Introduction
Non-alcoholic steatohepatitis (NASH) is the advanced
form of non-alcoholic fatty liver disease (NAFLD) and characterized
by intrahepatic lipid accumulation, steatosis and inflammation
(1). NASH can further progress to
cirrhosis and even hepatocellular carcinoma (HSC). NASH also
significantly increases the risk of cardiovascular disease and
diabetes and has been identified as an increasing public health
burden (2). Inflammation and
oxidative stress serve a critical role in the progression of NASH.
Hence, intervention in inflammation and oxidative stress will
benefit NASH treatment.
Peroxisome proliferator-activated receptor γ
(PPARγ), a ligand-activated transcription factor, belongs to
nuclear receptor subfamily that is involved in the regulation of
lipid metabolism, glucose homeostasis, inflammation and cellular
growth (3). It has been reported
that in an animal model of NASH, the activation of PPARγ regulates
the polarization of the macrophages to M2 subtype, thus preventing
development of NASH (4).
Rosiglitazone, the widely investigated PPARγ agonist, has been
documented to be able to prevent NASH development (5,6).
However, the mechanism by which PPARγ prevents NASH remains to be
elucidated.
MicroRNAs (miRNAs/miRs) are a family of small
(19–25-nucleotide) endogenous noncoding RNAs that regulate gene
expression by pairing interactions with mRNAs, which can lead to
degradation or translation repression of target mRNAs. miR-21-5p
has been reported to be involved in liver lipid metabolism through
regulating a variety of targets, such as fatty acid binding protein
7, phosphate and tension homolog and SMAD family member 7 (SMAD7)
(7–9). Previous studies have also demonstrated
that miR-21-5p is highly expressed in NASH (9,10),
suggesting that miR-21-5p may be a useful target for NASH
treatment. However, the mechanism of miR-21-5p participation in
regulating NASH remains to be elucidated.
Secreted frizzled-related protein 5 (SFRP5) is an
anti-inflammatory adipokine that regulates metabolic homeostasis
via regulating the Wnt signaling pathways (11,12).
In a previous study, SFRP5 protein levels were significantly
downregulated in NASH (13).
However, the effect of SFRP5 on NASH remains unknown.
The present study revealed for the first time, to
the best of the authors' knowledge, that PPARγ downregulated the
expression of miR-21-5p, leading to the upregulation of SFRP5,
which contributed to the inhibition of hepatic inflammation and
oxidative stress of nonalcoholic steatohepatitis, suggesting that
the PPARγ/miR-21-5p//SFRP5 signaling pathway may be a promising
target for NASH treatment.
Materials and methods
Reagents and plasmids
Rosiglitazone (Rosi) was purchased from Cayman
Chemical Company. miR-21-5p mimic, inhibitor and negative control
(NC) were synthesized by Shanghai GenePharma Co., Ltd. Superoxide
dismutase (SOD) assay kit, methane dicarboxylic aldehyde (MDA)
assay kit, glutathione peroxidase (GSH) assay kit and oxidized
glutathione (GSSG) assay kit were purchased from Beijing Solarbio
Science & Technology Co., Ltd. miRNA Detection kit was
purchased from GeneCopoeia, Inc. ChIP kit was from Merck KGaA.
Dual-luciferase reporter assay system was from Promega Corporation.
Human miR-21-5p promoter region or mutations in PPARγ binding sites
were synthesized by Sangon Biotech Co., Ltd. The 3′-UTR of SFRP5
mRNA or mutant binding site was synthesized by Sangon Biotech Co.,
Ltd.
Clinical samples
A total of 20 liver tissue samples from patients
with NASH and 20 normal individuals were obtained from Banan
People's Hospital(Chongqing, China) between April 2020 and November
2020. All procedures were conducted in accordance with the ethical
rules stated in the Declaration of Helsinki and were approved by
the Ethics Committee of Banan People's Hospital (approval number
BNRMYY20200016). A written informed consent was obtained from each
subject before their participation. The clinical characteristics of
the patients are summarized in Table
I.
 | Table I.Characteristics of the study
individuals. |
Table I.
Characteristics of the study
individuals.
| Characteristic | Control (n=20) | NASH (n=20) | P-value |
|---|
| Age (years) | 43.5±3.6 | 44.2±4.3 | n.s. |
| Sex
(male/female) | 11/9 | 13/7 | n.s. |
| BMI
(kg/m2) | 22.3±2.7 | 25.8±3.5 | <0.05 |
| Fasting glucose
(mmol/l) |
5.3±1.7 |
5.9±1.2 | <0.05 |
| Fasting insulin
(mIU/l) | 10.4±2.9 | 15.3±1.8 | <0.05 |
| Triglycerides
(mmol/l) |
1.55±1.32 |
3.48±1.09 | <0.05 |
| Total cholesterol
(mmol/l) |
4.15±2.48 |
5.02±2.18 | n.s. |
| LDL-C (mmol/l) |
2.24±1.21 |
2.97±1.18 | n.s. |
| AST (IU/l) |
31.5±12.3 |
39.7±11.4 | <0.05 |
| ALT (IU/l) |
35.2±10.8 |
44.8±10.2 | <0.05 |
| Current smokers
(%) | 2.5 | 3 | n.s |
Cell culture and treatment
Liver cancer cell line HepG2 was purchased from
China Center for Type Culture Collection and the cell line was
authenticated by STR profiling. Cells were cultured in Dulbecco's
modified Eagle's medium (DMEM) with 10% fetal bovine serum (both
Gibco; Thermo Fisher Scientific, Inc.), streptomycin (100 mg/ml)
and penicillin (100 U/ml) at 37°C in a 5% CO2 humid
incubator. HepG2 cells were treated with 1 mm free fatty acid for
16 h to establish a NASH cell model and then the cells were treated
with DMSO, 10 M Rosi or 10 µm Rosi in the presence of miR-21-5p
mimic or the combination of miR-21-5p mimic and inhibitor for 24 h,
the cells were harvested for further experiment.
Mice and treatment
Male C57BL/6J mice (six weeks old) were purchased
from Beijing Huafukang Bioscience Co., Ltd. and housed in
specific-pathogen-free (SPF) laboratory. To induce NASH (14,15),
mice were fed with methionine and choline deficient (MCD) diet
(Research Diets A02082002B). Normal mice were fed with a control
diet (Research Diets A02082003B). After MCD treatment for 5 weeks,
mice were randomly divided into three groups (n=5): i) NASH + DMSO:
These mice were fed with MCD diet and a treated with vehicle saline
containing 0.1% DMSO intraperitoneally daily for 3 weeks; ii) NASH
+ Rosi: These mice were fed with MCD diet and treated with 10 mg/kg
Rosi intraperitoneally daily for 3 weeks; iii) NASH + Rosi +
miR-21-5p mimic: These mice were fed with MCD diet and were
received intraperitoneal injection of 10 mg/kg Rosi daily and were
injected via the tail vein daily with 8 mg/kg miR-21-5p mimic for 3
weeks (16,17). The experiment lasted for 8 weeks in
total and mice were euthanized by cervical dislocation following
mild (4 ml) diethyl ether (Sigma Aldrich, USA) anesthesia. The
liver tissues were removed for hematoxylin and eosin (H&E)
staining and the blood was collected from abdominal aorta for
biochemical detections. All animal experiments were performed
according to Guide for the Care and Use of Laboratory Animals 8th
edition (18) and approved by the
Animal Care and Use Committee of Banan People's Hospital (approval
no. BNRMYY20200029).
