUT1 was relatively resistant to the aging
process, except in the dentate gyrus; VGLUT1 immunoreactivity was
markedly increased in the outer molecular layer of the dentate
gyrus of the PM6 and PM24 groups compared with the PM1 group;
however, no significant differences in VGLUT1 protein levels were
observed in the hippocampal homogenates. This result is consistent
with a previous study using senescence-accelerated mice (SAMP8), in
that no significant changes were observed in VGLUT1 in the
hippocampus of 2-, 6- and 12-month-old control mice and SAMP8
(10). However, in Wistar rats
VGLUT1 expression was decreased in the hippocampus with aging at
PM18 (9). The slight increase in
VGLUT1 may be associated with mild cognitive impairment as VGLUT1
is upregulated under these conditions (14), and working and reference memory in
the gerbil starts to decrease at PM18 (30). In SAMP8 mice, memory impairments,
based on a T-maze foot-shock avoidance task, have been reported at
8 to 12 months of age (31).
In the present study, VGLUT2 tended to decrease with
age in the hippocampus. This result is consistent with a previous
study, which observed that VGLUT2 was seen to constantly decrease
with age from 12 months onwards in the hippocampus of Wistar rats
(9). In addition, a significant
decline in γ-aminobutyric acid (GABA) and glutamate levels have
been observed in the hippocampal homogenates of aged (12-month-old)
SAMP8 compared with those of the adult (2-month-old) SAMP8
(13). In the present study,
morphological evidence has been presented of significant increases
in VGLUT2 immunoreactivity in the hippocampal CA1-3 regions of the
PM18 group, particularly in the non-pyramidal cells, which then
markedly decreased in the hippocampus at PM24. Glutamic acid
decarboxylase, a rate-limiting enzyme for GABA synthesis,
immunoreactive interneurons in the CA1 region have been
demonstrated to be significantly decreased in the hippocampal CA1
region of middle-aged (15–17 months) and old-aged (25–29 months)
rats (32). Collectively, these
results suggested that the increase in glutamate and decrease in
GABA levels may be the cause of cell damage in the non-pyramidal
cells of middle-aged animals. In animal models of Alzheimer's
disease, glutamatergic and GABAergic presynaptic boutons are
increased during the early stages of the amyloid pathology
(33,34). Similarly, in humans with mild
cognitive impairment, glutamatergic presynaptic bouton density has
been shown to increase in the mid-frontal gyrus, while the brains
of patients with moderate and severe Alzheimer's disease have
exhibited a significant depletion in presynaptic bouton density
(14).
In conclusion, VGLUT1 and VGLUT2 are expressed
differentially in the hippocampus, and VGLUT1 is relatively
resistant to changes induced by the aging process; however, VGLUT2
appears to decrease in the hippocampus with age. The increase in
VGLUT2 in the non-pyramidal cells of the PM18 group may be closely
associated with the reduction in memory function during the aging
process of Mongolian gerbils. This result may be applicable to the
development of anti-aging drugs to modulate glutamate and the GABA
ratio in the hippocampus in the aging process, and the efficient
transport system of glutamate may targeted overcome the decreases
in hippocampal functions in the aging process.
Acknowledgements
The authors would like to thank Mr. Seung Uk Lee and
Mrs. Hyun Sook Kim for technical help in the present study.
Funding
The present study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education (grant no.
NRF-2013R1A1A2059364). The present study was also partially
supported by the Research Institute for Veterinary Science, Seoul
National University.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors contributions
HJ, DY and IH designed the experiments and the
study. HJ, DY, JP and JK looked after the animals and performed the
morphological experiments. DK conducted western blot analysis. JC,
MW and YY participated in designing and discussing the study. HJ
and IH wrote this manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The handling and care of the animals conformed to
guidelines compliant with the current international laws and
policies [National Institutes of Health (NIH) Guide for the Care
and Use of Laboratory Animals; NIH Publication no. 85-23, 1985,
revised 1996]. Ethical approval was obtained from the Institutional
Animal Care and Use Committee of Kangwon National University
(Chuncheon, Gangwon, Republic of Korea) for all animal procedures
in the present study (no. KW-160802-2).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Omote H and Moriyama Y: Vesicular
neurotransmitter transporters: An approach for studying
transporters with purified proteins. Physiology (Bethesda).
