Introduction
Acute myeloid leukemia (AML) is a type of
hematological malignancy caused by uncontrolled clonal
proliferation of hematopoietic stem cells (1,2).
The World Health Organization identifies primordial cells ≥20% as
the diagnostic standard of AML (3). In order to improve the accuracy of
acute leukemia diagnosis, morphology, immunology, cytogenetics and
molecular biology typing is commonly used, including
cytomorphology, immunology, cytogenetics and molecular biology
(4). The detection of specific
immunophenotypic markers (such as CD13, CD33, CD34 and CD117) is
important for the classification of AML (5,6).
Among them, the antigens associated with good prognosis include
myeloperoxidase, CD38 and CD19, and the antigens associated with
poor prognosis include CD56, CD7, CD123, CD34 and CD11b. Common
cytogenetic abnormalities in AML include t (15;17), 8-trisomy, t
(8;21), inv (16) or t (16;16)
and 11q23 rearrangements (7,8).
Different karyotypes affect the clinical prognosis and have
independent prognostic value (9).
Molecular genetic abnormalities are also important in improving the
prognosis and treatment of AML. Mutations in CCAAT enhancer binding
protein α, isocitrate dehydrogenase NADP+1 and nucleophosmin 1
indicate a good prognosis, while mutations in FMS-related receptor
tyrosine kinase 3, mixed lineage leukemia, ASXL transcriptional
regulator 1 and PHD finger protein 6 indicate a poor prognosis
(10,11). Finally, the abnormal expression of
oncogenes (N-ras, K-ras and Bcl-2) and suppressor genes (RB
transcriptional corepressor 1, p53 and lactate dehydrogenase) in
the serum is closely associated with the occurrence of AML
(12,13). In conclusion, the identification
and detection of pathogenic genes is an important basis for the
study of the pathogenesis, clinical treatment and prognosis of
AML.
Glycolysis is the main energy supply mode for most
tumor cells to maintain the high metabolic demand for
proliferation, which is called the ‘Warburg effect’ (14–16). Proteins related to oxidative
phosphorylation are highly enriched in the protein expression
profile of AML cells (17).
Simultaneously, the survival of AML cells is highly dependent on
mitochondrial function (18).
Inhibition of mitochondrial protein synthesis and mitochondrial DNA
replication in AML cells can effectively inhibit AML cell
proliferation (19). Compared
with most tumor cells, AML cells have their own unique metabolic
mode and their energy metabolism depends more on oxidative
phosphorylation, which is different from the traditional Warburg
effect. However, to the to the best of the authors' knowledge, its
mechanism is not clear.
The electron transport chain, also known as the
respiratory chain, is a system composed of a series of electron
carriers that transfer electrons from NADH or flavin adenine
dinucleotide to oxygen (20–22). In the process of electron
transfer, free energy is gradually released. Simultaneously, most
of the energy is stored in ATP molecules through oxidative
phosphorylation. NADH dehydrogenase is the main entrance of the
electron transport chain, catalyzing the transfer of electrons from
NADH to coenzyme Q. This reaction is the first step in the electron
transport chain and serves a vital role in energy metabolism
(23,24). NADH dehydrogenase subunits
1/2/3/4/4 L/5/6 (ND 1/2/3/4/4 L/5/6) are involved in encoding NADH
dehydrogenase. Previous studies on ND1/2/3/4/4 L/5/6 have focused
more on genetic diseases, such as Leber hereditary optic
neuropathy, Leigh syndrome and myocardial mitochondrial disease
(25,26). However, the relationship between
NADH dehydrogenase subunit (ND1/2/3/4/4L/5/6) and AML metabolism
has yet to be reported.
In the present study, analysis of The Cancer Genome
Atlas (TCGA) database revealed that the expression levels of
ND1/4/5 in AML were higher compared with those in normal samples.
Notably, a pan-cancer analysis (27) demonstrated that the expression of
ND1 was upregulated only in AML, ND2 was upregulated only in AML
and thymoma, and ND4 was upregulated only in AML and kidney
chromophobe). It was demonstrated that silencing of ND1/4/5 could
inhibit the proliferation of AML cells in cell and animal models.
Furthermore, it was revealed that oxidative phosphorylation and
energy metabolism of AML cells were decreased after silencing of
ND1/4/5. In conclusion, the present study suggested that ND1/4/5
may be involved in the regulation of oxidative phosphorylation
metabolism in AML as a potential cancer-promoting factor.
Materials and methods
Data collection
The gene expression RNA sequencing data and clinical
information of 132 patients with AML from TCGA (28) were obtained from UCSC Xena
(https://xenabrowser.net/datapages/?dat-aset=TCGA-LAML.htseq_counts.tsv).
The dataset ID in UCSC Xena was TCGA-LAML.htseq_counts.tsv. The RNA
expression data were processed by log2 (count+1)
standardization and the genes with >20% missing values (value=0)
were removed. Additionally, the clinical information of the
patients with AML was collected from TCGA.
Clinical sample collection
Between December 2019 and December 2020, the
peripheral blood of 20 healthy individuals and 20 patients with AML
was obtained from Kunming Yan'An Hospital (Kunming, China). There
were 10 males and 10 females, with a median age of 41 years (range,
8–79 years). The diagnostic criteria of AML patients followed FAB
classification (29) and NCCN
(National Comprehensive Cancer Network) AML diagnosis and treatment
guidelines (30). The present
study was approved by the ethics committee of Kunming Yan'An
Hospital (Kunming, China; approval no. 2021-03-01). All
participants provided written informed consent. The clinical
information of the patients with AML was also collected (Table I).
 | Table I.Clinical features of collected tissue
samples from AML patients |
Table I.
