Introduction
Gastric cancer (GC), developing from the lining of
the stomach, is one of the most common cancers worldwide,
particularly in East Asia and China (1). It is the third leading cause of death
from cancer and accounts for 9% of mortality worldwide (2). If untreated, tumor cells often
metastasize to other parts of the body, particularly the lungs,
liver, bone, and lymph nodes; therefore, the prognosis of GC is
generally unfavorable (3). The
5-year survival rate for GC is reported to be <10% (4). In China, the majority of patients
with GC are diagnosed at a late stage and the prognosis is
unfavorable (1). Therefore,
understanding of the molecular mechanisms and identification of the
key biomarkers associated with GC progression is essential for the
diagnosis and therapy of GC.
The conventional view of gene regulation in biology
is primarily concentrated on the protein-coding genes. However, the
human genome project suggested that ~1.2% of the mammalian genome
encodes proteins (5,6) and most of the genome is transcribed
into long non-coding RNAs (lncRNAs) (7,8).
LncRNAs are RNA molecules >200 nucleotides in length.
Dysregulated lncRNAs have been demonstrated to have an important
role in tumorigenesis and cancer metastasis (9–11).
The association between aberrant expression of lncRNAs and GC has
been previously investigated. For example lncRNA-HMlincRNA717 was
determined to have a crucial role during GC occurrence and
progression (12). Song et
al (13) performed lncRNA
microarray analysis and identified 135 differentially expressed
lncRNAs between GC and normal tissues (13). However, numerous lncRNAs have been
identified they are not sufficient for the treatment of GC.
In the present study, the lncRNA sequencing for GC
tissues was performed using a transcriptome sequencing technique.
The differentially expressed lncRNAs between GC and normal adjacent
tissues were identified. The bioinformatics analysis included
prediction of target genes and function enrichment analysis.
Finally, the lncRNAs predicted by the present study were verified
by reverse transcription-quantitative polymerase chain reaction
(RT-qPCR). The current study aimed to investigate the additional
lncRNAs associated with GC, which may be used as potential markers
for the diagnosis and treatment of GC.
Materials and methods
Tissue samples
Between October 2015 and January 2016, a total of 3
male patients with GC (aged 65–76 years old) were included in the
current study, whose diagnoses were pathologically confirmed. The
cancer tissues and the normal adjacent tissues were obtained from
clinically ongoing surgical specimens, were snap frozen with liquid
nitrogen and subsequently stored at −80°C until RNA extraction.
All patients have provided written informed consent
prior to participating in the present study. The procedures in the
current study were approved by the Protection of Human Ethics
Committee of Shanghai Shuguang Hospital Affiliated with Shanghai
University of TCM (Shanghai, China).
Transcriptome sequencing
Total RNAs from gastric cancer tissues (3 samples)
and normal adjacent tissues (3 samples) were extracted using the
RNAiso Plus (Takara Biotechnology Co., Ltd., Dalian, China).
Evaluation of the quality and integrity of the total RNA was
performed using 1% agarose gel electrophoresis (visualized using
ethidium bromide), and an Agilent 2100 Bioanalyzer (Agilent
Technologies, Inc., Santa Clara, CA, USA). Following this, a cDNA
library was established using the NEBNext Ultra RNA Library Prep
kit (New England Biolabs, Inc., Ipswich, MA, USA) prior to Illumina
sequencing. The transcriptome sequencing of mRNA and lncRNA was
performed on an Illumina gene analyzer (Illumina, Inc., San Diego,
USA).
The data have been deposited at National Center for
Biotechnology Information (NCBI) Sequence Read Archive (SRA)
database under accession number: SRP092509.
Quality control of the sequencing data was performed
to identify the clean reads using the FASTX-toolkit (version
0.0.13) (14). The obtained clean
reads were aligned to the human reference genome hg19 using TopHat
software (version 2.10) (15).
Then based on the mRNA and lncRNA annotation information provided
by gencode version 24 (mapped to GRCh37) (16) database, the Fragments Per Kilobase
of transcript per Million mapped reads values of mRNA and lncRNA
and the reads number of lncRNA were identified using StringTie tool
(version 1.2.3) (17).
Bioinformatics analysis of sequencing
data
The differentially expressed lncRNAs and genes
(DEGs) between the cancer group and the control group were
identified using the limma package (18) in R (version 3.2.5) with the
following criteria: |log2 fold change (FC)|>1 and
P<0.05.
The downstream target genes of the differentially
expressed lncRNAs were predicted based on the co-expression
associations between lncRNAs and mRNAs. The threshold values were
correlation coefficient >0.8 and P<0.05. Additionally, the
network of lncRNAs and their target genes was constructed using
Cytoscape (version 3.0) (19).
Subsequently, DEGs and target genes of lncRNAs were
used to perform Gene Ontology (GO) functional and Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway enrichment
analyses with the clusterprofiler package (20) in R.