28:39–50. 2013.PubMed/NCBI
|
|
2
|
Südhof TC: The synaptic vesicle cycle: A
cascade of protein-protein interactions. Nature. 375:645–653. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Daniels RW, Collins CA, Chen K, Gelfand
MV, Featherstone DE and DiAntonio A: A single vesicular glutamate
transporter is sufficient to fill a synaptic vesicle. Neuron.
49:11–16. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ishikawa T, Sahara Y and Takahashi T: A
single packet of transmitter does not saturate postsynaptic
glutamate receptors. Neuron. 34:613–621. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wojcik SM, Rhee JS, Herzog E, Sigler A,
Jahn R, Takamori S, Brose N and Rosenmund C: An essential role for
vesicular glutamate transporter 1 (VGLUT1) in postnatal development
and control of quantal size. Proc Natl Acad Sci USA. 101:pp.
7158–7163. 2004; View Article : Google Scholar : PubMed/NCBI
|
|
6
|
El Mestikawy S, Wallén-Mackenzie A, Fortin
GM, Descarries L and Trudeau LE: From glutamate co-release to
vesicular synergy: Vesicular glutamate transporters. Nat Rev
Neurosci. 12:204–216. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tordera RM, Totterdell S, Wojcik SM, Brose
N, Elizalde N, Lasheras B and Del Rio J: Enhanced anxiety,
depressive-like behaviour and impaired recognition memory in mice
with reduced expression of the vesicular glutamate transporter 1
(VGLUT1). Eur J Neurosci. 25:281–290. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He H, Mahnke AH, Doyle S, Fan N, Wang CC,
Hall BJ, Tang YP, Inglis FM, Chen C and Erickson JD:
Neurodevelopmental role for VGLUT2 in pyramidal neuron plasticity,
dendritic refinement, and in spatial learning. J Neurosci.
32:15886–15901. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Canas PM, Duarte JM, Rodrigues RJ, Köfalvi
A and Cunha RA: Modification upon aging of the density of
presynaptic modulation systems in the hippocampus. Neurobiol Aging.
30:1877–1884. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cheng XR, Yang Y, Zhou WX and Zhang YX:
Expression of VGLUTs contributes to degeneration and acquisition of
learning and memory. Neurobiol Learn Mem. 95:361–375. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin L, Cao B, Xu Z, Sui Y, Chen J, Luan Q,
Yang R, Li S and Li KF: In vivo HMRS and lipidomic profiling
reveals comprehensive changes of hippocampal metabolism during
aging in mice. Biochem Biophys Res Commun. 470:9–14. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ménard C, Quirion R, Vigneault E, Bouchard
S, Ferland G, El Mestikawy S and Gaudreau P: Glutamate presynaptic
vesicular transporter and postsynaptic receptor levels correlate
with spatial memory status in aging rat models. Neurobiol Aging.
36:1471–1482. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang H, Lian K, Han B, Wang Y, Kuo SH,
Geng Y, Qiang J, Sun M and Wang M: Age-related alterations in the
metabolic profile in the hippocampus of the senescence-accelerated
mouse prone 8: A spontaneous Alzheimer's disease mouse model. J
Alzheimers Dis. 39:841–848. 2014.PubMed/NCBI
|
|
14
|
Bell KF, Bennett DA and Cuello AC:
Paradoxical upregulation of glutamatergic presynaptic boutons
during mild cognitive impairment. J Neurosci. 27:10810–10817. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kashani A, Lepicard E, Poirel O, Videau C,
David JP, Fallet-Bianco C, Simon A, Delacourte A, Giros B, Epelbaum
J, et al: Loss of VGLUT1 and VGLUT2 in the prefrontal cortex is
correlated with cognitive decline in Alzheimer disease. Neurobiol
Aging. 29:1619–1630. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Buchhalter JR: Animal models of inherited
epilepsy. Epilepsia. 34 Suppl 3:S31–S41. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Paul LA, Fried I, Watanabe K, Forsythe AB
and Scheibel AB: Structural correlates of seizure behavior in the
Mongolian gerbil. Science. 213:924–926. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Troup GM, Smith GS and Walford RL: Life
span, chronologic disease patterns, and age-related changes in
relative spleen weights for the mongolian gerbil (Meriones
unguiculatus). Exp Gerontol. 4:139–143. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
[NRC] National Research Council: Guide for
the Care and Use of Laboratory Animals. 7th. National Academy
Press; Washington DC: 1996
|
|
20
|
Loskota WA, Lomax P and Verity MA: A
stereotaxic atlas of the Mongolian Gerbil Brain (Meriones
unguiculatus)Ann Arbor Science Publishers Inc.; Ann Arbor, MI:
pp. 70–79. 1974
|
|
21
|
Hioki H, Fujiyama F, Taki K, Tomioka R,
Furuta T, Tamamaki N and Kaneko T: Differential distribution of
vesicular glutamate transporters in the rat cerebellar cortex.