Clinical features of collected tissue
samples from AML patients
| Clinical
characteristics | Value (n=20) |
|---|
| Age (years) |
|
|
Median | 41 |
|
Mean | 41.8 |
|
Range | 8-79 |
| Sex (n) |
|
|
Female | 10 |
|
Male | 10 |
| FAB systems
(n) |
|
| M0 | 1 |
| M1 | 2 |
| M2 | 3 |
| M3 | 4 |
| M4 | 4 |
| M5 | 3 |
| M6 | 2 |
| M7 | 1 |
| White blood cell
(×109/l) |
|
|
Median | 38.3 |
|
Mean | 50.4 |
|
Range | 1-230 |
| Hemoglobin (g/l,
n) |
|
|
<80 | 14 |
|
≥80 | 6 |
| Platelet
(×109/l, n) |
|
|
<50 | 16 |
|
≥50 | 4 |
| Ratio of bone
marrow blasts (n) |
|
|
<60% | 9 |
|
≥60% | 11 |
Cell lines
The human acute promyelocytic leukemia cell line
(HL-60) was purchased from Cobioer Biosciences Co., Ltd. HL-60
cells were cultured in Iscove's modified Dulbecco's medium (cat.
no. 12440053; Thermo Fisher Scientific, Inc.) containing 20% fetal
bovine serum (cat. no. 10099141; Thermo Fisher Scientific, Inc.)
and 1% penicillin-streptomycin solution (cat. no. C0222; Beyotime
Institute of Biotechnology). Cells were grown with 5%
CO2 at 37°C. The HL-60 cells were cultured in hypoxia
under 95% N2 and 5% CO2. The HL-60 cells were
treated with 0.5 µM rotenone (cat. no. R8875; MilliporeSigma) or 1
µM oligomycin (cat. no. SC0366; Beyotime Biotechnology Inc.) at
37°C for 24 h. The endoribonuclease-prepared small interfering RNAs
(esiRNAs) targeting human mitochondrion (MT)-ND1, MT-ND4 or MT-ND5
(cat. nos. EHU100901, EHU101821 and EHU101561; MilliporeSigma) were
used to silence ND1, ND4 or ND5 in HL-60 cells. esiRNA targeting
enhanced green fluorescent protein (cat. no. EHUEGFP;
MilliporeSigma) was used as a control. pCMV3-C-GFPSpark (cat. no.
CV026; Sino Biological, Inc.) was used as an overexpression vector
to construct the pCMV3-ND1/4/5 recombinant plasmid, and
pCMV3-untagged (cat. no. CV011; Sino Biological, Inc.) was used as
the negative control vector. siRNA/plasmids (20 pmol) were added to
50 µl opti-MEM (cat. no. 11058021; Thermo Fisher Scientific, Inc.)
medium without serum. 1 µl Lipofectamine (cat. no. A12621; Thermo
Fisher Scientific, Inc.) was also added to 50 µl opti-MEM
serum-free medium. The two were mixed and placed at room
temperature for 20 min to form the complex. The mixture was added
to the cell suspension which was cultured at 37°C and 5%
CO2. After 48 h, other experimental steps were carried
out. HL-60 cells stably transfected with siRNA negative control
(siNC) or siRNA ND1/4/5 (siND1/4/5) were constructed. siRNA ND1:
TGATCAGGGTGAGCATCAAACTCAAACTACGCCCTGATCGGCGCACTGCGAGCAGTAGCCCAAACAATCTCATATGAAGTCACCCTAGCCATCATTCTACTATCAACATTACTAATAAGTGGCTCCTTTAACCTCTCCACCCTTATCACAACACAAGAACACCTCTGATTACTCCTGCCATCATGACCCTTGGCCATAATATGATTTATCTCCACACTAGCAGAGACCAACCGAACC.
siRNA ND4:
CAGCCACATAGCCCTCGTAGTAACAGCCATTCTCATCCAAACCCCCTGAAGCTTCACCGGCGCAGTCATTCTCATAATCGCCCACGGACTCACATCCTCATTACTATTCTGCCTAGCAAACTCAAACTACGAACGCACTCACAGTCGCATCATAATCCTCTCTCAAGGACTTCAAACTCTACTCCCACTAATAGCTTTTTGATGACTTCTAGCAAGCCTCGCTAACCTCGCCTTACCCCCCACTATTAACCTACTGGGAGAACTCTCTGTGCTAGTAACCACGTTCTCCTGATCAAA
siRNA ND5:
ACATCTGTACCCACGCCTTCTTCAAAGCCATACTATTTATGTGCTCCGGGTCCATCATCCACAACCTTAACAATGAACAAGATATTCGAAAAATAGGAGGACTACTCAAAACCATACCTCTCACTTCAACCTCCCTCACCATTGGCAGCCTAGCATTAGCAGGAATACCTTTCCTCACAGGTTTCTACTCCAAAGACCACATCATCGAAACCGCAAACATATCATACACAAACGCCTGAGCCCTATCTATTACTCTCATCGCTACCTCCCTG.
Tumor xenograft model
A total of 20 BALB/c-nu mice (male, 6 weeks old,
18–20 g) were purchased from SipeiFu Biotechnology Co., Ltd. Nude
mice were adaptively fed in a specific pathogen-free environment
for 7 days. The study protocol was ethically approved by the
Kunming Yan'An Hospital Experimental Animal Ethics Committee
(Kunming, China; approval no. 2021016). The whole animal experiment
process also followed the Animal Research: Reporting In Vivo
Experiments guidelines (31).
Mice were randomly divided into a control group (siNC) and an
experimental group (siND1/4/5) with 10 mice in each group. The cell
suspension (HL-60-siNC or HL-60-siND1/4/5) with 2×106
cells per mouse was injected into the right axillary skin of mice
after inhalation anesthesia with 2% isoflurane (cat. no.
26675-46-7; MilliporeSigma). The induction concentration of
isoflurane in mice was 4% and the maintenance concentration was 2%.
The whole experimental period was 23 days starting with the receipt
of nude mice. Mice were raised in a specific pathogen-free level
environment with suitable temperature (26–28°C), humidity (40–60%),
ventilation (10–15 times per hour) and light (10 h of light, 14 h
of no light). Each mouse had an independent sterile cage to ensure
sufficient activity space. The feed and drinking water of mice were
also pathogen-free and was used for ad libitum feeding. The health
and behavior of the mice were observed and recorded every day.