RT-qPCR verification of the expression
of lncRNAs
Total RNA was extracted from tissues (3 GC tissues
and 3 normal adjacent tissues) using RNAiso Plus (9109; Takara
Biotechnology Co., Ltd.). The concentration and purity of the
isolated RNA was determined using TECAN infinite M100 PRO Biotek
microplate reader (Tecan Group, Ltd., Mannedorf, Switzerland) and
reverse transcription (37°C for 15 min and 85°C for 5 sec) was
performed according to the PrimeScript RT Master mix RR036A (Takara
Biotechnology Co., Ltd.). qPCR was performed using SYBRGreen kit
(cat no. 4367659; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The reaction procedures were as follows: 50°C for 3 min, 95°C
for 3 min, 95°C for 10 sec, and 60°C for 30 sec, for 40 cycles, the
melting process was 60 to 95°C (increments of 0.5°C for 10 sec).
According to the results of bioinformatics analysis, the
expressions of 7 lncRNAs, including AC016735.2, RP11-243M5.2,
RP11-400N13.2, RP11-400N13.3, AP001626.1, LINC01139 and
RP11-54H7.4, were detected with the primers presented in Table I. The expression levels were
calculated using the 2−ΔΔCq method (21).
 | Table I.Primer sequences of the long noncoding
RNAs for reverse transcription- quantitative polymerase chain
reaction. |
Table I.
Primer sequences of the long noncoding
RNAs for reverse transcription- quantitative polymerase chain
reaction.
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|
| AC016735.2 |
CTGCTTCTCACTGCCTCG |
TTTCCCAAATGGTCCTCC |
| RP11-243M5.2 |
TTGCGTGAAAGCGTATGG |
GAAAGCAGCCTTGAGAACAGAG |
| RP11-400N13.2 |
CCCCTGTCCTCCTGCTCTT |
CGGGCAGTGTCAGTCTTCA |
| RP11-400N13.3 |
GCAGATGGCAAAGGATAAAGC |
GGTGATATACGATGCAACGGTG |
| AP001626.1 |
AGCTGCACCAAGGAGAATC |
CAAAGCCAAGGTCCACTGTT |
| LINC01139 |
ACCAGTCACCCAACCAGAGC |
AAGCGTAAGAATGAAGACCAGTG |
| RP11-54H7.4 |
TCCACTCTAGGTTCCCACG |
CCTGACATTCCTGCCTTCTT |
| GAPDH |
TGACAACTTTGGTATCGTGGAAGG |
AGGCAGGGATGATGTTCTGGAGAG |
Statistical analysis
Data are presented as the mean ± standard error of
mean. The statistical analysis was performed by Graphpad Prism
