Introduction
Oxidative stress reflects a disruption in the
balance between antioxidant defences and the production of reactive
oxygen species (ROS) (1). These
ROS are produced by cell metabolism, and affect systemic and tissue
immunity, and signal transduction (2). The accumulation of ROS instigates
cell damage and death, thus contributing to neuropathology
(3). This can lead to
neurodegenerative diseases, such as Alzheimer's disease (AD) and
Parkinson's disease (4–7). One of the major symptomatic
treatments of AD is to downregulate acetylcholinesterase (AChE),
thereby increasing acetylcholine in the brain (8). The drugs employed are AChE
inhibitors, including donepezil and tacrine. However, these are
linked to a number of side effects such as diarrhea, nausea and
anorexia (9). Huperzine A (HupA),
as shown in Fig. 1A, has been
isolated from the herb Lycopodium serratum, which is widely
used in traditional Chinese medicine (10). HupA has been shown to alleviate
oxidative stress and improve cognitive function in the elderly
(11–19). Oxidative stress impairs cholinergic
neurotransmission, therefore potentially accelerating cognitive
decline (20). Thus, antioxidants
may add to therapeutic strategies to attenuate neurotoxicity,
improving neurological outcomes in neurodegenerative pathologies.
Astaxanthin (AXT; C40H52O4;
Fig. 1B) is an antioxidative
carotenoid found in a variety of species, including crustaceans,
fish, algae, yeast and bird feathers (21). The antioxidant capacity of AXT is
~1.5X that of vitamin E (22).
This study reports on previously unknown findings concerning the
synergistic antioxidative effects of combining AXT and HupA, using
a previously established screening system to characterize
therapeutic agents that can scavenge free radicals and protect
cells from tert-butyl hydroperoxide (TBHP) (23).
Materials and methods
Chemicals
HupA and AXT (>98% in purity) were purchased from
Sichuan Weikeqi Biological Technology Co., Ltd. TBHP solution was
purchased from Shanghai Macklin Biochemical Co., Ltd. Dulbecco's
modified Eagle's medium (DMEM), fetal bovine serum (FBS) and 0.25%
(w/v) trypsin solution were purchased from Gibco; Thermo Fisher
Scientific, Inc. Aβ25–35 was purchased from BIOSS. MTT
and 2,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) were
purchased from Sigma-Aldrich; Merck KGaA. Kits for superoxide
dismutase (SOD; cat no. A001-1), lactate dehydrogenase (LDH; cat
no. BC2025) and malondialdehyde (MDA; cat no. A003-1) were
purchased from Nanjing Jiancheng Bioengineering Institute Co., Ltd.
All other chemicals were of analytical grade.
Cell culture and drug treatment
PC12 cells obtained from the Shanghai Institute of
Cell Biology, Chinese Academy of Sciences were cultured in DMEM
supplemented with 10% FBS, 1% antibiotics (penicillin and
streptomycin) at 37°C in a humidified atmosphere of 95% air and 5%
CO2. HupA and AXT were dissolved in dimethyl sulfoxide
(DMSO) and diluted to 1 mM with serum-free DMEM.
TBHP-induced cell injury model
PC12 cells were seeded in 96-well plates at a
density of 5×103 cells/well for 24 h. TBHP was freshly
prepared from 30% stock solution prior to each experiment. Seeded
cells were incubated with different concentrations (25, 50, 100,
200, 300, 400 and 500 µM) of TBHP for 24 h at 37°C. Subsequently,
20 µl MTT solution (5 mg/ml) was added for 4 h at 37°C. Finally,
the supernatant was removed and the formazan crystals were
dissolved in 150 µl DMSO. The amount of formazan was measured at
570 nm in a microplate reader (BioTek Instruments, Inc.), with the
formula: Cell viability (%)=absorption value experimental
group/absorption value of control group ×100%. Cell viability in
the control cells was expressed as 100%. The protective effect of
HupA and AXT was quantified in the same conditions while cells were
incubated with different concentrations of HupA (0.625, 6.25, 62.5,
125, 250, 500 and 1,000 µM) or AXT (0.01, 0.1, 1 and 20 µM) for 24
h at 37°C. The cell viability was determined by an MTT assay as
described above.