Weight loss, loss of appetite, weakness, infection of body organs
and excessive tumor volume in mice were regarded as the humane end
points. A total of 20 mice were euthanized at the terminal point of
the experiment and no mice died during the experiment. All mice
were sacrificed using intraperitoneal injection of 200 mg/kg
pelltobarbitalum solution (cat. no. P-010; MilliporeSigma) on the
18th day after injection. Before euthanasia, the mice were given
orally administrated ibuprofen (40 mg/kg; cat. no. 14883;
MilliporeSigma) with water to relieve pain. The criteria for
euthanasia were respiratory and cardiac arrest and disappearance of
nerve reflex. The long diameter (A) and short diameter (B) of the
tumor were measured every 2 days and tumor volume was calculated
according to the following formula: V = AB2/2. The
tumors were removed surgically and images were captured using a
camera (Alpha 7S III; Sony Corporation).
Differentially expressed mRNA (DEmRNA)
screening
Isolation of protein-coding genes from the RNA
expression matrix from the TCGA database was performed using the
dplyr and tidyr packages (32,33) in the R programming language
(version 3.6.2; http://www.r-project.org/). DEmRNAs were screened
using the limma (version 3.50.1) package (34) in R3.6.2 (35) according to the threshold defined
as P<0.01 and |log2foldchange|>4 (AML
group vs. normal group). DEmRNAs were visualized by a heatmap using
the pheatmap (version 1.0.12) package (36) in R3.6.2.
Pan-cancer analysis
The differential expression of ND1, ND4 and ND5
across cancers was analyzed using the Gene Expression Profiling
Interactive Analysis (http://gepia2.cancer-pku.cn/#index) online website
(37).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the cell lines or tumor
tissues using a Micro Scale RNA Isolation Kit (cat. no. AM1931;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. The total RNA concentration was measured by a micro
ultraviolet-visible spectrophotometer (NanoDrop™ One; Thermo Fisher
Scientific, Inc.). Equal amounts of RNA (2 µg) in each group were
reverse transcribed into cDNA using the High Capacity cDNA Reverse
Transcription Kit (cat. no. 4374967; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. The reverse
transcription conditions were as follows: 25°C for 10 min; 37°C for
120 min; and 85°C for 4 min. qPCR assays were performed using a
DyNAmo ColorFlash SYBR Green qPCR Kit (cat. no. F416S; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. The qPCR conditions were as follows: Pre-denaturation
(95°C for 7 min, 1 cycle); denaturation and annealing (95°C for 10
sec, 60°C for 20 sec, 40 cycles); extension (95°C for 15 sec, 60°C
for 15 sec, 1 cycle). The following primer sequences were used: ND1
forward, 5′-CAACATCGAATACGCCGCAG-3′ and reverse,
5′-AATCGGGGGTATGCTGTTCG-3′; ND4 forward, 5′-ACAAGCTCCATCTGCCTACG-3′
and reverse, 5′-GAAGCTTCAGGGGGTTTGGA-3′; ND5 forward,
5′-CACATCTGTACCCACGCCTT-3′ and reverse, 5′-TGCTATAGGCGCTTGTCAGG-3′;
and GAPDH forward, 5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′. GAPDH was the internal reference for
the RT-qPCR assay. The relative mRNA expression levels were
calculated using the 2−ΔΔCq method (38).
Reactive oxygen species (ROS)
accumulation
ROS accumulation was detected using a Reactive
Oxygen Species Assay Kit (cat. no. CA1410; Beijing Solarbio Science
& Technology Co., Ltd.). Briefly, dichlorofluorescin diacetate
(DCFH-DA) was diluted with serum-free medium at a ratio of 1:1,000
to make the final concentration 10 µmol/l. The cells were fixed at
room temperature with 95% ethanol for 30 min. After fixation, the
cells (106 cells/ml) were suspended in diluted DCFH-DA
and incubated in a cell incubator at 37°C for 20 min. The cells
were washed three times with serum-free cell culture medium. The
staining state of cells was directly observed under a fluorescence
microscope (BX53; Olympus Corporation). Quantitative analysis was
performed by flow cytometry (Model: BD Accuri C6; BD Biosciences
Inc.) with an excitation wavelength of 525 nm and an emission
wavelength of 488 nm. The result of the flow cytometry was analyzed
using BD Accuri C6 software (version: 1.0.264.21).
ELISA
ELISA was used to detect the contents of ND1, ND4
and ND5 in peripheral blood of AML patients and healthy people
according to the manufacturer's instructions of Human NADH
Dehydrogenase Subunit 1, 4 and 5 ELISA kit (cat. nos. abx576646,
abx536145 and abx152448; Abbexa, Ltd.). The optical density was
determined at OD450 nm using an microplate photometer
(Multiskan FC; Thermo Fisher Scientific, Inc.).
ATP content detection
ATP in cells and tissues was detected using an ATP
content detection kit (cat. no. BC0300; Beijing Solarbio Science
& Technology Co., Ltd.) according to the manufacturer's
instructions. ATP content was detected at a wavelength of 340 nm by
an enzyme-labeled instrument (Multiskan FC; Thermo Fisher
Scientific, Inc.).
Extracellular oxygen consumption
assay
Extracellular oxygen consumption in cells was
detected using an Extracellular Oxygen Consumption Assay kit (cat.
no. ab197243; Abcam) according to the manufacturer's instructions.
The extracellular O2 consumption signal was measured at
1.5 min intervals for 90 min by an enzyme-labeled instrument
(Multiskan FC; Thermo Fisher Scientific, Inc.). The excitation
wavelength was 380 nm and the emission wavelength was 650 nm.
Human NADH dehydrogenase (complex I)
content detection
Human NADH dehydrogenase (complex I) content in
cells and mouse tumor tissues was detected using a Human NADH
Dehydrogenase SimpleStep ELISA® Kit (cat. no. ab178011;
Abcam) according to the manufacturer's instructions. The absorbance
value was recorded at 450 nm and substituted into the standard
curve to calculate the content of complex I.