(version 5.01) using the Student's t-test (Graphpad Software, Inc.,
San Diego, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
High-throughput sequencing
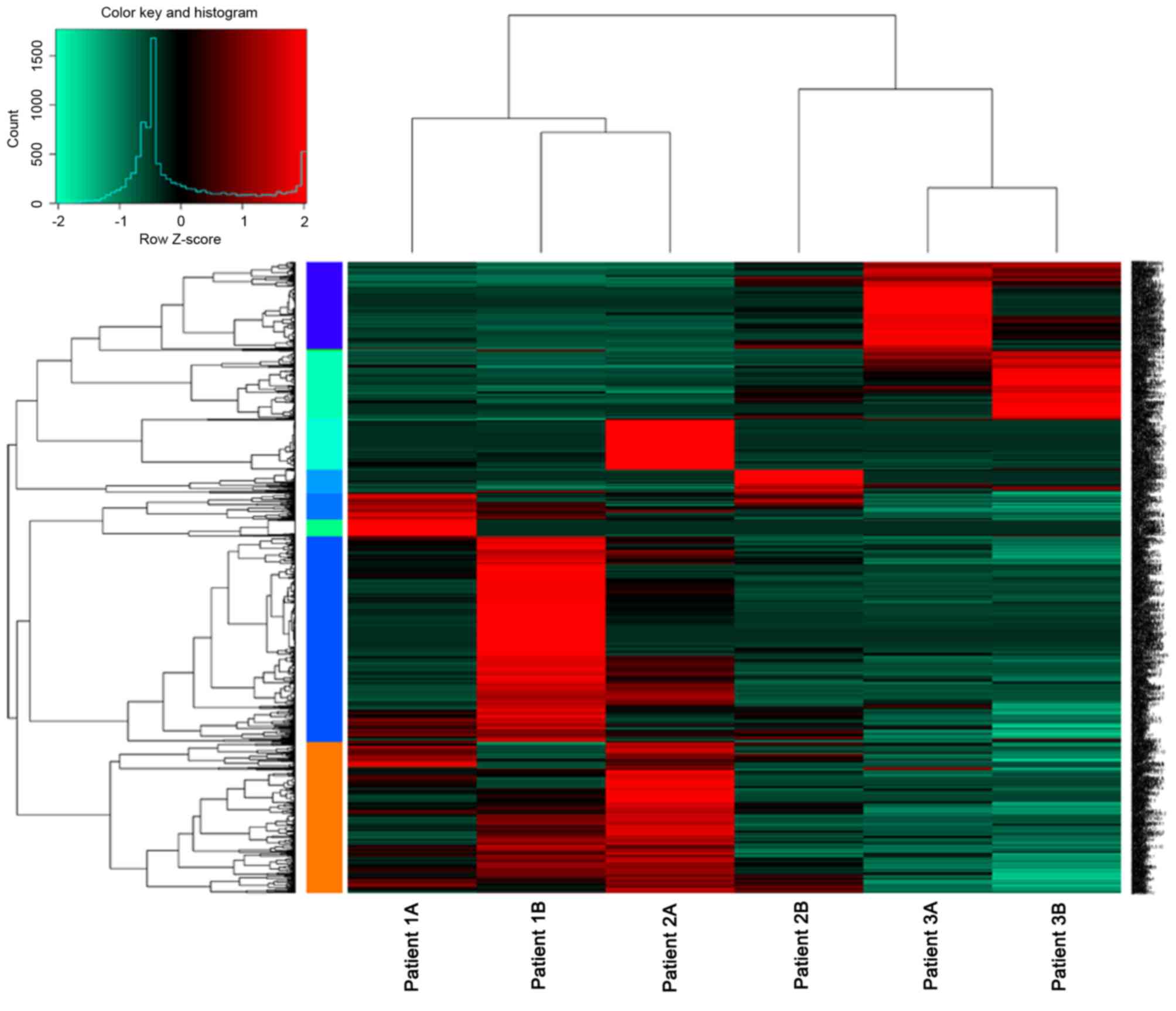
From the 6 samples, a total of 3,4290 mRNAs and
10,148 lncRNAs were identified, which were expressed in at least
one sample. With the criteria of|log2FC|>1 and
P<0.05, a total of 1,181 DEGs were identified, 902 were
upregulated and 279 were downregulated. Additionally, 390
differentially expressed lncRNAs, including 163 upregulated and 227
upregulated lncRNAs were identified (Fig. 1). The top 10 differentially
expressed lncRNA with higher FCs, including RP11-171I2.1,
RP11-171I2.1 and AC016735.2 are presented in Table II.
| Table II.Top 10 differentially expressed
upregulated and downregulated lncRNAs. |
Table II.
Top 10 differentially expressed
upregulated and downregulated lncRNAs.
| A, Downregulated |
|---|
|
|---|
| ID | logFC | P-value |
|---|
| RP11-171I2.1 | −4.549491715 | 0.006324281 |
| RP5-994D16.9 | −4.73821376 | 0.002778706 |
| RP11-139H15.6 | −4.802835645 | 0.002652066 |
| RP11-637N19.1 | −4.807460785 | 0.005211747 |
| RP11-243M5.2 | −4.87029533 | 0.011518821 |
| RP11-382A20.5 | −4.909287572 | 0.002375869 |
| AC003090.1 | −5.054470484 | 0.002583845 |
| FGF14-IT1 | −5.152474325 | 0.004193094 |
| AC053503.12 | −5.200695133 | 0.045876461 |
| RP11-16P20.4 | −5.309441725 | 0.00370217 |
|
| B,
Upregulated |
|
| ID | logFC | P-value |
|
| RP3-416H24.1 | 6.312295751 | 0.002010312 |
| RP5-1185I7.1 | 5.627125267 | 0.002013113 |
| LINC01087 | 5.562031916 | 0.00323185 |
| RP11-1007G5.2 | 5.506776995 | 0.000837008 |
| LINC00483 | 5.440975498 | 0.01629253 |
| LINC00618 | 5.425270941 | 0.000784189 |
| RP11-120K24.4 | 5.413330508 | 0.002061786 |
| AC016735.2 | 5.383341519 | 0.030647411 |
| AC110769.3 | 5.338202236 | 0.001217988 |
| LA16c-325D7.1 | 5.296535904 | 0.002461903 |
Bioinformatics analysis of lncRNA
sequencing data
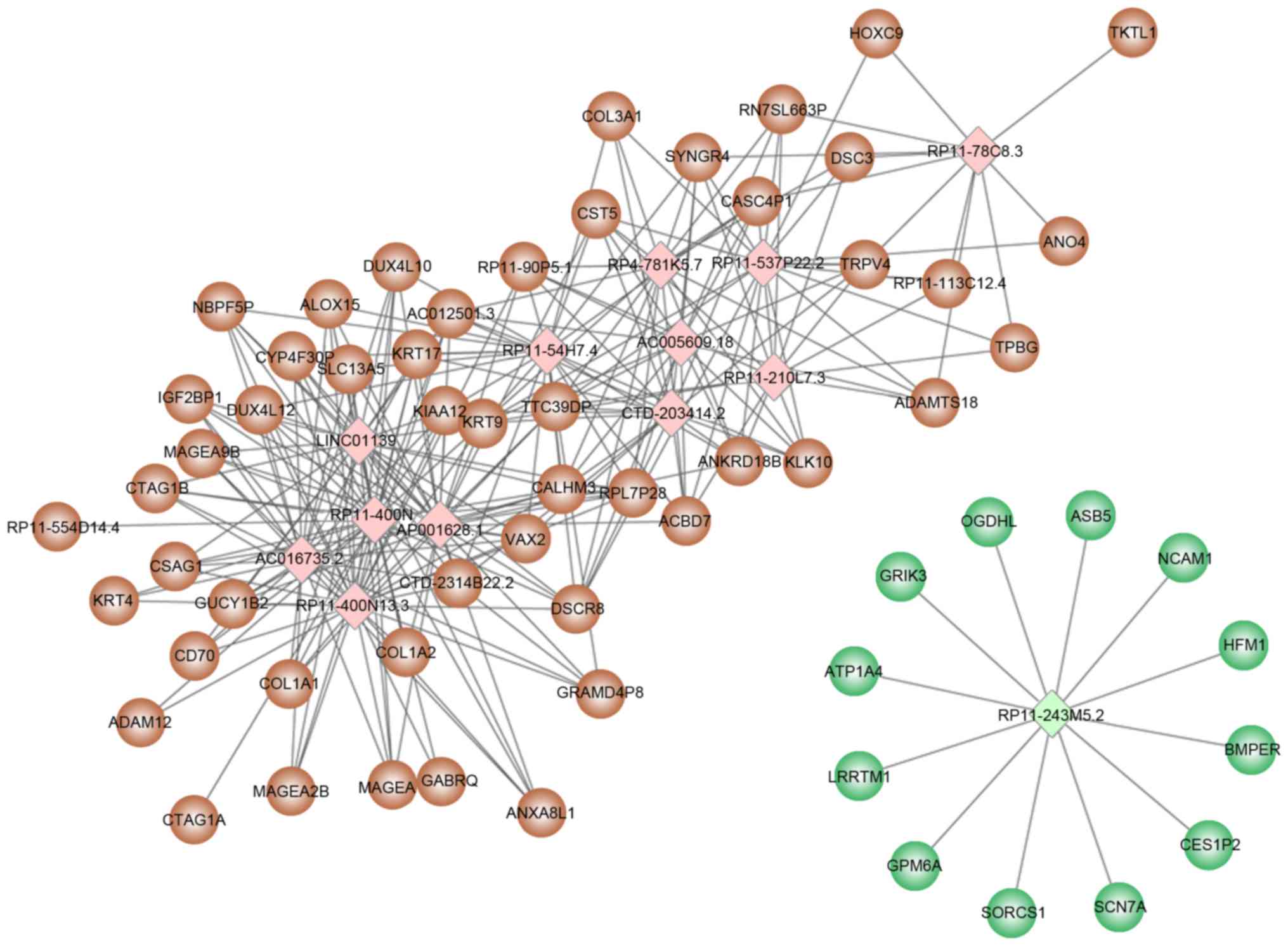
Based on the threshold values of correlation