Cells in the TBHP-induced injury model were
incubated in separate groups: Control group (culture medium), model
group (culture medium), HupA group (0.625, 6.25, 62.5, 125, 250 and
500 µM) and AXT group (0.01, 0.1, 1 and 10 µM) for 24 h at 37°C.
Subsequently, 100 µM TBHP was added to all groups, except the
control group which contained only culture medium, for 24 h.
Finally, cell viability was determined via an MTT assay.
Subsequently, the optimal effective concentrations of HupA and AXT
were selected. PC12 cells were then incubated in separate groups
with different conditions: Control group (culture medium), model
group (culture medium), HupA group (500 µM), AXT group (0.1 µM) and
HupA + AXT group (500 + 0.1 µM) for 24 h at 37°C. This was followed
by the addition of 100 µM TBHP to all groups, except the control
group, for 24 h. Finally, the cell viability was determined via an
MTT assay.
Morphological assay
The cells were incubated at 37°C in separate groups:
Control group, model group, HupA group (500 µM), AXT (0.1 µM) group
and HupA + AXT group (500 µM + 0.1 µM). After processing, the cells
were fixed by paraformaldehyde for 15 min at room temperature. The
cellular morphological changes were stained by hematoxylin-eosin
staining for 15 min at room temperature and were examined under an
inverted phase contrast microscope at a magnification of ×400
(Nikon Corporation).
LDH content in supernatant of culture
medium
PC12 cells were seeded into 6-well plates, with
2.5×105 cells/well. After treatment as described above
in the Morphological assay section, supernatant of the culture
medium was collected to evaluate the LDH content. All samples were
detected according to the manufacturer's protocols.
Measurement of ROS production
Intracellular accumulation of ROS was measured using
DCFH-DA. PC12 were seeded into 6-well plates with
2.5×105 cells/well. After treatment, DCFH-DA (10 µM) was
added at 37°C for 30 min in the dark. Thereafter, the cells were
washed twice with PBS and harvested. The intracellular ROS levels
were measured by flow cytometry. The mean fluorescence intensity
was analyzed using NovoExpress 1.2.1 data analysis software (ACEA
Biosciences). All experiments were performed at least three
times.
Total SOD (T-SOD) and MDA content
assays
After treatment, PC12 cells were harvested and
washed with ice-cold PBS twice. The T-SOD content and MDA activity
in cell homogenates were determined according to the manufacturer's
protocols.
Ah25–35-induced cell
injury
Aβ25–35 was dissolved in sterilized PBS
at a concentration of 1 mM and incubated for 7 days at 37°C for
aggregation. The solution was diluted in culture medium prior to
each experiment. PC12 cells were seeded in 96-well plates at a
density of 5×103 cells/well for 24 h. Then, cells were
incubated with different concentrations (5, 10, 20 and 40 µM) of
Aβ25–35 for 24 h at 37°C. Subsequently, MTT (20 µl)
solution (5 mg/ml) was added to each well for 4 h at 37°C. Finally,
the supernatant was removed and the formed formazan crystals were
dissolved in 150 µl DMSO, and the plate was shaken for 10 min.
Afterwards, the amount of the formazan was measured
spectrophotometrically at 570 nm.
PC12 cells were cultured on 96-well plates and
pretreated with HupA (0.625, 6.25, 62.5, 125, 250 and 500 µM) or
AXT (0.01, 0.1, 1 and 10 µM) for 24 h before the introduction of
Aβ25–35 (20 µM). Cell viability was determined using an
MTT assay. Subsequently, the optimal effective concentrations of
HupA and AXT were selected. Cells were incubated and separated in
groups with different conditions: Control group (culture medium),
model group (culture medium), HupA group (250 µM), AXT group (0.1
µM) and HupA + AXT group (250 + 0.1 µM) for 24 h at 37°C.
Subsequently, Aβ25–35 (20 µM) was added to all groups,
except the control group, for 24 h. Finally, the cell viability was
determined via an MTT assay as aforementioned.
Statistical analysis
All results were expressed as the mean ± SEM and
analyzed by SPSS 17.0 (SPSS, Inc.). The statistical significance of
the differences between groups was analyzed by one-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered
significant.