Transfections and luciferase
assays
HepG2 were seeded into 24-well plates at a density
of 1×105 cells/well. The cells were grown to 70%
confluence in 24-well plates before transfection at 37°C. Next,
cells were transfected with the pmirGLO reporter plasmid (empty
plasmid) (Promega Corporation), pmirGLO reporter plasmid containing
3′-UTR of SFRP5 mRNA, miR-21-5p mimic (50 nm;
5′-GUCUCAAUAUUAGGCUAAGCUAU-3′), miR-21-5p inhibitor (50 nm;
5′-AUAGCUUAGCCUAAUAUUGAGAC-3′), mimic negative control (NC;
5′-UUCUCCGAACGUGUCACGUUU-3′) or inhibitor NC
(5′-CAGUACUUUUGUGUAGUACAA-3′) (Shanghai GenePharma Co., Ltd), using
Lipofectamine® 2000 (Invitrogen, Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions at
room temperature. Subsequently, the transfected cells were cultured
for 24 h at 37°C in a 5% CO2 humid incubator. Then, the
luciferase activity was detected according to the instructions of
the Dual-Luciferase reporter gene assay kit (Thermo Fisher
Scientific, Inc.). Renilla luciferase activity was
normalized to firefly luciferase activity and the result was shown
as relative luciferase activity. Transfection experiments were
performed 3 times in triplicate. Data were represented as the
ratios of luciferase activities/Renilla activities.
RNA interference assays
The HepG2 cells were seeded into 6-well plates, when
the confluence of HepG2 cells was 70%, small interfering RNA
(siRNA) against PPARγ mRNA (100 nm; cat. no. sc-29455) and NC siRNA
(100 nm; scrambled siRNA, sc-37007; both Santa Cruz Biotechnology)
were transfected into HepG2 cells using Lipofectamine®
3000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C
according to the instructions of the manufacture. Subsequently, 24
h after cell transfection different treatments and examinations
were performed.
Reverse transcription-quantitative
(RT-q) PCR
Total RNAs were extracted from cells
(1×106 cells/well) or liver tissues (100 mg) with TRIzol
reagent (Thermo Fisher Scientific, Inc.). Reverse transcription of
total RNA was performed using Primescript RT reagent kit (Takara
Bio, Inc.) according to the manufacturer's protocol. Subsequently,
qPCR was performed with SYBR green qPCR Master Mix (Promega
Corporation) according to the manufacturer's protocol using the ABI
7500 system (Thermo Fisher Scientific, Inc.), and the relative mRNA
expression levels were normalized to β-actin. miRNAs were extracted
with E.Z.N.A. microRNA kit (Omega Bio-Tek, Inc.) and then the
expression of miR-21-5p was determined using All-in-One miRNA
qRT-PCR Detection kit (GeneCopoeia, Inc.) according to the
manufacturer's instructions and the relative expression level was
normalized against U6 small nuclear RNA (U6 snRNA). The PCR
reaction conditions were as follows: Initial denaturation at 95°C
for 2 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 30
sec. The experiments were performed three times. miRNA and mRNA
expression levels were quantified using the 2−ΔΔCq
method (19). Primer sequences are
listed in Table II.
| Table II.Primer sequences used for reverse
transcription-quantitative PCR. |
Table II.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Forward primer
sequence | Reverse primer
sequence |
|---|
| mouse PPARγ |
5′-GGAAGACCACTCGCATTCCTT-3′ |
5′-TCGCACTTTGGTATTCTTGGAG-3′ |
| human PPARγ |
5′-TCGAGGACACCGGAGAGG-3′ |
5′-GTGTCAACCATGGTCATTTCGTT-3′ |
| mouse
miR-21-5p |
5′-TAGCTTATCAGACTGATGTTGA-3′ | Universal adaptor
(supplied in the kit) |
| human
miR-21-5p |
5′-TAGCTTATCAGACTGATGTTGA-3′ | Universal adaptor
(supplied in the kit) |
| mouse SFRP5 |
5′-AAGTTCCCCCTGGACAACGA-3′ |
5′-AATGCGCATCTTGACCACAAA-3′ |
| human SFRP5 |
5′-CCACAAGTTCCCCCTGGACA-3′ |
5′-TGCGCATTTTGACCACAAAGTCA-3′ |
| mouse TNF-α |
5′-CAAACCACCAAGTGGAGGAG-3′ |
5′-GTGGGTGAGGAGCACGTAGT-3′ |
| human TNF-α |
5′-CCCTCCTTCAGACACCCT-3′ |
5′-GGTTGCCAGCACTTCACT-3′ |
| mouse IL-6 |
5′-AGTTGCCTTCTTGGGACTGA-3′ |
5′-TCCACGATTTCCCAGAGAAC-3′ |
| human IL-6 |
5′-CAATAACCACCCCTGACC-3′ |
5′-GCGCAGAATGAGATGAGTT-3′ |
| mouse MCP-1 |
5′-ACCTTTTCCACAACCACCT-3′ |
5′-GCATCACAGTCCGAGTCA-3′ |
| human MCP-1 |
5′-TTTTCCCCTAGCTTTCCC-3′ |
5′-GCAATTTCCCCAAGTCTCT-3′ |
| mouse β-actin |
5′-TGTTACCAACTGGGACGACA-3′ |
5′-GGGGTGTTGAAGGTCTCAAA-3′ |
| human β-actin |
5′-GTGAAGGTGACAGCAGTCGGTT-3′ |
5′-GAAGTGGGGTGGCTTTTAGGA −3′ |
| mouse U6 snRNA |
5′-ACTAAGCGGCCTGACTGAAG-3′ |
5′-GCCATTGTCCTTGTGACGTG-3′ |
| human U6 snRNA |
5′-CGCTTCGGCAGCACATATACTAA-3′ |
5′-TATGGAACGCTTCACGAATTTGC-3′ |
Western blotting
Whole proteins from cell/tissue samples were lysed
with RIPA lysis buffer (Beyotime Institute of Biotechnology).