coefficient >0.8 and P<0.05, 157 differentially expressed
lncRNAs were selected for target prediction and a total of 231
target genes were identified. Among the 157 differentially
expressed lncRNAs, 13 lncRNAs (12 upregulated and 1 downregulated)
predicted an additional 10 target genes (Table III), including RP11-400N13.2,
RP11-400N13.3, AP001626.1 and RP11-54H7.4. The regulatory network
constructed with 13 lncRNAs and their target genes is presented in
Fig. 2.
| Table III.Long noncoding RNAs with more than 10
target genes. |
Table III.
Long noncoding RNAs with more than 10
target genes.
| ID | No of target
genes |
log2FC | P-value |
|---|
| AC005609.18 | 16 | 5.258702066 | 0.002564097 |
| AC016735.2 | 23 | 5.383341519 | 0.030647411 |
| AP001626.1 | 30 | 3.480674967 | 0.039101426 |
| CTD-2034I4.2 | 19 | 3.225981981 | 0.039614051 |
| LINC01139 | 27 | 3.784674378 | 0.034659687 |
| RP11-210L7.3 | 15 | 3.183358792 | 0.03741905 |
| RP11-243M5.2 | 12 | −4.87029533 | 0.011518821 |
| RP11-400N13.2 | 32 | 4.411473414 | 0.015832445 |
| RP11-400N13.3 | 32 | 4.01570403 | 0.031992561 |
| RP11-537P22.2 | 19 | 3.519296946 | 0.027224444 |
| RP11-54H7.4 | 25 | 3.227024693 | 0.043767147 |
| RP11-782C8.3 | 11 | 3.386174545 | 0.029429789 |
| RP4-781K5.7 | 18 | 3.607427164 | 0.025935631 |
Due to the high number of lncRNAs identified, the
functions of individual lncRNAs were not analyzed. The present
study focused on the functions of the 13 aforementioned lncRNAs. A
total of 50 and 12 target genes were predicted for the upregulated
and downregulated lncRNAs, respectively. Functional enrichment
analysis of the 62 target genes determined that the upregulated
lncRNAs were significantly enriched in functions associated with
collagen fibril organization, whereas the downregulated lncRNA was
significantly associated with ion transmembrane transport and
regulation of membrane potential (Table IV).
| Table IV.Functional enrichment analysis of the
target genes of the 13 lncRNAs with more than 10 target genes. |
Table IV.