Cell Counting Kit-8 (CCK-8) assay
Cell suspension was cultured in a 96-well plate (100
µl; 5,000 cells per well). The cells were cultured at 37°C with 5%
CO2, and 10 µl CCK-8 solution (cat. no. CA1210; Beijing
Solarbio Science & Technology Co., Ltd.) was added to each
well. The cells were cultured in the incubator for 4 h. The
absorbance at 450 nm was measured by an enzyme-labeled instrument
(Multiskan FC; Thermo Fisher Scientific, Inc.).
Statistical analysis
The R programming language (version 3.6.2;
http://www.r-project.org/) and SPSS 15.0
software (SPSS, Inc.) were used for statistical analysis. All
experiments were repeated at least three times and data are
presented as the mean ± SD. A two-tailed unpaired Student's t-test
was used to determine the statistical significance of the
experimental results. One-way ANOVA with the Least Significant
Difference post hoc test was used for the comparison of multiple
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Clinical features of patients with AML
from TCGA
RNA sequencing data from a total of 132 patients
with AML were included in the present study. The average age of the
patients with AML was 53 years and only 55 patients were over 60
years old. There were 61 female patients and 71 male patients.
Among them, 80 patients succumbed to AML disease and the average
survival time was 418 days. The French-American-British
classification of patients was as follows: M0, 12 (9.09%); M1, 32
(24.24%); M2, 32 (24.24%); M3, 14 (10.61%); M4, 27 (20.45%); M5, 12
(9.09%); M6, 2 (1.52%); and M7, 1 (0.76%). Detailed clinical
information is presented in Table
II.
| Table II.Clinical and demographic data from
the 132 patients with acute myeloid leukemia in The Cancer Genome
Atlas database. |
Table II.
Clinical and demographic data from
the 132 patients with acute myeloid leukemia in The Cancer Genome
Atlas database.
| Clinical
characteristics | Value (n=132) |
|---|
| Mean age ± SD,
years | 53.27±16.36 |
| Age, ≥60 years
(n) | 55 |
| Sex, n (%) |
|
|
Female | 61 (46.21) |
|
Male | 71 (53.79) |
| Vital status, n
(%) |
|
|
Dead | 80 (60.61 |
|
Alive | 52 (39.39 |
| Mean days to death
± SD | 418.94±387.92 |
| Cytogenetics risk
category, n (%) |
|
|
Poor | 27 (20.45) |
|
Intermediate/normal | 73 (55.30) |
|
Favorable | 30 (22.73) |
|
Unknown | 2 (1.52) |
| FAB classification,
n (%) |
|
| M0 | 12 (9.09) |
| M1 | 32 (24.24) |
| M2 | 32 (24.24) |
| M3 | 14 (10.61) |
| M4 | 27 (20.45) |
| M5 | 12 (9.09) |
| M6 | 2 (1.52) |
| M7 | 1 (0.76) |
DEmRNAs
A total of 284 mRNAs were differentially expressed
between the normal and AML groups. Among them, 245 (86.27%) mRNAs
were upregulated in the AML samples, while 39 (13.73%) were
downregulated in the AML samples from TCGA database (Table SI). A heatmap was used to
visualize differentially expressed genes (Fig. 1A). The mRNA expression levels of
ND1, ND4 and ND5 were upregulated in the AML samples compared with
the normal samples (Fig. 1B;
P<0.001). Peripheral blood samples were collected from 20
healthy individuals and 20 patients with AML to verify the
differential expression of ND1, ND4 and ND5. RT-qPCR results
demonstrated that the mRNA expression levels of ND1, ND4 and ND5
were upregulated in the AML samples compared with in the normal
samples (Fig. 1C; P<0.05).
ELISA results revealed that the secretion levels of ND1, ND4 and
ND5 in the serum of patients with AML were higher than those in the
normal group (Fig. 1D;
***P<0.001, *P<0.05).
 | Figure 1.Expression levels of ND1, ND4 and ND5
are upregulated in AML. (A) Differentially expressed mRNAs in AML
based on TCGA data were visualized using a heatmap. Blue indicates
downregulated genes and red indicates upregulated genes. (B)
Scatter diagram indicating the expression levels of ND1, ND4 or ND5
in TCGA. Each scatter represents an independent sample. Normal
group vs. AML group, ***P<0.001. (C) Reverse
transcription-quantitative PCR assay indicating the mRNA expression
levels of ND1, ND4 or ND5 in normal and AML clinical samples.
Normal groups vs. AML groups, *P<0.05. (D) ELISAs indicated the
secretion level of ND1, ND4 or ND5 in serum in normal and AML
clinical samples. Normal group vs. AML group, *P<0.05,
***P<0.001. AML, acute myeloid leukemia; ND1, NADH dehydrogenase
subunit 1; ND4, NADH dehydrogenase subunit 4; ND5, NADH
dehydrogenase subunit 5; TCGA, The Cancer Genome Atlas. |
Differential expression analysis of
ND1/4/5 in pan-cancer
The differential expression of ND1, ND4 and ND5
across cancers was analyzed using the Gene Expression Profiling
Interactive Analysis online website. Compared with in normal
samples, ND1 was significantly upregulated only in AML samples. ND4
was significantly upregulated in AML and thymoma samples. ND5 was
significantly upregulated in AML and kidney chromophobe samples
(Fig. 2; P<0.001). Thus, the
upregulation of ND1, ND4 and ND5 expression is almost a unique
phenomenon in AML.
Carcinogenic effect of ND1, ND4 and
ND5
siRNA-mediated silencing of ND1, ND4, ND5 or ND1/4/5
in HL-60 cells resulted in decreased mRNA expression levels of
these genes (Fig. 3A).