Protein concentrations were determined using the BCA assay(Beyotime
Institute of Biotechnology). The proteins (50 µg) were separated by
12% SDS- polyacrylamide gel electrophoresis and transferred onto
PVDF membranes (EMD Millipore). Subsequently, the membranes were
blocked with 5% bull serum albumin (Beyotime Institute of
Biotechnology) at room temperature and then incubated at 4°C
overnight with primary antibody for SFRP5 (1:1,000; Abcam; cat. no.
ab230425), PPARγ (1:500; Santa Cruz Biotechnology, Inc.; cat. no.
sc-271392) or β-actin (1:2,000; Abcam; cat. no. ab8227). Following
washing with TBST (10 M Tris, 150 mm NaCl, 0.05% Tween-20), the
membranes were incubated with HRP-conjugated secondary antibody
(goat anti Rabbit IgG; Abcam; cat. no. ab205718, 1:10,000) and
incubated at 37°C for 1 h. Finally, the blots were visualized using
the enhanced chemiluminescence detection reagents (Thermo Fisher
Scientific, Inc.) The grey value of protein bands was analyzed
using ImageJ software 1.46r (National Institutes of Health).
Bioinformatics analysis
The putative PPARγ response element (PPARE) in
miR-21-5p promoter region was predicted by NUBIScan
(nubiscan.unibas.ch). The potential binding sites of miR-21-5p in
3′-UTR of human SFRP5 mRNA were predicted using the online
databases miRBase (mirbase.org),
TargetScan (targetscan.org) and miRanda
(microrna.org).
Chromatin immunoprecipitation (ChIP)
assays
ChIP assays were performed using EZ-ChIP kit (EMD
Millipore) according to the manufacturer's protocol. Briefly, HepG2
cells (1×107) were treated with 0.1% DMSO or 5 µm Rosi
at 37°C for 24 h and then harvested after fixation with 1%
formaldehyde for 10 min at room temperature to cross-link the
nuclear proteins to DNA. Then, the cells were lysed in SDS lysis
buffer at room temperature for 10 min and the chromatin was
sonicated with Ultrasonic Sonicator at 30% of maximum power 10
times for 10-sec pulses on ice, with 2 min interval. to shear the
DNA to an average length between 200 and 1,000 bp, followed by
immunoprecipitation with the antibody directed against PPARγ (Santa
Cruz Biotechnology, Inc.; cat. no. sc-271392×), taking IgG as a
negative control. The precipitated DNAs were purified and subjected
to PCR amplification that cover the PPARγ response element (PPARE)
in miR-21-5p promoter region (−276 to −127), taking ‘input’ (the
total DNA extract) as a positive control while ‘no DNA’ as a
negative control (20).
Oil Red O staining
The treated cells were fixed with 4%
paraformaldehyde for 30 min at room temperature and washed three
times with PBS. Next, cells were soaked in 60% Oil Red O stock
solution (0.25 g Oil Red O/100 ml isopropanol) diluted by distilled
water for 30 min at 25°C. The stained cells were washed using PBS
until the background became clear (21). Finally, images were captured under a
light microscope (Olympus Corporation) 10 randomly selected fields
of view were evaluated at ×200 magnification.
Hematoxylin and eosin (H&E)
staining
The mouse liver tissues were harvested and gently
cut into 0.3×0.5×0.5 cm cubes and fixed with 4% polyoxymethylene at
4°C for 24 h. The fixed liver tissues were dehydrated with gradient
ethanol solutions at 25°C. After washing with xylene two times at
25°C, 1.5 h each, the samples were embedded in paraffin. The
paraffin blocks were sectioned at 5 µm. Sections from each paraffin
block were stained with hematoxylin for 3 min and counterstained
with 1% Eosin Y for 10 min at 25°C to examine the pathologic
structures of the tissues. Histological steatosis, inflammation and
fibrosis were graded according to the NASH activity score (NAS)
(22,23) by a certified pathologist. Images
were captured by light microscopy (200× magnification; Olympus
Corporation); ≥10 randomly selected fields of view were
assessed.
Statistical analysis
Statistical analyses and graphical representations
were performed using GraphPad Prism 5.0 software (GraphPad Software
Inc). Data are presented as the mean ± standard deviation.
Comparisons among multiple groups were analyzed using one-way ANOVA
followed by Tukey's post hoc test. Comparisons between two groups
were analyzed using a paired t-test. The correlation analysis was
performed using Pearson's correlation coefficient. P<0.05 was
considered to indicate a statistically significant difference.
Results
The level of PPARγ is inversely
correlated with that of miR-21-5p in NASH in both mice and
humans
It has been reported that PPARγ and miR-21-5p are
associated with progression of NASH and have been proposed as
potential targets for NASH treatment. Hence, the relationship
between PPARγ and miR-21-5p was explored in NASH in mice and
humans. As shown Fig. 1A and B,
PPARγ was lowly expressed while miR-21-5p was highly expressed in
the liver tissue of mouse model of NASH and the human tissue sample
from patients with NASH (Fig. 1D and
E). Furthermore, there was a significantly inverse correlation
between the level of PPARγ and miR-21-5p in NASH in both mice and
humans (Fig. 1C and F).
Activation of PPARγ inhibits lipid
droplet accumulation, hepatic inflammation and oxidative stress by
downregulating miR-21-5p in vitro
To investigate whether PPARγ is involved in the
regulation of miR-21-5p, HepG2 cells were treated with the PPARγ
agonist Rosi and the expression of miR-21-5p was detected. As shown
in Fig. 2A, activation of PPARγ
with Rosi decreased miR-21-5p expression in a dose-dependent
manner. Knockdown of PPARγ by siRNA (cat. no. sc-29455; Santa Cruz
Biotechnology, Inc.), in which the silencing effect was high
(Fig. 2B), significantly attenuated
the Rosi-mediated downregulation of miR-21-5p (Fig. 2C). It is well established that lipid
droplet accumulation, hepatic inflammation and oxidative stress
serve important roles in the progression of NASH (24–26).
Therefore, the levels of lipogenesis, inflammatory cytokines and
oxidative stress were evaluated in an in vitro model of
NASH, in which the cells were treated with PPARγ agonist Rosi and
miR-21-5p mimic or miR-21-5p inhibitor. As shown in Fig. 2D, the number of lipid particles was
markedly decreased in cells treated with Rosi, in which PPARγ was
activated, relative to the cells treated with DMSO. However, the
number of lipid particles was increased in the cells treated with
miR-21-5p mimic, which the miR-21-5p was overexpressed after
transfected with miR-21-5p mimic (Fig.
2E). In addition, the expression levels of inflammatory markers
TNF-α, IL-6 and monocyte chemotactic protein (MCP)-1 (Fig. 2F) and oxidative stress (expression
levels of oxidative stress markers including MDA, SOD and GSH/GSSG
ratio; Fig. 2G-I) in NASH cells was
inhibited by activation of PPARγ, but the inhibitory effect was
attenuated by miR-21-5p mimic (Fig.