Functional enrichment analysis of the
target genes of the 13 lncRNAs with more than 10 target genes.
| A, Upregulated |
|---|
|
|---|
| Ontology | ID | Function | Count | FDR |
|---|
| BP | GO:0043588 | Skin
development | 5 | 0.011708982 |
| BP | GO:0030199 | Collagen fibril
organization | 3 | 0.011708982 |
| BP | GO:0071230 | Cellular response
to amino acid stimulus | 3 | 0.012776741 |
| BP | GO:0043589 | Skin
morphogenesis | 2 | 0.012776741 |
| BP | GO:0070208 | Protein
heterotrimerization | 2 | 0.016397947 |
| CC | GO:0005583 | Fibrillar collagen
trimer | 3 | 0.000116529 |
| CC | GO:0098643 | Banded collagen
fibril | 3 | 0.000116529 |
| CC | GO:0098644 | Complex of collagen
trimers | 3 | 0.000426968 |
| CC | GO:0005581 | Collagen
trimer | 3 | 0.016659231 |
| CC | GO:0044420 | Extracellular
matrix component | 3 | 0.037435206 |
| MF | GO:0048407 | Platelet-derived
growth factor binding | 3 | 0.000119244 |
| MF | GO:0005201 | Extracellular
matrix structural constituent | 3 | 0.016290955 |
| F | GO:0005198 | Structural molecule
activity | 6 | 0.036696011 |
| MF | GO:0019838 | Growth factor
binding | 3 | 0.040871579 |
| KEGG | hsa04974 | Protein digestion
and absorption | 3 | 0.002473586 |
| KEGG | hsa05146 | Amoebiasis | 3 | 0.002473586 |
| KEGG | hsa04933 | AGE-RAGE signaling
pathway in diabetic complications | 3 | 0.002473586 |
| KEGG | hsa04611 | Platelet
activation | 3 | 0.003235067 |
| KEGG | hsa04512 | ECM-receptor
interaction | 2 | 0.026270733 |
|
| B,
Downregulated |
|
|
Ontology | ID |
Function | Count | FDR |
|
| BP | GO:0055078 | Sodium ion
homeostasis | 2 | 0.164234693 |
| BP | GO:0042391 | Regulation of
membrane potential | 3 | 0.164234693 |
| BP | GO:0034220 | Ion transmembrane
transport | 4 | 0.164234693 |
| BP | GO:0007416 | Synapse
assembly | 2 | 0.164234693 |
| BP | GO:0035725 | Sodium ion
transmembrane transport | 2 | 0.164234693 |
| CC | GO:1902495 | Transmembrane
transporter complex | 3 | 0.02856848 |
| CC | GO:1990351 | Transporter
complex | 3 | 0.02856848 |
| CC | GO:0098794 | postsynapse | 3 | 0.02856848 |
| CC | GO:0030424 | Axon | 3 | 0.02856848 |
| CC | GO:0030426 | Growth cone | 2 | 0.041273443 |
| MF | GO:0015075 | Ion transmembrane
transporter activity | 4 | 0.011272955 |