Results
HupA and AXT have protective effects
in the TBHP injury model
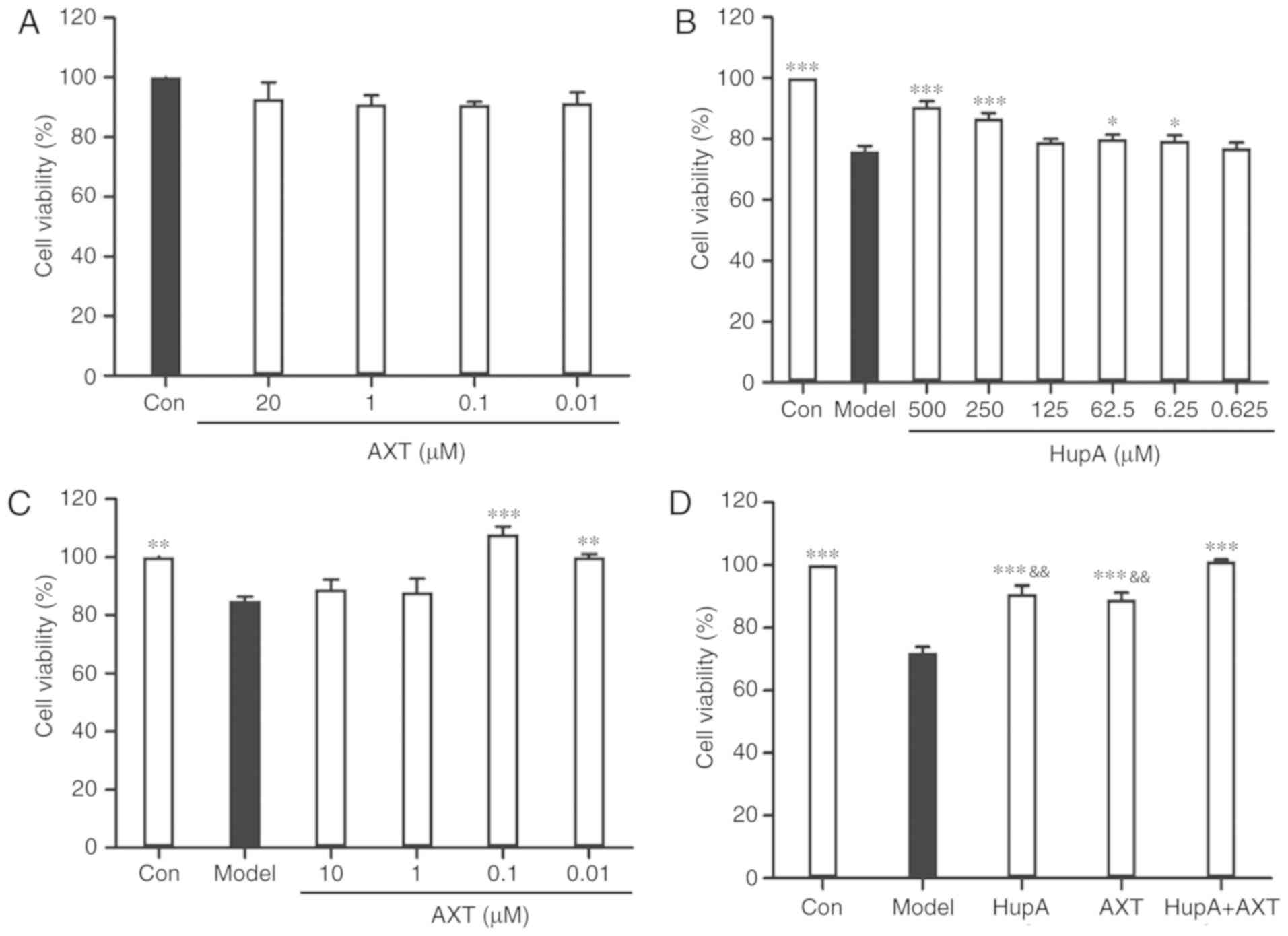
Incubation of PC12 cells with TBHP (100 µM) for 24 h
reduced cell survival to 59.35±1.22% of the control value (Fig. 1C). In subsequent experiments, PC12
cells were pretreated with HupA or AXT for 24 h, with a subsequent
addition of TBHP (100 µM) for 24 h. Exposure of PC12 cells to
different concentrations of HupA (0.625, 6.25, 62.5, 125, 250 and
500 µM) and AXT (0.01, 0.1, 1 and 20 µM) in control conditions had
no significant effect on cell survival (Figs. 1D and 2A), but a significant effect was observed
at 1,000 µM HupA (Fig. 1D). This
is due to DMSO being cytotoxic at certain concentrations. As
presented in Fig. 2B, 90.75±1.67%
(86.86±1.66%) of the cultured cells survived after 24 h treatment
with 500 µM (250 µM) HupA, which meant that 250–500 µM HupA had
protective effects against TBHP toxicity and that the most
effective concentration of HupA was 500 µM. In Fig. 2C, 101.80±2.61% of the cultured
cells survived after 24 h treatment with AXT (0.1 µM), so these
concentrations were selected for use in the subsequent experiments.