2F-I). Inhibition of miR-21-5p by miR-21-5p inhibitor
significantly enhanced the PPARγ-mediated inhibition of lipogenesis
(Fig. 2D), inflammation (Fig. 2F) and oxidative stress in NASH cells
(Fig. 2G-I). Taken together, these
results indicated that PPARγ repressed progression of NASH via
downregulation of miR-21-5p.
 | Figure 2.Activation of PPARγ inhibits lipid
droplet accumulation, hepatic inflammation and oxidative stress by
downregulating miR-21-5p. (A) HepG2 cells were treated with Rosi
(0.5, 5 and 10 µm) or control (0.1% DMSO) for 48 h and then the
expression of miR-21-5p was detected by qPCR. (B) siRNA against
PPARγ (si-PPARγ), or scrambled siRNA negative control (si-control)
was transfected into HepG2 cells and the expression of PPARγ was
detected by western blotting. (C) Following transfection with
si-control or si-PPARγ, HepG2 cells were treated with DMSO or 10 µm
Rosi for 24 h. Then the level of miR-21-5p was assayed by qPCR.
Data are expressed as means ± SD. *P<0.05, **P<0.01 vs. DMSO.
(D) HepG2 cells were treated with 1 mm free fatty acid for 16 h to
establish a NASH cell model and then the cells were treated with
DMSO, Rosi (10 µm), Rosi (10 µM) + miR-21-5p mimic (50 nm) or 10 µm
Rosi + miR-21-5p (50 nm) mimic + miR-21-5p inhibitor (50 nm) for 24
h. The effect of Rosi on lipid accumulation in NASH cells was
assayed by using oil Red O staining (magnification, ×200), the
arrows indicate lipid particles. (E) miR-21-5p mimic (50 nm) or
mimic NC (50 nm) were transfected into HepG2 cells. After 24 h, the
cells were harvested and the miR-21-5p was detected by qPCR. (F)
The inflammatory cytokines TNF-α, IL-6 and MCP-1 were detected by
qPCR. (G) The lipid peroxidation product MDA was tested using an
MDA assay kit. (H) The activity of SOD was measured using a SOD
assay kit. (I) The ratio of GSH/GSSG was evaluated. Data are
expressed as means ± SD. **P<0.01 vs. Rosi,
##P<0.01 vs. Rosi + miR-21-5p mimic. PPARγ,
peroxisome proliferator-activated receptor γ; miR, microRNA; Rosi,
rosiglitazone; qPCR, reverse transcription-quantitative PCR; siRNA,
small interfering RNA; NC, negative control; NASH, non-alcoholic
steatohepatitis; MCP, monocyte chemotactic protein; MDA, methane
dicarboxylic aldehyde; SOD, superoxide dismutase; GSH, glutathione
peroxidase; GSSG, oxidized glutathione. |
PPARγ suppresses the transcriptional
activity of miR-21-5p gene promoter
As a transcription factor, PPARγ usually regulates
gene transcription by binding to the PPARE in the promoter region
of target gene (27). The present
study found a putative PPARE ER4 (TGTCCTAATAAGGACT, −188 to −173)
in miR-21-5p promoter region by online bioinformatics analysis
(NUBIScan; Fig. 3A). To investigate
the mechanism by which PPARγ downregulates miR-21-5p, the
luciferase reporter plasmids containing wild-type (pGL3-PPARE-WT)
or mutant PPARγ (pGL3-PPARE-Mut) binding sites were constructed
(Fig. 3A) and the luciferase
reporter assays were performed. As shown in Fig. 3B, activation of PPARγ significantly
decreased the luciferase activity of construct pGL3-PPARE-WT,
whereas the luciferase activity of construct pGL3-PPARE-Mut was not
affected, indicating that PPARγ suppressed the transcriptional
activity of miR-21-5p promoter. In addition, the ChIP assay
demonstrated that PPARγ could directly bind to PPARE ER4 of the
miR-21-5p promoter region (Fig.
3C). These results reveal that PPARγ downregulated miR-21-5p by
binding to its promoter region.
 | Figure 3.PPARγ suppresses transcriptional
activity of miR-21-5p and upregulates SFRP5. (A) Diagram of the
putative wild-type PPARE ER4-WT and mutant ER4 (ER4-Mut, the
mutated bases are underlined) in miR-21-5p gene promoter region.
(B) HepG2 cells were transfected with luciferase reporter plasmids
pGL3-PPARE-WT, or pGL3-PPARE-Mut or pGL3-Basic and pRL-TK and
cultured for 12 h and then treated with control (0.1% DMSO) or 5 µm
Rosi for 24 h. Dual-luciferase reporter assays were performed. The
firefly luciferase activity was normalized to Renilla
luciferase activity and the result was shown as relative luciferase
activity. Data are expressed as mean ± SD of three assays performed
in triplicate. **P<0.01. (C) HepG2 cells were treated with 0.1%
DMSO or 5 µm Rosi for 24 h and the cells harvested for chromatin
immunoprecipitation assay. PPARγ, peroxisome proliferator-activated
receptor γ; miR, microRNA; SFRP5, secreted frizzled-related protein
5; PPARE, PPARγ response element; PPARγ, peroxisome
proliferator-activated receptor γ; WT, wild-type; Mut, mutant;
Rosi, rosiglitazone. |
PPARγ downregulates miR-21-5p while
upregulates SFRP5
miRNAs regulate gene expression by pairing
interactions with mRNAs, which can lead to degradation or
translation repression of target mRNAs (28). The online databases (TargetScan,
miRBase, miRanda) predicted SFRP5, an anti-inflammatory adipokine
that regulates progression of NASH, as a potential target gene of
miR-21-5p (Fig. 4A). To investigate
whether miR-21-5p is involved in the regulation of SFRP5, HepG2
cells were treated with miR-21-5p mimic or inhibitor. Treatment
with miR-21-5p mimic significantly reduced the mRNA and protein
levels of SFRP5, which was clearly attenuated by miR-21-5p
inhibitor (Fig. 4B and C). In
addition, SFRP5 was lowly expressed while miR-21-5p highly
expressed in human liver tissue samples from NASH patients and
there was a significant inverse correlation between SFRP5 and
miR-21-5p levels (Fig 4D).
miR-21-5p mimic notably decreased the luciferase activity of
pmir-SFRP5-UTR-WT (the luciferase reporter plasmid containing the
predicted miR-21-5p wild-type binding sites but not the binding
site mutant 3′-UTR), which was alleviated by miR-21-5p inhibitor
(Fig. 4E). These results suggested
that SFRP5 is one of the target genes of miR-21-5p. It was found
that activation of PPARγ enhanced the expression of SFRP5 and
overexpression of miR-21-5p markedly inhibited the PPARγ-induced
SFRP5 expression (Fig. 4F and G).