| MF | GO:0022891 | Substrate-specific
transmembrane transporter activity | 4 | 0.011272955 |
| MF | GO:0022857 | Transmembrane
transporter activity | 4 | 0.011272955 |
| MF | GO:0005216 | Ion channel
activity | 3 | 0.011272955 |
| MF | GO:0046873 | Metal ion
transmembrane transporter activity | 3 | 0.011272955 |
RT-qPCR verification of the expression
of lncRNAs
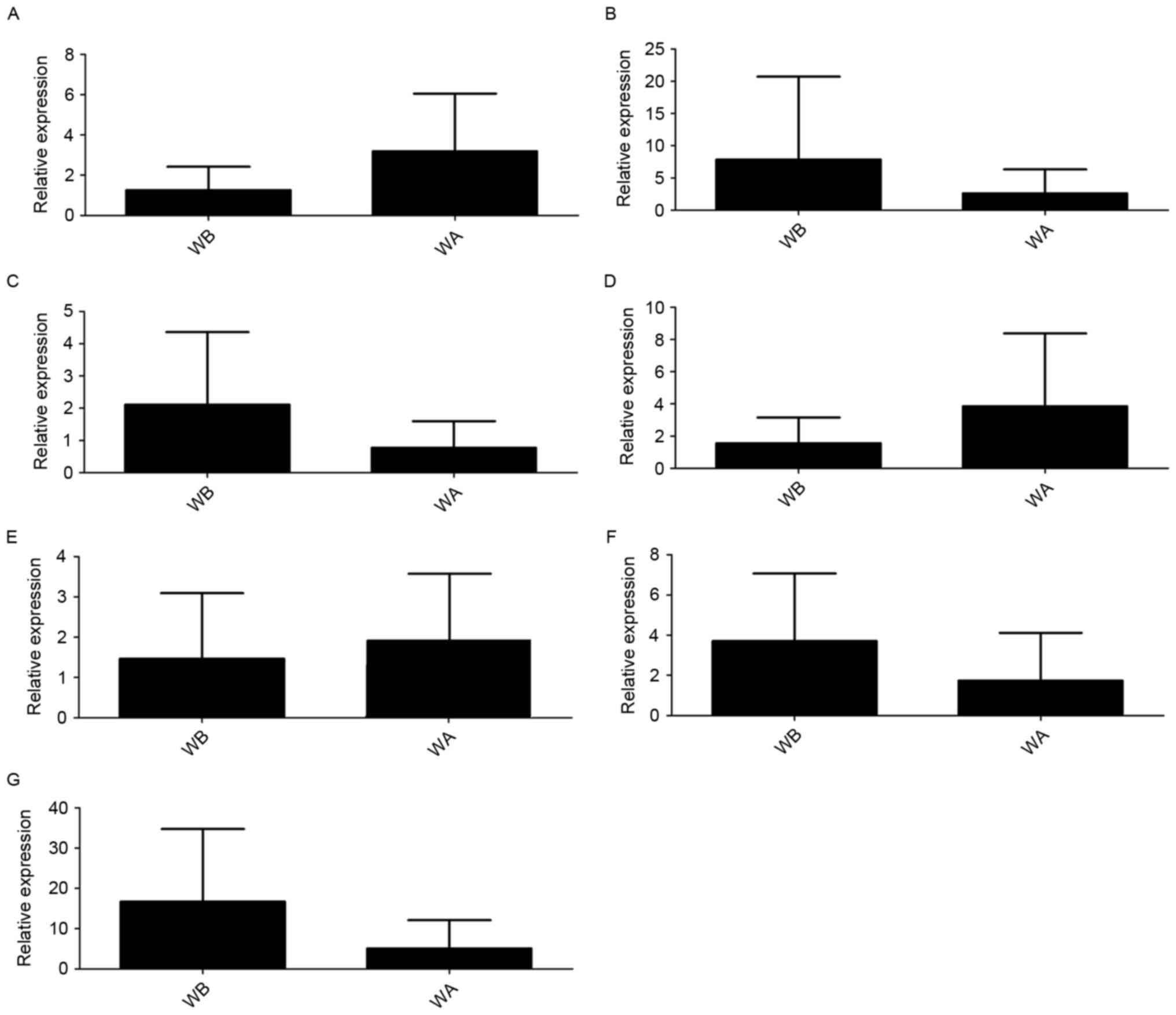
According to the findings of the transcriptome
sequencing and bioinformatics analyses, AC016735.2, RP11-243M5.2,
RP11-400N13.2, RP11-400N13.3, AP001626.1, LINC01139 and RP11-54H7.4
were upregulated, and only RP11-243M5.2 was downregulated.
The RT-qPCR validation confirmed that AC016735.2,
AP001626.1 and RP11-400N13.3 were upregulated, whereas
RP11-243M5.2, RP11-400N13.2, LINC01139 and RP11-54H7.4 were
downregulated in GC tissues compared with normal adjacent tissues.
It is of note that although upregulation and downregulation were
detected, no significant difference was identified (Fig. 3). The findings of AC016735.2,
AP001626.1, RP11-400N13.3 and RP11-243M5.2 were considered to be
consistent with the predicted lncRNAs in the bioinformatics
analysis.
Discussion
Previous studies have identified lncRNAs to be
important in the governing of fundamental biological processes,
where aberrant expression may be associated with various human
cancers (22,23). The present study identified 390
differentially expressed lncRNAs between GC and normal adjacent
tissues via transcriptome sequencing and bioinformatics analysis.
The upregulated lncRNAs were significantly enriched in functions
associated with collagen fibril organization, whereas the
downregulated lncRNA was significantly associated with ion
transmembrane transport and regulation of membrane potential.
Following RT-qPCR validation, AC016735.2, AP001626.1, RP11-400N13.3
and RP11-243M5.2 were considered to be consistent with the results
of the bioinformatics prediction, suggesting that they may have a
role in the tumorigenesis of GC.
RP11-400N13.3, AP001626.1 and AC016735.2 were all
upregulated lncRNAs, and were predicted to regulate >10 target
genes, including collagen type I a 1 (COL1A1),
COL1A2, and arachidonate 15-lipoxygenase (ALOX15). It
is of note that the three target genes were also DEGs.