Additionally, ND1, ND4 and ND5-silenced HL-60 cells showed
decreased cell viability, with the triple-KD (siND1/4/5) group
showing the largest decrease to 24.22±7.12% compared with the
control (siNC) group (Fig. 3B). A
xenograft model was used to identify the effect of ND1/4/5 on AML
growth in vivo. As shown in Fig. 3C, tumors in the siND1/4/5 group
grew slower than those in the negative control group. RT-qPCR was
performed on all tumor tissues. RT-qPCR showed that the mRNA
expression levels of ND1, ND4 and ND5 were significantly
downregulated in the siND1/4/5 group compared with the siNC group
(P<0.001; Fig. 3D). The volume
of tumors in the siNC was larger than that of siND1/4/5 groups over
time (Fig. 3E). The average
volume of tumors in the siNC or siND1/4/5 group at day 18 was
1,339.33±247.53 mm3 or 1,004.91±257.25 mm3.
Compared with siNC group, the average volume in siND1/4/5 group was
decreased (P<0.001; Fig. 3F).
The average weight of tumors in the siND1/4/5 group at 18th days
was 77.21±16.72% of that in the control (siNC) group (P<0.001;
Fig. 3G).
 | Figure 3.ND1, ND4 and ND5 promote the
proliferation of AML cells. (A) Silencing of ND1, ND4, ND5 and
ND1/4/5 in the HL-60 cell line. RT-qPCR assays indicated the gene
silencing effect in the HL-60 cell line. siND1, siND4, siND5 or
siND1/4/5 group vs. siNC group, ***P<0.001. (B) Cell Counting
Kit-8 assays demonstrated that HL-60 cells had decreased viability
after silencing of ND1, ND4, ND5 or ND1/4/5. siND1, siND4, siND5 or
siND1/4/5 group vs. siNC group, ***P<0.001. (C) A xenograft
model was used to identify the effect of ND1/4/5 on AML growth
in vivo. (D) RT-qPCR demonstrated that the mRNA expression
levels of ND1, ND4 and ND5 were downregulated in the siND1/4/5
group compared with the siNC group. siND1/4/5 group vs. siNC group,
***P<0.001. (E) Volume of tumors in the siNC and siND1/4/5
groups over time (4, 6, 8, 10, 12, 14, 16 and 18 days). (F) The
volume of tumors in the siNC group and siND1/4/5 group on the 18th
day. siND1/4/5 group vs. siNC group, ***P<0.001. (G) Weight of
tumors in the siNC group and siND1/4/5 group. siND1/4/5 group vs.
siNC group, ***P<0.001. AML, acute myeloid leukemia; ND1, NADH
dehydrogenase subunit 1; ND4, NADH dehydrogenase subunit 4; ND5,
NADH dehydrogenase subunit 5; RT-qPCR, reverse
transcription-quantitative PCR; si, small interfering RNA; siNC,
small interfering RNA negative control. |
ND1/4/5 promotes oxidative
phosphorylation in AML cells
Most tumors can survive and proliferate rapidly
under hypoxia (39). The present
study demonstrated that the viability of HL-60 cells was decreased
under hypoxia; this was 81.89±3.80% of that under normoxia
(Fig. 4A). Furthermore, when
HL-60 cells were treated with 0.5 µM rotenone or 1 µM oligomycin
(respiratory chain inhibitors) for 24 h, cell viability was
decreased to 63.56±7.38% and 75.12±2.13%, respectively, compared
with that in the control group (Fig.
4B). The present study explored whether ND1/4/5 overexpression
could alleviate the negative effects of rotenone. After
verification of the overexpression of ND1/4/5 in HL-60 cells
transfected with the PCMV3-ND1/4/5 recombinant plasmid, it was
found that it could not alleviate the inhibition of cell viability
in AML cells caused by rotenone, an inhibitor of respiratory chain
complex I (Fig. 4C). However,
silencing of ND1/4/5 downregulated the expression levels of complex
I in HL-60 cells and mouse tumor tissues (Fig. 4D). Next, the effect of ND1/4/5 on
the energy metabolism of AML cells was assessed in terms of ATP
production in HL-60 cells or tumor tissues. After silencing of
ND1/4/5, ATP production in HL-60 cells or tumor tissue was
decreased to 50.48±7.56% or 40.48±21.68% of the control (siNC),
respectively (Fig. 4E). After
silencing of ND1/4/5, the oxygen consumption of HL-60 cells was
decreased significantly to 78.56±8.34% compared with that in the
control (siNC) group (Fig. 4F).
Similarly, after silencing of ND 1/4/5, the ROS production of HL-60
cells was decreased significantly to 38.26±2.13% of that of the
control (siNC) group (Fig. 4G).
In conclusion, these data suggest that the silencing of ND1/4/5 can
inhibit the proliferation of AML cells and reduce the oxidative
phosphorylation.