These results indicated that activation of PPARγ suppresses
expression of miR-21-5p and upregulates its target gene SFRP5.
 | Figure 4.PPARγ downregulates miR-21-5p but
upregulates SFRP5. (A) Diagrams of the predicted potential binding
sites of miR-21-5p in 3′-UTR of SFRP5 mRNA. HepG2 cells were
transfected with miR-21-5p mimic, inhibitor, or NC and cultured for
24 h and the (B) mRNA and (C) protein levels of SFRP5 were examined
by qPCR or western blotting. (D) The correlation between the
miR-21-5p and SFRP5 mRNA levels in 20 liver tissue samples from
NASH patients was analyzed using Pearson's correlation coefficient
(r=−0.629, P<0.01). (E) HepG2 cells were transfected with the
luciferase reporter plasmid in the presence of miR-21-5p mimic,
inhibitor, or mimic NC and inhibitor NC and cultured for 24 h. The
luciferase activities were measured by dual-luciferase reporter
assays. The firefly luciferase activity was normalized to
Renilla luciferase activity. HepG2 cells were transfected
with miR-21-5p mimic or mimic NC and cultured for 12 h and then
cells were treated with control (0.1% DMSO) or 5 µm Rosi. After
treatment for 24 h, the (F) mRNA and (G) protein levels of SFRP5
were examined by qPCR or western blotting. Data are expressed as
means ± SD. **P<0.01 vs. Rosi. PPARγ, peroxisome
proliferator-activated receptor γ; miR, microRNA; SFRP5, secreted
frizzled-related protein 5; NC, negative control; qPCR, reverse
transcription-quantitative PCR; NASH, non-alcoholic
steatohepatitis; Rosi, rosiglitazone. |
Activation of PPARγ represses the
progression of NASH by regulating the miR-21-5p/SFRP5 axis in
mice
NASH was induced in C57Bl/6 mice by the MCD diet and
this NASH model was used for evaluating the effects of the
PPARγ/miR-21-5p/SFRP5 pathway on NASH progression. Hepatic
histopathology analysis with H&E staining demonstrated that
various degrees of liver steatosis and numerous lipid droplets of
different sizes were observed in MCD-fed mice. However, these
pathological conditions were significantly alleviated by Rosi
treatment. In addition, treatment of miR-21-5p mimic, which the
miR-21-5p was overexpressed in the liver by caudal vein injection
(Fig. 5A), clearly reduced the
PPARγ-mediated inhibition of liver steatosis and lipid accumulation
in NASH model (Fig. 5B). In
addition, the levels of inflammatory cytokines and oxidative
related genes were tested in the liver tissues of mice with
MCD-induced NASH. As shown in Fig.
5C, activation of PPARγ dramatically suppressed the expression
of proinflammatory cytokines such as TNF-α, IL-6 and MCP-1. Similar
trends were observed in the levels of oxidative stress-related
genes including MDA, SOD and GSH/GSSG ratio in the liver tissue of
mice with MCD-induced NASH (Fig.
5D-F). However, overexpression of miR-21-5p markedly suppressed
PPARγ-mediated inhibition of hepatic inflammation and oxidative
stress (Fig. 5D-F). In addition,
activation of PPARγ markedly decreased miR-21-5p while increased
SFRP5 in MCD-fed mice (Fig. 5G and
H). These results imply that activation of PPARγ inhibited the
progression of NASH by manipulating the miR-21-5p/SFRP5 axis in
vivo.
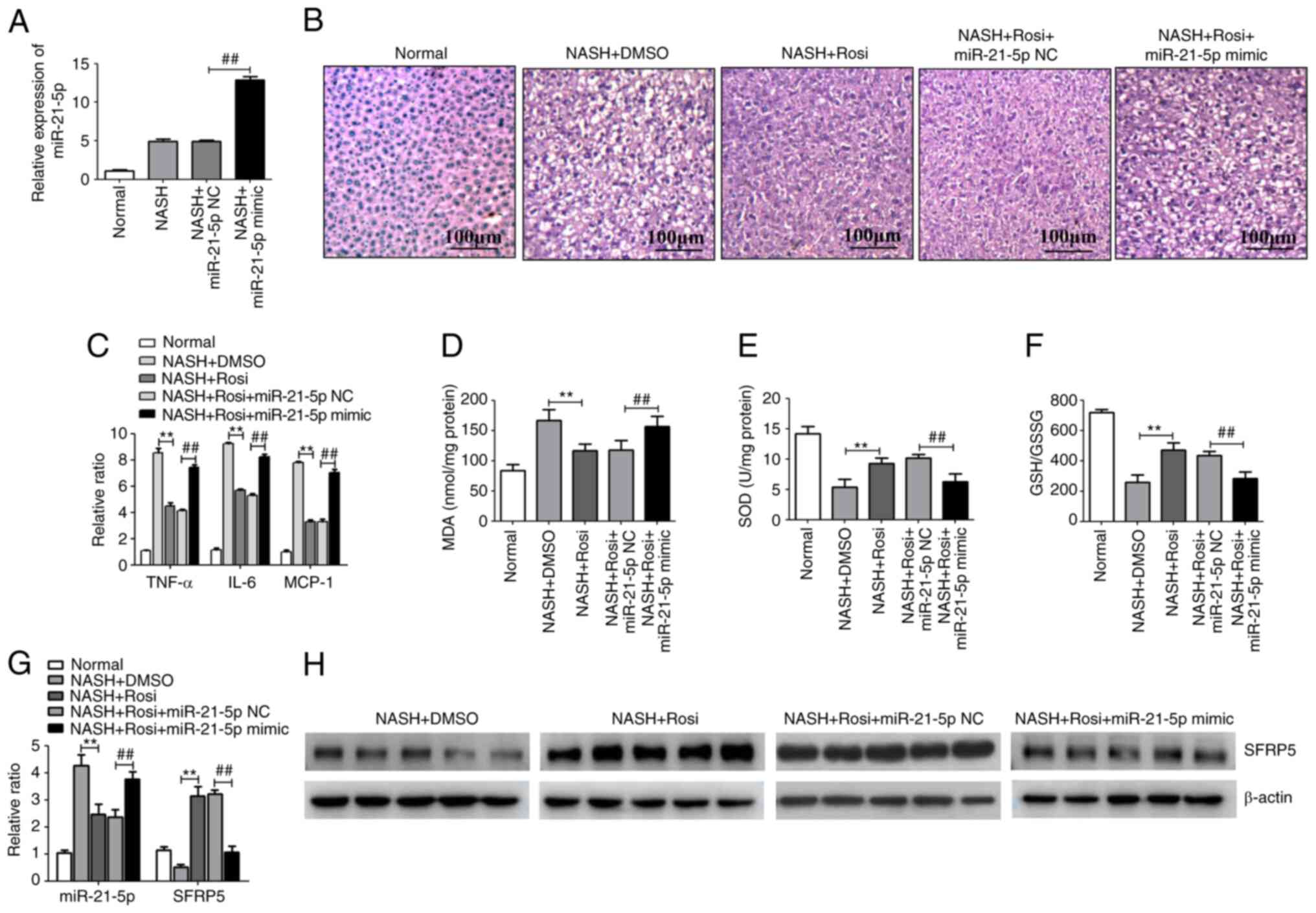 | Figure 5.Activation of PPARγ represses the
progression of NASH by regulating the miR-21-5p/SFRP5 axis in mice.