COL1A1 and COL1A2 encode type I collagen, which is
the most abundant collagen of the human body that forms collagen
fibers. COL1A1 and COL1A2 were identified to be
significantly enriched in GO function associated with collagen
fibril organization. Type I collagen has an important role in
fibrosis and cancer progression (24). A previous study determined that
collagen is a major contributor to diffusive hindrance in human
tumors (25). Additionally, type I
collagen is a prevalent component of the stromal extracellular
matrix (26). The stromal
extracellular matrix is a barrier to a progressing cancer cell,
changes of which contribute to metastasis in cancer (27). Therefore, it is possible that the
three lncRNAs may be involved in GC metastasis by regulating
COL1A1 and COL1A2.
ALOX15 encoding protein is part of the
lipoxygenases family. Human lipoxygenases are widely distributed in
human organs, tissues and cells (28), catalyzing peroxidation of
unsaturated fatty acid producing various types of eicosanoids
(29). It has been previously
reported that many cancers are driven by lipoxygenases and their
metabolites (30,31). A previous study determined that
ALOX15 is an important factor in the regulation of colorectal
epithelial cell terminal differentiation and apoptosis (32). In the current study, ALOX15
was significantly enriched in engulfment of apoptotic cell
(GO:0043652), which may confirm its role in apoptosis. Therefore,
RP11-400N13.3, AP001626.1 and AC016735.2 may also regulate
ALOX15 and have a role in the progression of GC.
RP11-243M5.2 was a downregulated lncRNA and had 12
target genes, including ATPase Na+/K+ transporting subunit a 4
(ATP1A4) and sodium voltage-gated channel a subunit 7
(SCN7A), which were significantly involved in functions
associated with ion homeostasis and ion transmembrane transport. It
has been previously established that cells require a balance of
ions across their cell membrane in order to ensure cell survival.
The homeostatic intracellular ionic environment is necessary for
the correct functioning of gene expression, hormone release and
cellular proteins (33,34). It is of note that Bortner and
Cidlowski (35) have reported that
intracellular ion homeostasis has an important role in the
regulation of the cell death and changes may alter the apoptotic
rate of cells. Evading apoptosis by generating genetic mutations is
a key mechanism of carcinogenesis (36). Therefore, the present study
suggested that RP11-243M5.2 may have a role in the carcinogenesis
of GC by regulating ATP1A4 and SCN7A to participate
in functions associated to ion homeostasis and ion transmembrane
transport.
AC016735.2, AP001626.1, RP11-400N13.3 and
RP11-243M5.2 were verified by RT-qPCR; however, the results were
not statistically significant. This may be due to the heterogeneity
between the samples used in the transcriptome sequencing and
RT-qPCR.
In conclusion, by transcriptome sequencing and
RT-qPCR experiments the present study identified 4 lncRNAs,
including AC016735.2, AP001626.1, RP11-400N13.3 and RP11-243M5.2 to
have an important role in the pathogenesis of GC. They may be used
as potential diagnosis or treatment biomarkers of GC in the
future.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YW drafted the manuscript and acquired and analysed
the data. JZ interpreted the data and revised the manuscript.
Ethics approval and consent to
participate
The procedures in the current study were approved by
the Protection of Human Ethics Committee of Shanghai Shuguang
Hospital Affiliated with Shanghai University of TCM (Shanghai,
China).
Consent for publication
All patients have provided written informed consent
prior to participating in the present study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Li H, Yu B, Li J, Su L, Yan M, Zhu Z and
Liu B: Overexpression of lncRNA H19 enhances carcinogenesis and
metastasis of gastric cancer. Oncotarget. 5:2318–2329. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
IARC: World Cancer Report 2014. World
Health Organization; Geneva: 2015
|
|
3
|
Ruddon RW: Cancer Biology. Oxford Univ Pr.
2007.
|
|
4
|
Orditura M, Galizia G, Sforza V, et al:
Treatment of gastric cancer. World J Gastroenterol. 1635–1649.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
ENCODE Project Consortium, . Birney E,
Stamatoyannopoulos JA, Dutta A, Guigó R, Gingeras TR, Margulies EH,
Weng Z, Snyder M, Dermitzakis ET, et al: Identification and
analysis of functional elements in 1% of the human genome by the
ENCODE pilot project. Nature. 447:799–816. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Johnson JM, Edwards S, Shoemaker D and
Schadt EE: Dark matter in the genome: Evidence of widespread
transcription detected by microarray tiling experiments. Trends
Genet. 21:93–102. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Furuno M, Pang KC, Ninomiya N, Fukuda S,
Frith MC, Bult C, Kai C, Kawai J, Carninci P, Hayashizaki Y,
Mattick JS and Suzuki H: Clusters of internally primed transcripts
reveal novel long noncoding RNAs. PLoS Genet. 2:e372006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shore AN, Herschkowitz JI and Rosen JM:
Noncoding RNAs Involved in mammary gland development and
tumorigenesis: There's a long way to go. J Mammary Gland Biol
Neoplasia. 17:43–58. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huarte M, Guttman M, Feldser D, Garber M,
Koziol MJ, Kenzelmann-Broz D, Khalil AM, Zuk O, Amit I, Rabani M,
et al: A large intergenic noncoding RNA induced by p53 mediates
global gene repression in the p53 response. Cell. 142:409–419.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Whitehead J, Pandey GK and Kanduri C:
Regulation of the mammalian epigenome by long noncoding RNAs.