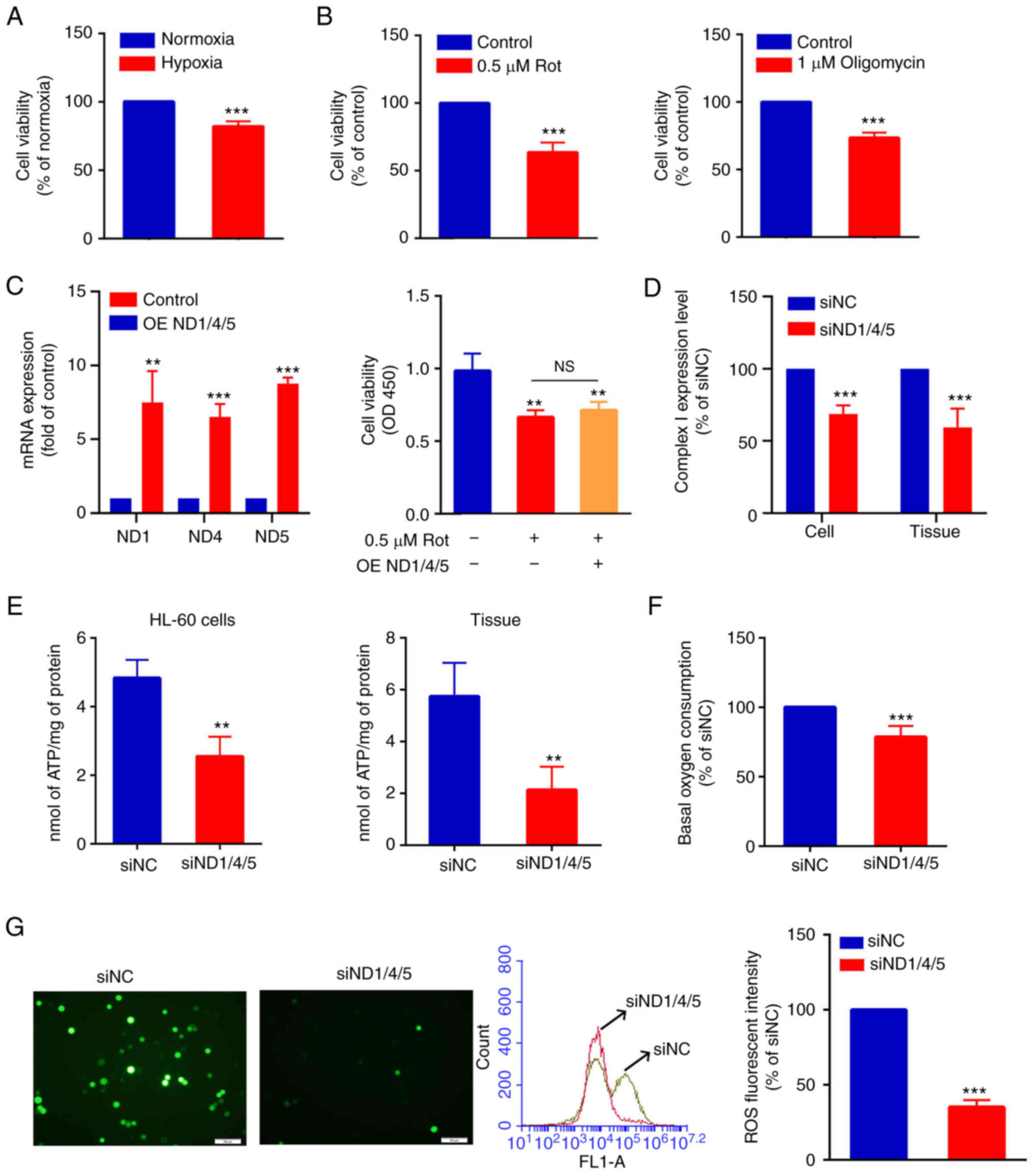 | Figure 4.ND1/4/5 promotes oxidative
phosphorylation of acute myeloid leukemia cells. (A) CCK-8 assays
showed that HL-60 cells had decreased viability under hypoxia
conditions. Normoxia group vs. hypoxia group, ***P<0.001. (B)
CCK-8 assays showed that the cell viability of HL-60 cells treated
with 0.5 µM rotenone or 1 µM oligomycin was decreased, compared
with that in the control group. Rotenone or oligomycin groups vs.
control groups, ***P<0.001. (C) Reverse
transcription-quantitative PCR assay (left) showing that the mRNA
expression levels of ND1, ND4 and ND5 were increased in the HL-60
cells transfected with the ND1/4/5 overexpression vector compared
with the negative control cells. OE ND1/4/5 group vs. control
group, **P<0.01, ***P<0.001. The CCK-8 assay (right) showed
that overexpression of ND1/4/5 could not alleviate the inhibition
of HL-60 cell viability by rotenone. Rotenone group or rotenone +
OE ND1/4/5 group vs. control group, **P<0.01. (D) ELISAs
indicated the differences in the expression levels of complex I
between the siNC and siND1/4/5 groups in HL-60 cells and tumor
tissues. siNC group vs. siND1/4/5 group, ***P<0.001. (E) ATP
production in the siNC group and siND1/4/5 group in HL-60 cells and
tumor tissues. siNC group vs. siND1/4/5 group, **P<0.01. (F)
Basal oxygen consumption in the siNC group and siND1/4/5 group in
HL-60 cells. siNC group vs. siND1/4/5 group, ***P<0.001. (G) ROS
production was detected using DCFH-DA probes by fluorescence
microscopy (magnification, ×200, left). Flow cytometry histogram
showing ROS levels in the siNC group and the siND1/4/5 group
(middle). The fluorescence intensity of ROS was measured by flow
cytometry (right). siNC group vs. siND1/4/5 group, ***P<0.001.
CCK-8, Cell Counting Kit-8; DCFH-DA, dichlorofluorescin diacetate;
ND1, NADH dehydrogenase subunit 1; ND4, NADH dehydrogenase subunit
4; ND5, NADH dehydrogenase subunit 5; OD, optical density; OE
ND1/4/5, overexpression of ND1/4/5; ROS, reactive oxygen species;
Rot, rotenone; si, small interfering RNA; siNC, small interfering
RNA negative control. |
Discussion
AML is one of the most common types of leukemia,
with a 5-year overall survival rate of 40–45% and <10% in young
and elderly patients, respectively (40). In the present study, it was found
and verified that the expression levels of ND1/4/5 in AML were
upregulated using a pan-cancer analysis of the TCGA database. A
number of studies have reported the association between the ND1/4
and the development of other tumors. For example, Jiang et
al (41) analyzed the ND1,
ND4 and GAPDH levels in plasma and blood cells from 75 patients
with thyroid cancer, which indicated that the ratio of ND1/ND4 in
thyroid cancer was abnormally elevated. Liu et al (42) found a total of 39 gene mutations
in human esophageal cancer cells (EC9706, TE-1 and ECA109). Among
them, the mutation frequency of the mitochondria encoded cytochrome
B, ND5 and ND4 genes was the highest. Therefore, the aforementioned
data together with the abnormal upregulation of ND1/4/5 in AML
suggest that ND1/4/5 could be considered as an important factor in
the occurrence and development of AML.