Mice were fed for 8 weeks either a methionine- and
choline-sufficient diet (Normal) or a methionine- and
choline-deficient (MCD) diet (NASH model). Model group mice were
treated with vehicle (saline containing 0.1% DMSO), Rosi (10
mg/kg), Rosi (10 mg/kg) + miR-21-5p NC (8 mg/kg), or Rosi (10
mg/kg) + miR-21-5p mimic (8 mg/kg) daily for 3 weeks (n=5). (A) The
level of miR-21-5p in liver tissues of mice which treated with
miR-21-5p NC or miR-21-5p mimic were detected by qPCR. (B) The
photomicrographs of liver sections stained with routine hematoxylin
and eosin (magnification, ×200). (C) The inflammatory cytokines
TNF-α, IL-6 and MCP-1 in the liver tissue were detected by qPCR.
(D) The lipid peroxidation product MDA was tested using an MDA
assay kit. (E) The activity of SOD was measured using a SOD assay
kit. (F) The ratio of GSH/GSSG was evaluated. Data are expressed as
means ± SD. **P<0.01 vs. DMSO, ##P<0.01 vs.
miR-21-5p NC. (G) The expression of miR-21-5p and SFRP5 in the
liver tissues was detected by qPCR. **P<0.01 vs. DMSO,
##P<0.01 vs. miR-21-5p NC. (H) The protein level of
SFRP5 in the liver tissue was detected by western blotting. PPARγ,
peroxisome proliferator-activated receptor γ; NASH, non-alcoholic
steatohepatitis; miR, microRNA; SFRP5, secreted frizzled-related
protein 5; Rosi, rosiglitazone; NC, negative control; qPCR, reverse
transcription-quantitative PCR; MCP, monocyte chemotactic protein;
MDA, methane dicarboxylic aldehyde; SOD, superoxide dismutase; GSH,
glutathione peroxidase; GSSG, oxidized glutathione; SFRP5, secreted
frizzled-related protein 5; NASH, non-alcoholic steatohepatitis;
Rosi, rosiglitazone. |
Discussion
NASH has become a global burden with an increasing
prevalence and leads to cirrhosis and HSC, which results in a high
rate of mortality per year worldwide. At present, no approved
pharmacological treatment is available for NASH, despite
development of a large variety of drugs. Currently, the detailed
mechanism of drug therapies remains to be elucidated. Thus, more
studies are required to explore the mechanisms of NASH and
potential pharmacological treatment options (29).
PPARγ is a ligand-activated transcriptional factor
that serves an important role in regulating glucose and lipid
metabolism (30,31). Rosi is a synthetic ligand of PPARγ
that is clinically used as a insulin sensitizer in the treatment of
T2DM (32). Rosi has shown
promising results in preclinical studies. Administration of Rosi
improved high-fat diet induced hepatic steatosis and lipid
metabolism through reducing hepatic Toll like receptor 4/NF-κB
expression and M1-polarized Kupffer cells (33). In MCD-diet-induced fibrosing NASH
models, Rosi may ameliorate hepatic fibrosis by activating PPARγ,
which can inhibit HSC activation and suppress TGF-β1 and CTGF
expression (34). In a biopsy
study, Rosi treatment of NASH patients for 48 weeks results in
improved hepatic steatosis, necroinflammation and ballooning
(35). In patients with NASH,
treatment with Rosi for 1 year improves steatosis and reduced
insulin resistance in most patients (36). Although PPARγ agonists have had
beneficial effects in in preclinical models of NAFLD/NASH, their
effectiveness in human pathology is limited. In addition, the
adverse effects or limited potency of PPARγ agonists also limit
their application (37–39). Therefore, the mechanism by which
PPARγ prevents NASH needs further study, which would aid
development of novel anti-NASH strategies.
As previously reported, miR-21-5p is induced in NASH
progression and recognized as a biomarker in patients with NASH
(40,41). Lack of miR-21-5p leads to
significantly less fibrogenesis, TGF-β production and downstream
signaling of the TGF-β pathway (9).
miR-21-5p ablation results in a progressive decrease in steatosis,
inflammation and lipoapoptosis in NASH (42). Therefore, miR-21-5p may be a useful
target for NASH treatment. The present study found that miR-21-5p
was markedly upregulated while PPARγ was downregulated in the liver
tissue of mouse model of NASH and the human tissue sample from
patients with NASH. There was also a significant inverse
correlation between the level of PPARγ and miR-21-5p in NASH in
mice and humans. These results suggested that there may be a
regulatory relationship between PPARγ and miR-21-5p in NASH
progress. Previous researches have shown that PPARγ, as a
transcription factor, can recognize and bind to PPARE in the
promoter region of target genes to regulate the expression of
target genes (43–46). Therefore, the present study found
some PPAREs in the miR-21-5p promoter region by bioinformatics
analysis and the reporter assay and ChIP assay demonstrated that
PPARγ could directly bind to PPARE ER4 of the miR-21-5p promoter
region, suggesting that PPARγ downregulated miR-21-5p through
binding to its promoter region.
miRNA can regulate multiple genes by targeting mRNAs
in partial sequence homology, leading to mRNA degradation or
translation repression (28).
Online database searches (miRBase, TargetScan and miRanda)
predicted some possible target genes including secreted
frizzled-related protein (SFRP)5, which belongs to the SFRP family.
Studies have found that SFRP serves a regulatory role in the Wnt
signaling pathways (47–49), specifically inhibiting the
combination of Wnt protein with its cell membrane receptors and
blocking the downstream Wnt signaling pathways by binding to
extracellular Wnt-5a or Wnt-3a (12). Kupffer cell activation and
intrahepatic inflammation can also be suppressed with recombinant
SFRP5, thus improving NASH (14).
Clinical investigations have revealed that SFRP5 protein levels are
significantly lower in NASH patients than in control subjects
(13), suggesting that SFRP5 could
have important research significance in the progress of NASH. A
previous study has shown that PPARγ directly binds to the SFRP5
promoter domain and regulates the SFRP5 in 3T3-L1 cells (50). The present study discovered that
PPARγ upregulated SFRP5 via downregulating miR-21-5p in
vitro. It also found that miR-21-5p expression was inversely
correlated with SFRP5 expression in liver tissue samples from NASH
patients and miR-21-5p directly bound to SFRP5 3′-UTR and inhibited
SFRP5 expression, indicating that SFRP5 is a target gene of
miR-21-5p. The present study also demonstrated that the
PPARγ/miR-21-5p/SFRP5 pathway is a novel mechanism underlying the
anti-NASH effects of PPARγ, indicating that PPARγ can regulate
SFRP5 in direct and indirect manners. However, more in-depth
research is needed to explore the role of the PPARγ/miR-21-5p/SFRP5
pathway so as to explore novel targets for treating NASH.