Cell viability of cells incubated with TBHP in the presence of HupA
(500 µM), AXT (0.1 µM) and HupA + AXT (500 µM + 0.1 µM) was
respectively 90.89±2.62, 89.73±2.29 and 101.34±0.51% of the control
(Fig. 2D).
The morphology of the cultured cells was assessed
using transmitted phase-contrast microscopy. The control group
showed round cell bodies with clear edges and fine dendritic
networks (Fig. 3A). Incubation
with TBHP (100 µM) for 24 h induced the shrinkage of cell bodies
and the disruption of dendritic networks, indicating that the cell
injury model was successively established by TBHP induction
(Fig. 3B). Following HupA, AXT and
HupA + AXT pretreatment, these morphological manifestations marking
cell damage in cells were alleviated, indicating that HupA, AXT and
HupA + AXT may be able to prevent cell morphology damage by TBHP
(Fig. 3C-E).
Measurement of LDH production
As shown in Table
I, the concentration of LDH in the model group (313.42±9.55
U/l) was significantly higher than in the control group
(145.57±12.11 U/l; P<0.001, n=3). However, both HupA
(229.77±9.26 U/l) and AXT (201.28±13.81 U/l) groups were lower than
the model group (P<0.001, n=3). Moreover, the HupA + AXT group
(162.37±9.04) was significantly lower than the HupA group
(P<0.01, n=3) and AXT group (P<0.05, n=3). These results
indicated that while HupA or AXT could inhibit cell death caused by
TBHP, the protective effect of HupA combined with AXT is higher
than that of either HupA or AXT alone.
 | Table I.Effects of HupA and AXT on LDH, SOD
and MDA content in PC12 cells. |
Table I.
Effects of HupA and AXT on LDH, SOD
and MDA content in PC12 cells.
| Group | LDH (U/l) | SOD (U/ml) | MDA (nmol/ml) |
|---|
| Control |
145.57±12.11c |
46.27±2.11c,f |
0.96±0.46c,e |
| Model | 313.42±9.55 | 16.29±1.80 | 3.85±0.22 |
| TBHP + HupA |
229.77±9.26c,e |
19.33±2.46a,f |
3.43±0.13f |
| TBHP + AXT |
201.28±13.81c,d |
23.09±1.65b,d |
2.38±0.19c,e |
| TBHP + HupA +
AXT |
162.37±9.04c |
29.44±2.12c |
1.67±0.11c |
HupA + AXT decreases intracellular ROS
accumulation in the TBHP injury model
As presented in Figs.
4 and 5, the intracellular ROS
level of PC12 cells measured by flow cytometry was visibly
increased after being exposed to TBHP (100 µM) for 24 h (Fig. 4C; Fig.
4A was the control group without DCFH-DA and Fig. 4B was the control group with
DCFH-DA). However, with the pretreatment of HupA, AXT and HupA +
AXT, the ROS levels were reduced from 68.07±6.56% in the model
group to 11.91±0.79% (P<0.05, n=3; Fig. 4D), 11.67±0.75% (P<0.05, n=3;
Fig. 4E) and 6.12±0.82%
(P<0.05, n=3; Fig. 4F),
respectively. The HupA + AXT group is significantly lower than the
HupA (P<0.05, n=3) and AXT group (P<0.05, n=3). This further
demonstrated the increased protective effect of combining HupA and
AXT.