Neuroscience. 117:1–6. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Takamori S, Rhee JS, Rosenmund C and Jahn
R: Identification of a vesicular glutamate transporter that defines
a glutamatergic phenotype in neurons. Nature. 407:189–194. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takamori S: VGLUTs: ‘Exciting’ times for
glutamatergic research? Neurosci Res. 55:343–351. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fremeau RT Jr, Troyer MD, Pahner I,
Nygaard GO, Tran CH, Reimer RJ, Bellocchio EE, Fortin D,
Storm-Mathisen J and Edwards RH: The expression of vesicular
glutamate transporters defines two classes of excitatory synapse.
Neuron. 31:247–260. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Herzog E, Bellenchi GC, Gras C, Bernard V,
Ravassard P, Bedet C, Gasnier B, Giros B and El Mestikawy S: The
existence of a second vesicular glutamate transporter specifies
subpopulations of glutamatergic neurons. J Neurosci. 21:RC1812001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fremeau RT Jr, Kam K, Qureshi T, Johnson
J, Copenhagen DR, Storm-Mathisen J, Chaudhry FA, Nicoll RA and
Edwards RH: Vesicular glutamate transporters 1 and 2 target to
functionally distinct synaptic release sites. Science.
304:1815–1819. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fujiyama F, Furuta T and Kaneko T:
Immunocytochemical localization of candidates for vesicular
glutamate transporters in the rat cerebral cortex. J Comp Neurol.
435:379–387. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Halasy K, Hajszan T, Kovács EG, Lam TT and
Leranth C: Distribution and origin of vesicular glutamate
transporter 2-immunoreactive fibers in the rat hippocampus.
Hippocampus. 14:908–918. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Herzog E, Takamori S, Jahn R, Brose N and
Wojcik SM: Synaptic and vesicular co-localization of the glutamate
transporters VGLUT1 and VGLUT2 in the mouse hippocampus. J
Neurochem. 99:1011–1018. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hwang IK, Yoo KY, Jung BK, Cho JH, Kim DH,
Kang TC, Kwon YG, Kim YS and Won MH: Correlations between neuronal
loss, decrease of memory, and decrease expression of brain-derived
neurotrophic factor in the gerbil hippocampus during normal aging.
Exp Neurol. 201:75–83. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Flood JF and Morley JE: Age-related
changes in footshock avoidance acquisition and retention in
senescence accelerated mouse (SAM). Neurobiol Aging. 14:153–157.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shi L, Argenta AE, Winseck AK and
Brunso-Bechtold JK: Stereological quantification of
GAD-67-immunoreactive neurons and boutons in the hippocampus of
middle-aged and old Fischer 344 × Brown Norway rats. J Comp Neurol.
478:282–291. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bell KF, de Kort GJ, Steggerda S,
Shigemoto R, Ribeiro-da-Silva A and Cuello AC: Structural
involvement of the glutamatergic presynaptic boutons in a
transgenic mouse model expressing early onset amyloid pathology.
Neurosci Lett. 353:143–147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu L, Wong TP, Côté SL, Bell KF and Cuello
AC: The impact of Aβ-plaques on cortical cholinergic and
non-cholinergic presynaptic boutons in Alzheimer's disease-like
transgenic mice. Neuroscience. 121:421–432. 2003. View Article : Google Scholar : PubMed/NCBI
|