In summary, the findings of the present study
demonstrated that the activation of PPARγ attenuated hepatic
inflammation and oxidative stress in nonalcoholic steatohepatitis
via downregulating miR-21-5p. Mechanism studies found that PPARγ
could bind to PPARE ER4 of the miR-21-5p promoter region and
inhibited miR-21-5p, which directly bound to SFRP5 3′-UTR and
inhibited SFRP5 expression. These findings suggested that the
PPARγ/miR-21-5p/SFRP5 axis could improve the progression of NASH
and may represent a potential strategy for the treatment of
NASH.
Acknowledgements
Not applicable.
Funding
The study was funded by the grant from the Natural
Science Foundation of Chongqing (grant no. cstc2018jcyjAX0280).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YJ, JC and XiyZ designed the present study, analyzed
the data and wrote the manuscript. XiyZ, FD, YZ and XiaZ conducted
the experiments. FD helped to analyze the data. YJ and JC
supervised the present study and had full access to all the data.
YJ performed the manuscript submission and revised the manuscript.
YJ and JC confirm the authenticity of all the raw data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental procedures were approved by the
Ethics Committee of Banan People's Hospital. (approval no.
BNRMYY20200016). A written informed consent was obtained from each
subject before their participation.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Younossi ZM, Koenig AB, Abdelatif D, Fazel
Y, Henry L and Wymer M: Global epidemiology of nonalcoholic fatty
liver disease-meta-analytic assessment of prevalence, incidence,
and outcomes. Hepatology. 64:73–84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sanyal AJ: Past, present and future
perspectives in nonalcoholic fatty liver disease. Nat Rev
Gastroenterol Hepatol. 16:377–386. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jain MR, Giri SR, Bhoi B, Trivedi C, Rath
A, Rathod R, Ranvir R, Kadam S, Patel H, Swain P, et al: Dual PPAR
α/γ agonist saroglitazar improves liver histopathology and
biochemistry in experimental NASH models. Liver Int. 38:1084–1094.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhong X and Liu H: Honokiol attenuates
diet-induced non-alcoholic steatohepatitis by regulating macrophage
polarization through activating peroxisome proliferator-activated
receptor gamma. J Gastroenterol Hepatol. 33:524–532. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Choudhary NS, Kumar N and Duseja A:
Peroxisome proliferator-activated receptors and their agonists in
nonalcoholic fatty liver disease. J Clin Exp Hepatol. 9:731–739.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Skat-Rordam J, Ipsen DH, Lykkesfeldt J and
Tveden-Nyborg P: A role of peroxisome proliferator-activated
receptor γ in non-alcoholic fatty liver disease. Basic Clin
Pharmacol Toxicol. 124:528–537. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ahn J, Lee H, Jung CH and Ha T: Lycopene
inhibits hepatic steatosis via microRNA-21-induced downregulation
of fatty acid-binding protein 7 in mice fed a high-fat diet. Mol
Nutr Food Res. 56:1665–1674. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wei J, Feng L, Li Z, Xu G and Fan X:
MicroRNA-21 activates hepatic stellate cells via PTEN/Akt
signaling. Biomed Pharmacother. 67:387–392. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dattaroy D, Pourhoseini S, Das S, Alhasson
F, Seth RK, Nagarkatti M, Michelotti GA, Diehl AM and Chatterjee S:
Micro-RNA 21 inhibition of SMAD7 enhances fibrogenesis via
leptin-mediated NADPH oxidase in experimental and human
nonalcoholic steatohepatitis. Am J Physiol Gastrointest Liver
Physiol. 308:G298–G312. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Loyer X, Paradis V, Henique C, Vion AC,
Colnot N, Guerin CL, Devue C, On S, Scetbun J, Romain M, et al:
Liver microRNA-21 is overexpressed in non-alcoholic steatohepatitis
and contributes to the disease in experimental models by inhibiting
PPARα expression. Gut. 65:1882–1894. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ouchi N, Higuchi A, Ohashi K, Oshima Y,
Gokce N, Shibata R, Akasaki Y, Shimono A and Walsh K: Sfrp5 is an
anti-inflammatory adipokine that modulates metabolic dysfunction in
obesity. Science. 329:454–457. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Y, Rankin SA, Sinner D, Kenny AP, Krieg
PA and Zorn AM: Sfrp5 coordinates foregut specification and
morphogenesis by antagonizing both canonical and noncanonical Wnt11
signaling. Genes. 22:3050–3063. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gutierrez-Vidal R, Vega-Badillo J,
Reyes-Fermin LM, Hernandez-Perez HA, Sanchez-Munoz F, Lopez-Alvarez
GS, Larrieta-Carrasco E, Fernandez-Silva I, Mendez-Sanchez N, Tovar
AR, et al: SFRP5 hepatic expression is associated with
non-alcoholic liver disease in morbidly obese women. Ann Hepatol.
14:666–674. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen L, Zhao X, Liang G, Sun J, Lin Z, Hu
R, Chen P, Zhang Z, Zhou L and Li Y: Recombinant SFRP5 protein
significantly alleviated intrahepatic inflammation of nonalcoholic
steatohepatitis. Nutr Metab (Lond). 14:562017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rinkiko S, Pang XC, Yuan ZW, Chen SY, Zhu
YZ and Xie Y: Combinational applicaton of silybin and tangeretin
attenuates the progression of non-alcoholic steatohepatitis (NASH)
in mice via modulating lipid metabolism. Pharmacol Res.
151:1045192020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang K, Liu CY, Zhou LY, Wang JX, Wang M,
Zhao B, Zhao WK, Xu Sh, Fan LH, Zhang XJ, et al: APF lncRNA
regulates autophagy and myocardial infarction by targeting
miR-188-3p. Nat Commun. 6:67792015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang K, Long B, Zhou LY, Liu F, Zhou QY,
Liu CY, Fan YY and Li PF: CARL lncRNA inhibits anoxia-induced
mitochondrial fission and apoptosis in cardiomyocytes by impairing
miR-539-dependent PHB2 downregulation. Nat Commun. 5:35962014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the Care and Use of Laboratory Animals, 8th
edition. National Academies Press (US); Washington, DC: 2011
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu C, Liu D, Zhang Y, Lou G, Huang G, Chen
B, Shen X, Gao M, Gong W, Zhou P, et al: LXRα-mediated
downregulation of FOXM1 suppresses the proliferation of
hepatocellular carcinoma cells. Oncogene. 33:2888–2897. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhong D, Yan Z, Zeng YJ, Gao M, Wu GZ, Hu
CJ, Huang G and He FT: MicroRNA-613 represses lipogenesis in HepG2
cells by downregulating LXRα. Lipids Health Dis. 12:322013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liang W, Menke AL, Driessen A, Koek GH,
Lindeman JH, Stoop R, Havekes LM, Kleemann R and van den Hoek AM:
Establishment of a general NAFLD scoring system for rodent models
and comparison to human liver pathology. PLoS One. 9:e1159222014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takahashi Y and Fukusato T: Histopathology
of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis.