Effects of HupA and AXT on MDA content
and T-SOD activity in PC12 cells
As shown in Table
I, in the presence of TBHP, MDA levels were increased from
0.96±0.46 to 3.85±0.22 nmol/ml (P<0.001, n=3), while SOD levels
were decreased from 46.27±2.11 to 16.29±1.80 U/ml (P<0.001,
n=3). Pre-incubation with HupA significantly decreased the level of
MDA to 3.43±0.13 nmol/ml and increased the activity of SOD to 19.33
U/ml (P<0.05, n=3). Pre-incubation with AXT significantly
decreased the level of MDA to 2.38±0.19 (P<0.001, n=3) and
increased the activity of SOD to 23.09±1.65 (P<0.01, n=3). The
MDA level of HupA + AXT group (1.67±0.11) was significantly lower
than the HupA (P<0.001, n=3) and AXT group (P<0.01, n=3). The
SOD level of the HupA + AXT group (29.44±2.12) was significantly
higher than the HupA (P<0.001, n=3) and AXT group (P<0.01,
n=3), thus indicating again that HupA and AXT in combination are
more effective than when used alone.
HupA and AXT decrease
Aβ25–35-induced cytotoxicity
As shown in Fig.
6A, the cell viability of PC12 cells after treatment with 5,
10, 20 and 40 µM of Aβ25–35 was 89.36±3.26, 66.51±2.26,
55.29±0.46 and 46.94±2.32% of the control groups, respectively
(n=5). Based on the results above, the concentration of 20 µM
Aβ25–35 was used for further studies. After pretreatment
with HupA (250 µM) and AXT (0.1 µM) cell viability was 79.85±1.27
and 71.89±1.97%, (Fig. 6B and C),
respectively. Although there was no statistical difference in
viability between the 250 and 500 µM HupA group (Fig. 6B), the cell viability in the 250 µM
group was still higher than in the 500 µM group. So, 250 µM was
chosen as the optimal dose in the Aβ25–35-induced PC12
cell model study. To determine whether the combination of HupA and
AXT had a better neuroprotective effect than HupA or AXT alone, the
optimal effective concentrations of HupA (250 µM) and AXT (0.1 µM)
were selected. Fig. 6D showed that
pretreatment with HupA, AXT and HupA + AXT increased the cell
viability to 78.09±1.84, 72.17±1.21 and 86.21±1.51% (n=5).
Discussion
Oxidative stress contributes to the progression of
neurodegeneration by increasing neuronal cell death (20). Therefore, the elimination of excess
ROS or an increase in the capacity of endogenous antioxidative
activity may provide a rational strategy in the treatment of
neurodegenerative diseases (24).
The pathogenesis of neurodegenerative diseases involves multiple
mechanisms, so a combination of two or more drugs with different
targets may be advantageous over single drug treatments (25,26).
Pre-treatment with HupA and AXT significantly
reduced cell death in the TBHP injury model. Furthermore, combining
HupA and AXT was more effective compared with the administration of
a single drug. However, there is a potential limitation of the
study. In Fig. 2C, the optimal
dose of AXT combined with TBHP was determined to be 0.1 µM, with a
cell viability of 101.80±2.61%, but in Fig. 2D, the cell viability was
89.73±2.29% following treatment with 0.1 µM AXT + TBHP. This shows
a discrepancy between the AXT treatment groups despite using the
same optimal dose (0.1 µM). Because AXT was dissolved in DMSO and
it was diluted to 1 µM with serum-free DMEM as the original liquid,
it is possible that the AXT used in Fig. 2D was partially decomposed due to
its long time in storage.
According to cell morphological analysis, TBHP can
cause a disruption to dendritic networks and the shrinkage of cell
bodies; HupA and AXT can rescue both of these impairments. Another
effect of TBHP toxicity is associated with increased levels of LDH,
which disrupts cell membrane permeability (27). Incubation with TBHP increased the
concentrations of LDH in the culture medium, whereas HupA and AXT
reduce LDH. These results demonstrated that HupA and AXT can
protect PC12 cells from oxidative damage; however, it is the
combination of both drugs that offers the higher degree of
protection.
Another mechanism by which HupA and AXT exert their
antioxidant effects may be associated with direct ROS scavenging.
Levels of intracellular ROS were determined by monitoring the
conversion of DCFH-DA to dichlorofluorescein (DCF). The fluorescent
signal of DCF was enhanced in the TBHP injury model, while this
increase in fluorescence was notably alleviated by pre-treatment
with HupA and AXT. These results suggested that the antioxidant
effects of HupA and AXT could result from the inhibition of
intracellular ROS production. Again, combining both drugs increased
the degree of antioxidative protection.