Biochim Biophys Acta. 1790:936–947. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shao Y, Chen H, Jiang X, Chen S, Li P, Ye
M, Li Q, Sun W and Guo J: Low expression of lncRNA-HMlincRNA717 in
human gastric cancer and its clinical significances. Tumour Biol.
35:9591–9595. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Song H, Sun W, Ye G, Ding X, Liu Z, Zhang
S, Xia T, Xiao B, Xi Y and Guo J: Long non-coding RNA expression
profile in human gastric cancer and its clinical significances. J
Transl Med. 11:2252013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schmieder R and Edwards R: Quality control
and preprocessing of metagenomic datasets. Bioinformatics.
27:863–864. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Trapnell C, Pachter L and Salzberg SL:
TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics.
25:1105–1111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Harrow J, Frankish A, Gonzalez JM,
Tapanari E, Diekhans M, Kokocinski F, Aken BL, Barrell D, Zadissa
A, Searle S, et al: GENCODE: The reference human genome annotation
for The ENCODE Project. Genome Res. 22:1760–1774. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pertea M, Pertea GM, Antonescu CM, Chang
TC, Mendell JT and Salzberg SL: StringTie enables improved
reconstruction of a transcriptome from RNA-seq reads. Nat
Biotechnol. 33:290–295. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hauptman N and Glavač D: Long non-coding
RNA in cancer. Int J Mol Sci. 14:4655–4669. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qiu MT, Hu JW, Yin R and Xu L: Long
noncoding RNA: An emerging paradigm of cancer research. Tumour
Biol. 34:613–620. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mueller MM and Fusenig NE: Friends or
foes-bipolar effects of the tumour stroma in cancer. Nat Rev
Cancer. 4:839–849. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ramanujan S, Pluen A, Mckee TD, Brown EB,
Boucher Y and Jain RK: Diffusion and convection in collagen gels:
Implications for transport in the tumor interstitium. Biophys J.
83:1650–1660. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Keely PJ, Wu JE and Santoro SA: The
spatial and temporal expression of the alpha 2 beta 1 integrin and
its ligands, collagen I, collagen IV, and laminin, suggest
important roles in mouse mammary morphogenesis. Differentiation.
59:1–13. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stewart DA, Cooper CR and Sikes RA:
Changes in extracellular matrix (ECM) and ECM-associated proteins
in the metastatic progression of prostate cancer. Reprod Biol
Endocrinol. 2:22004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Berglund L, Björling E, Oksvold P,
Fagerberg L, Asplund A, Szigyarto CA, Persson A, Ottosson J,
Wernérus H, Nilsson P, et al: A genecentric Human Protein Atlas for
expression profiles based on antibodies. Mol Cell Proteomics.
7:2019–2027. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Haeggström JZ and Funk CD: Lipoxygenase
and leukotriene pathways: Biochemistry, biology, and roles in
disease. Chem Rev. 111:5866–5898. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Skrzypczakjankun E, Chorostowskawynimko J,
Selman SH and Jankun J: Lipoxygenases-A challenging problem in
enzyme inhibition and drug development. Current Enzyme Inhibition.
3:119–132. 2007. View Article : Google Scholar
|
|
31
|
Gohara A, Eltaki N, Sabry D, Murtagh D Jr,
Jankun J, Selman SH and Skrzypczak-Jankun E: Human 5-, 12- and
15-lipoxygenase-1 coexist in kidney but show opposite trends and
their balance changes in cancer. Oncol Rep. 28:1275–1282. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shureiqi I, Wu Y, Chen D, Yang XL, Guan B,
Morris JS, Yang P, Newman RA, Broaddus R, Hamilton SR, et al:
Critical Role of 15-Lipoxygenase-1 in colorectal epithelial cell
terminal differentiation and tumorigenesis. Cancer Res.
65:11486–11492. 2011. View Article : Google Scholar
|
|
33
|
Lang F, Ritter M, Gamper N, Huber S,
Fillon S, Tanneur V, Lepple-Wienhues A, Szabo I and Gulbins E: Cell
volume in the regulation of cell proliferation and apoptotic cell
death. Cell Physiol Biochem. 10:417–428. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lang F, Busch GL, Ritter M, Völkl H,
Waldegger S, Gulbins E and Häussinger D: Functional significance of
cell volume regulatory mechanisms. Physiol Rev. 78:247–306. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bortner CD and Cidlowski JA: The role of
apoptotic volume decrease and ionic homeostasis in the activation
and repression of apoptosis. Pflugers Arch. 448:313–318. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Su Z, Yang Z, Xu Y, Chen Y and Yu Q:
Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol
Cancer. 14:482015. View Article : Google Scholar : PubMed/NCBI
|