The present study demonstrated that the viability of
AML cell lines in vitro was decreased under respiratory
chain inhibitor treatment and hypoxia. Most tumors can survive
under hypoxia (43). Therefore,
it is suggested that the survival of AML cells depends on oxidative
phosphorylation. The present study further demonstrated that
ND1/4/5 silencing inhibited the proliferation of AML cells in
vitro and in a nude mouse model. Human mitochondrial DNA is
involved in encoding NADH dehydrogenase (ND1/2/3/4/4L/5/6), the
expression of which can affect oxidative phosphorylation (44,45). It is suggested that the special
oxidative phosphorylation metabolism of AML is related to the
upregulation of ND1/4/5, which maintains the survival of AML cells.
Our subsequent experiments confirmed our hypothesis. ND1/4/5
silencing of AML cells directly resulted in decreased expression of
complex I. After silencing ND1/4/5, ATP production, oxygen
consumption and ROS production in AML cells were decreased.
Combined with the aforementioned results, we hypothesized that the
silencing of ND1/4/5 can inhibit the proliferation of AML cells and
reduce the oxidative phosphorylation. ND1/4/5 may be a potential
oncogene.
In recent years, there have been a number of
scientific reports on the treatment of AML by inhibiting oxidative
phosphorylation. For example, Zhang et al (46) reported that a near-infrared
fluorescent dye, IR-26, exerted targeted therapeutic effects on AML
cells although impaired oxidative phosphorylation. Baccelli et
al (17) reported that
mubritinib, a known Erb-b2 receptor tyrosine kinase 2 inhibitor,
had anticarcinogenic effects through ubiquinone-dependent
inhibition of complex I activity. Carter et al (47) reported that the small molecule
compounds IACS-010759 and ME-344 inhibited oxidative
phosphorylation by targeting the electron transfer chain, thus
inhibiting the proliferation of AML cells. In the present study,
the potential oncogene ND1/4/5 was identified, which preliminarily
confirmed that upregulation of ND1/4/5 was associated with the
oxidative phosphorylation in AML. Similar to the reported oxidative
phosphorylation inhibitors, ND1/4/5 is expected to become a novel
target for the treatment of AML.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by The Key Laboratory of Tumor
Immunological Prevention and Treatment of Yunnan Province (grant
no. 2017DG004), The Joint Special Program of Applied Basic Research
of Kunming Medical University Young Doctor Program (grant no.
202001AY070001-153) and The Kunming Health Research Project (grant
no. 2020-21-01-111).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YK and YW designed the experiment and wrote the
manuscript. CP and HG contributed to the cell experiments. YD and
XY performed the statistical and bioinformatics analysis. JW and FK
performed the transplanted tumor experiment in nude mice. YK, HG
and YW confirmed the authenticity of all the raw data. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The Ethics
Committee of The Kunming Yan'An Hospital (approval no. 2021-03-01;
Kunming, China), and all patients signed informed consent forms.
The study protocol was ethically approved by the Kunming Yan'An
Hospital Experimental Animal Ethics Committee (approval no.
2021016; Kunming, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
De Kouchkovsky I and Abdul-Hay M: Acute
myeloid leukemia: A comprehensive review and 2016 update. Blood
Cancer J. 6:e4412016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thol F and Ganser A: Treatment of relapsed
acute myeloid leukemia. Curr Treat Options Oncol. 21:662020.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Čolović N, Denčić-Fekete M, Peruničić M
and Jurišić V: Clinical characteristics and treatment outcome of
hypocellular acute myeloid leukemia based on WHO classification.
Indian J Hematol Blood Transfus. 36:59–63. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Feng L, Li Y, Li Y, Jiang Y, Wang N, Yuan
D and Fan J: Whole exome sequencing detects CHST3 mutation in
patient with acute promyelocytic leukemia: A case report. Medicine
(Baltimore). 97:e122142018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen X and Cherian S: Acute myeloid
leukemia immunophenotyping by flow cytometric analysis. Clin Lab
Med. 37:753–769. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gorczyca W, Sun ZY, Cronin W, Li X, Mau S
and Tugulea S: Immunophenotypic pattern of myeloid populations by
flow cytometry analysis. Methods Cell Biol. 103:221–266. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Matsuo T, Kuriyama K, Miyazaki Y, Yoshida
S, Tomonaga M, Emi N, Kobayashi T, Miyawaki S, Matsushima T,
Shinagawa K, et al: Japan adult leukemia study group. The
percentage of myeloperoxidase-positive blast cells is a strong
independent prognostic factor in acute myeloid leukemia, even in
the patients with normal karyotype. Leukemia. 17:1538–1543. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee YJ, Huang YT, Kim SJ, Maloy M, Tamari
R, Giralt SA, Papadopoulos EB, Jakubowski AA and Papanicolaou GA:
Adenovirus viremia in adult CD34(+) selected hematopoietic cell
transplant recipients: Low incidence and high clinical impact. Biol
Blood Marrow Transplant. 22:174–178. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang JJ, Park TS and Wan TS: Recurrent
cytogenetic abnormalities in acute myeloid leukemia. Methods Mol
Biol. 1541:223–245. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kishtagari A, Levine RL and Viny AD:
Driver mutations in acute myeloid leukemia. Curr Opin Hematol.
27:49–57. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jurisić V, Pavlović S, Colović N,
Djordjevic V, Bunjevacki V, Janković G and Colović M: Single
institute study of FLT3 mutation in acute myeloid leukemia with
near tetraploidy in Serbia. J Genet. 88:149–152. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jurisic V, Radenkovic S and Konjevic G:
The actual role of LDH as tumor marker, biochemical and clinical
aspects. Adv Exp Med Biol. 867:115–124. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kavianpour M, Ahmadzadeh A, Shahrabi S and
Saki N: Significance of oncogenes and tumor suppressor genes in AML
prognosis. Tumour Biol. 37:10041–10052. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liberti MV and Locasale JW: The warburg
effect: How does it benefit cancer cells? Trends Biochem Sci.
41:211–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vaupel P, Schmidberger H and Mayer A: The
warburg effect: Essential part of metabolic reprogramming and
central contributor to cancer progression. Int J Radiat Biol.