Oxidative damage caused by ROS is usually
accompanied by an increase in lipid peroxidation (28). MDA was used as an index to estimate
the degree of lipid peroxidation (29). SOD is regarded as an important
protective system to prevent damage caused by ROS (30); in the TBHP injury model the SOD
activity was decreased, whereas MDA levels were increased.
Pretreatment of cells with HupA and AXT reversed these changes,
indicating that the enhancement of SOD activity and the inhibition
of lipid peroxides may represent another antioxidant mechanism of
HupA and AXT.
In order to further verify the relationship between
oxidative stress and neurotoxicity, the degree of protection of
HupA and AXT on the Aβ25–35-treated PC12 AD cells model
was evaluated via an MTT assay. Results indicated that HupA and AXT
increased cell viability, especially when used in combination.
In summary, this study demonstrated that HupA and
AXT protected PC12 cells against TBHP-induced cell oxidative stress
and against Aβ25–35 toxicity. Combining both of these
drugs further increased the degree of cell protection. Therefore, a
therapeutic combination of HupA and AXT could be beneficial in the
treatment of AD and other neurodegenerative diseases.
Acknowledgements
Not applicable.
Funding
The present study was financially supported by
Guangzhou Traditional Chinese Medicine and Integrated Chinese and
Western Medicine Science and Technology Project (grant no.
20192A011019).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XY, HMW, HKZ and HN participated in the design of
the study and preparation of the manuscript. GYH, JZ and LNL
performed most of the experiments. CJL and ZJZ carried out the
statistical analysis. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Migdal C and Serres M: Reactive oxygen
species and oxidative stress. Med Sci (Paris). 27:405–412. 2011.(In
French). View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Byun EB, Park WY, Kim WS, Song HY, Sung NY
and Byun EH: Neuroprotective effect of Capsicum annuum var.
Abbreviatum against hydrogen peroxide-induced oxidative stress in
HT22 hippocampus cells. Biosci Biotechnol Biochem. 82:2149–2157.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sullivan PG, Rabchevsky AG, Waldmeier PC
and Springer JE: Mitochondrial permeability transition in CNS
trauma: Cause or effect of neuronal cell death? J Neurosci Res.
79:231–239. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Namioka N, Hanyu H, Hirose D, Hatanaka H,
Sato T and Shimizu S: Oxidative stress and inflammation are
associated with physical frailty in patients with Alzheimer's
disease. Geriater Gerontol Int. 17:913–918. 2017. View Article : Google Scholar
|
|
5
|
Sankhla CS: Oxidative stress and
Parkinson's disease. Neurol India. 65:269–270. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Uttara B, Singh AV, Zamboni P and Mahajan
RT: Oxidative stress and neurodegenerative diseases: A review of
upstream and downstream antioxidant therapeutic options. Curr
Neuropharmacol. 7:65–74. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wadhwa R, Gupta R and Maurya PK: Oxidative
stress and accelerated aging in neurodegenerative and
neuropsychiatric disorder. Curr Pharm Des. 24:4711–4725. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shinotoh H, Namba H, Fukushi K, Nagatsuka
S, Tanaka N, Aotsuka A, Ota T, Tanada S and Irie T: Progressive
loss of cortical acetylcholinesterase activity in association with
cognitive decline in Alzheimer's disease: A positron emission
tomography study. Ann Neurol. 48:194–200. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mimica N and Presecki P: Side effects of
approved antidementives. Psychiat Danub. 21:108–113. 2009.
|
|
10
|
Cheng DH, Ren H and Tang XC: Huperzine A,
a novel promising acetylcholinesterase inhibitor. Neuroreport.
8:97–101. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang R, Zhang HY and Tang XC: Huperzine A
attenuates cognitive dysfunction and neuronal degeneration caused
by beta-amyloid protein-(1–40) in rat. Eur J Pharmacol.