World J Gastroenterol. 20:15539–15548. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Itoh M, Ogawa Y and Suganami T: Chronic
inflammation as a molecular basis of nonalcoholic steatohepatitis:
Role of macrophages and fibroblasts in the liver. Nagoya J Med Sci.
82:391–397. 2020.PubMed/NCBI
|
|
25
|
Smeuninx B, Boslem E and Febbraio MA:
Current and future treatments in the fight against non-alcoholic
fatty liver disease. Cancers (Basel). 28:17142020. View Article : Google Scholar
|
|
26
|
Ore A and Akinloye OA: Oxidative stress
and antioxidant biomarkers in clinical and experimental models of
non-alcoholic fatty liver disease. Medicina (Kaunas). 55:262019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Penvose A, Keenan JL, Bray D, Ramlall V
and Siggers T: Comprehensive study of nuclear receptor DNA binding
provides a revised framework for understanding receptor
specificity. Nat Commun. 10:25142019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Paik JM, Golabi P, Younossi Y, Mishra A
and Younossi ZM: Changes in the global burden of chronic liver
diseases from 2012 to 2017: The growing impact of NAFLD.
Hepatology. 72:1605–1616. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hong F, Xu P and Zhai Y: The opportunities
and challenges of peroxisome proliferator-activated receptors
ligands in clinical drug discovery and development. Int J Mol Sci.
19:21892018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhu Y, Alvares K, Huang Q, Rao MS and
Reddy JK: Cloning of a new member of the peroxisome
proliferator-activated receptor gene family from mouse liver. J
Biol Chem. 268:26817–26820. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Soccio RE, Chen ER and Lazar MA:
Thiazolidinediones and the promise of insulin sensitization in type
diabetes. Cell Metab. 20:573–591. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu HM, Ni XX, Xu QY, Wang Q, Li XY and Hua
J: Regulation of lipid-induced macrophage polarization through
modulating peroxisome proliferator-activated receptor-gamma
activity a ects hepatic lipid metabolism via a Toll-like receptor
4/NF-B signaling pathway. J Gastroenterol Hepatol. 35:1998–2008.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nan YM, Fu N, Wu WJ, Liang BL, Wang RQ,
Zhao SX, Zhao JM and Yu J: Rosiglitazone prevents nutritional
fibrosis and steatohepatitis in mice. Scand J Gastroenterol.
44:358–365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Neuschwander-Tetri BA, Brunt EM, Wehmeier
KR, Oliver D and Bacon BR: Improved nonalcoholic steatohepatitis
after 48 weeks of treatment with the PPAR-gamma ligand
rosiglitazone. Hepatology. 38:1008–1017. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ratziu V, Giral P, Jacqueminet S,
Charlotte F, Hartemann-Heurtier A, Serfaty L, Podevin P, Lacorte
JM, Bernhardt C, Bruckert E, et al: Rosiglitazone for nonalcoholic
steatohepatitis: one-year results of the randomized
placebo-controlled fatty liver improvement with rosiglitazone
therapy (FLIRT) trial. Gastroenterology. 135:100–110. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Berlie HD, Kalus JS and Jaber LA:
Thiazolidinediones and the risk of edema: A meta-analysis. Diabetes
Res Clin Pract. 76:279–289. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Singh S, Loke YK and Furberg CD:
Thiazolidinediones and heart failure: A teleo-analysis. Diabetes
Care. 30:2148–2153. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Loke YK, Kwok CS and Singh S: Comparative
cardiovascular effects of thiazolidinediones: Systematic review and
meta-analysis of observational studies. BMJ. 342:d13092011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu J, Xiao Y, Wu X, Jiang L, Yang S, Ding
Z, Fang Z, Hua H, Kirby MS and Shou J: A circulating microRNA
signature as noninvasive diagnostic and prognostic biomarkers for
nonalcoholic steatohepatitis. BMC Genomics. 19:1882018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Becker PP, Rau M, Schmitt J, Malsch
C..Hammer C, Bantel H, Mullhaupt B and Geier A: Performance of
serum microRNAs −122, −192 and −21 as biomarkers in patients with
non-alcoholic steatohepatitis. PLoS One. 10:e01426612015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rodrigues PM, Afonso MB, Simao AL,
Carvalho CC, Trindade A, Duarte A, Borralho PM, Machado MV,
Cortez-Pinto H, Rodrigues CM and Castro RE: miR-21-5p ablation and
obeticholic acid ameliorate nonalcoholic steatohepatitis in mice.
Cell Death Dis. 8:e27482017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nicole W and Kay-Dietrich W: The role of
PPARs in disease. Cells. 9:23672020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ji H, Wang H, Zhang F, Li X, Xiang L and
Aiguo S: PPARγ agonist pioglitazone inhibits microglia inflammation
by blocking p38 mitogen-activated protein kinase signaling
pathways. Inflamm Res. 59:921–929. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang D, Shi L, Xin W, Xu J, Xu J, Li Q, Xu
Z, Wang J, Wang G, Yao W, et al: Activation of PPARg inhibits
pro-inflammatory cytokines production by upregulation of miR-124 in
vitro and in vivo. Biochem Biophys Res Commun. 486:726–731. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Qiu Y, Yang J, Bian S, Guozhu C and Yu J:
PPARγ suppresses the proliferation of cardiac myxoma cells through
downregulation of MEF2D in a miR-122-dependent manner. Biochem
Biophys Res Commun. 474:560–565. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu W, Ji Y, Chu H, Wang M, Yang B and Yin
C: SFRP5 mediates downregulation of the wnt5a/caveolin-1/JNK
signaling pathway. J Endocrinol. 247:263–272. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zou DP, Chen YM, Zhang LZ, Yuan XH, Zhang
YJ, Inggawati A, Nguyet PT, Gao TW and Chen J: SFRP5 inhibits
melanin synthesis of melanocytes in vitiligo by suppressing the
Wnt/β-catenin signaling. Genes Dis. 8:677–688. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tong S, Ji Q, Du Y, Zhu X, Zhu C and Zhou
Y: Sfrp5/Wnt pathway: A protective regulatory system in
atherosclerotic cardiovascular disease. J Interferon Cytokine Res.
39:472–482. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zeng J, Hu J, Lian Y, Jiang Y and Chen B:
SFRP5 is a target gene transcriptionally regulated by PPARγ in
3T3-L1 adipocytes. Gene. 641:190–195. 2018. View Article : Google Scholar : PubMed/NCBI
|