421:149–156. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mei Z, Zheng P, Tan X, Wang Y and Situ B:
Huperzine A alleviates neuroinflammation, oxidative stress and
improves cognitive function after repetitive traumatic brain
injury. Metab Brain Dis. 32:1861–1869. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu Z and Wang Y: Huperzine A attenuates
hepatic ischemia reperfusion injury via anti-oxidative and
anti-apoptotic pathways. Mol Med Rep. 10:701–706. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi Q, Fu J, Ge D, He Y, Ran J, Liu Z, Wei
J, Diao T and Lu Y: Huperzine A ameliorates cognitive deficits and
oxidative stress in the hippocampus of rats exposed to acute
hypobaric hypoxia. Neurochem Res. 37:2042–2052. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pohanka M, Zemek F, Bandouchova H and
Pikula J: Toxicological scoring of Alzheimer's disease drug
huperzine in a guinea pig model. Toxicol Mech Methods. 22:231–235.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang HY, Zheng CY, Yan H, Wang ZF, Tang
LL, Gao X and Tang XC: Potential therapeutic targets of huperzine A
for Alzheimer's disease and vascular dementia. Chem Biol Interact.
175:396–402. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao X and Tang XC: Huperzine A attenuates
mitochondrial dysfunction in beta-amyloid-treated PC12 cells by
reducing oxygen free radicals accumulation and improving
mitochondrial energy metabolism. J Neurosci Res. 83:1048–1057.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xiao XQ, Wang R and Tang XC: Huperzine A
and tacrine attenuate beta-amyloid peptide-induced oxidative
injury. J Neurosci Res. 61:564–569. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xiao XQ, Wang R, Han YF and Tang XC:
Protective effects of huperzine A on beta-amyloid(25–35) induced
oxidative injury in rat pheochromocytoma cells. Neurosci Lett.
286:155–158. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Saxena G, Singh SP, Agrawal R and Nath C:
Effect of donepezil and tacrine on oxidative stress in
intracerebral streptozotocin-induced model of dementia in mice. Eur
J Pharmacol. 581:283–289. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ambati RR, Phang SM, Ravi S and
Aswathanarayana RG: Astaxanthin: Sources, extraction, stability,
biological activities and its commercial applications-a review. Mar
Drugs. 12:128–152. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Naguib YM: Antioxidant activities of
astaxanthin and related carotenoids. J Agric Food Chem.
48:1150–1154. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang Y, Liu X, Huang J, Zhong Y, Mao Z,
Xiao H, Li M and Zhuo Y: Inhibition of p38 mitogen-activated
protein kinase phosphorylation decrease tert-butyl
hydroperoxide-induced apoptosis in human trabecular meshwork cells.
Mol Vis. 18:2127–2136. 2012.PubMed/NCBI
|
|
24
|
Brookmeyer R, Johnson E, Ziegler-Graham K
and Arrighi HM: Forecasting the global burden of Alzheimer's
disease. Alzheimers Dement. 3:186–191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu W, Liu BH, Xie CL, Xia XD and Zhang YM:
Neuroprotective effects of N-acetyl cysteine on primary hippocampus
neurons against hydrogen peroxide-induced injury are mediated via
inhibition of mitogen-activated protein kinases signal transduction
and antioxidative action. Mol Med Rep. 17:6647–6654.
2018.PubMed/NCBI
|
|
26
|
Lee JH, Park SY, Shin YW, Kim CD, Lee WS
and Hong KW: Concurrent administration of cilostazol with donepezil
effectively improves cognitive dysfunction with increased
neuroprotection after chronic cerebral hypoperfusion in rats. Brain
Res. 1185:246–255. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cachard-Chastel M, Devers S, Sicsic S,
Langlois M, Lezoualc'H F, Gardier AM and Belzung C: Prucalopride
and donepezil act synergistically to reverse scopolamine-induced
memory deficit in C57Bl/6j mice. Behav Brain Res. 187:455–461.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ma S, Liu X, Xun Q and Zhang X:
Neuroprotective effect of Ginkgolide K against H2O2-induced PC12
cell cytotoxicity by ameliorating mitochondrial dysfunction and
oxidative stress. Biol Pharm Bull. 37:217–225. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang Z, Lv Z, Shao Y, Qiu Q, Zhang W,
Duan X, Li Y and Li C: Microsomal glutathione transferase 1
attenuated ROS-induced lipid peroxidation in Apostichopus
japonicus. Dev Comp Immunol. 73:79–87. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Suzuki S, Miyachi Y, Niwa Y and Isshiki N:
Significance of reactive oxygen species in distal flap necrosis and
its salvage with liposomal SOD. Br J Plast Surg. 42:559–564. 1989.
View Article : Google Scholar : PubMed/NCBI
|