95:912–919. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu XD, Shao SX, Jiang HP, Cao YW, Wang YH,
Yang XC, Wang YL, Wang XS and Niu HT: Warburg effect or reverse
warburg effect? A review of cancer metabolism. Oncol Res Treat.
38:117–122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baccelli I, Gareau Y, Lehnertz B, Gingras
S, Spinella JF, Corneau S, Mayotte N, Girard S, Frechette M,
Blouin-Chagnon V, et al: Mubritinib targets the electron transport
chain complex I and reveals the landscape of OXPHOS dependency in
acute myeloid leukemia. Cancer Cell. 36:84–99. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Herst PM and Berridge MV: Cell surface
oxygen consumption: A major contributor to cellular oxygen
consumption in glycolytic cancer cell lines. Biochim Biophys Acta.
1767:170–177. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Molina JR, Sun Y, Protopopova M, Gera S,
Bandi M, Bristow C, McAfoos T, Morlacchi P, Ackroyd J, Agip ANA, et
al: An inhibitor of oxidative phosphorylation exploits cancer
vulnerability. Nat Med. 24:1036–1046. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Toth A, Meyrat A, Stoldt S, Santiago R,
Wenzel D, Jakobs S, von Ballmoos C and Ott M: Kinetic coupling of
the respiratory chain with ATP synthase, but not proton gradients,
drives ATP production in cristae membranes. Proc Natl Acad Sci USA.
117:2412–2421. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li T and Le A: Glutamine metabolism in
cancer. Adv Exp Med Biol. 1063:13–32. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
van der Bliek AM, Sedensky MM and Morgan
PG: Cell biology of the mitochondrion. Genetics. 207:843–871. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hardeland R: Melatonin and the electron
transport chain. Cell Mol Life Sci. 74:3883–3896. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tatarková Z, Kuka S, Račay P, Lehotský J,
Dobrota D, Mištuna D and Kaplán P: Effects of aging on activities
of mitochondrial electron transport chain complexes and oxidative
damage in rat heart. Physiol Res. 60:281–289. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ji Y, Liang M, Zhang J, Zhang M, Zhu J,
Meng X, Zhang S, Gao M, Zhao F, Wei QP, et al: Mitochondrial
haplotypes may modulate the phenotypic manifestation of the
LHON-associated ND1 G3460A mutation in Chinese families. J Hum
Genet. 59:134–40. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rezvani Z, Didari E, Arastehkani A,
Ghodsinejad V, Aryani O, Kamalidehghan B and Houshmand M: Fifteen
novel mutations in the mitochondrial NADH dehydrogenase subunit 1,
2, 3, 4, 4L, 5 and 6 genes from Iranian patients with Leber's
hereditary optic neuropathy (LHON). Mol Biol Rep. 40:6837–6841.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nguyen L, Martens JWM, Van Hoeck A and
Cuppen E: Pan-cancer landscape of homologous recombination
deficiency. Nat Commun. 11:55842020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Blum A, Wang P and Zenklusen JC: SnapShot:
TCGA-analyzed tumors. Cell. 173:5302018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bennett JM, Catovsky D, Daniel MT,
Flandrin G, Galton DA, Gralnick HR and Sultan C: Proposals for the
classification of the acute leukaemias. French-American-British
(FAB) co-operative group. Br J Haematol. 33:451–458. 1976.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
O'Donnell MR, Tallman MS, Abboud CN,
Altman JK, Appelbaum FR, Arber DA, Attar E, Borate U, Coutre SE,
Damon LE, et al: Acute myeloid leukemia, version 2.2013. J Natl
Compr Canc Netw. 11:1047–1055. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
du Sert N, Ahluwalia A, Alam S, Avey MT,
Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl U, Emerson M, et
al: Reporting animal research: Explanation and elaboration for the
ARRIVE guidelines 2.0. PLoS Biol. 18:e30004112020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Riemondy KA, Sheridan RM, Gillen A, Yu Y,
Bennett CG and Hesselberth JR: valr: Reproducible genome interval
analysis in R. F1000Res. 6:10252017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mangiola S, Doyle MA and Papenfuss AT:
Interfacing Seurat with the R tidy universe. Bioinformatics.
24:btab4042021. View Article : Google Scholar
|
|
34
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: Limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
R Core Team R: A language and environment
for statistical computing. R Foundation for Statistical Computing,
Vienna, Austria. 2019. https://www.R-project.org/
|
|
36
|
Hu K: Become competent in generating
RNA-Seq heat maps in one day for novices without prior R
experience. Methods Mol Biol. 2239:269–303. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res.
45W:W98–W102. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Muz B, de la Puente P, Azab F and Azab AK:
The role of hypoxia in cancer progression, angiogenesis,
metastasis, and resistance to therapy. Hypoxia (Auckl). 3:83–92.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hassan C, Afshinnekoo E, Li S, Wu S and
Mason CE: Genetic and epigenetic heterogeneity and the impact on
cancer relapse. Exp Hematol. 54:26–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jiang Z, Bahr T, Zhou C, Jin T, Chen H,
Song S, Ikeno Y, Tian H and Bai Y: Diagnostic value of circulating
cell-free mtDNA in patients with suspected thyroid cancer: ND4/ND1
ratio as a new potential plasma marker. Mitochondrion. 55:145–153.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu ZW, Guo ZJ, Chu AL, Zhang Y, Liang B,
Guo X, Chai T, Song R, Hou G and Yuan JJ: High incidence of coding
gene mutations in mitochondrial DNA in esophageal cancer. Mol Med
Rep. 16:8537–8541. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Payen VL, Brisson L, Dewhirst MW and
Sonveaux P: Common responses of tumors and wounds to hypoxia.
Cancer J. 21:75–87. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
de Bièvre C and Dujon B: Organisation of
the mitochondrial genome of Trichophyton rubrum III. DNA sequence
analysis of the NADH dehydrogenase subunits 1, 2, 3, 4, 5 and the
cytochrome b gene. Curr Genet. 35:30–35. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cardol P, Lapaille M, Minet P, Franck F,
Matagne RF and Remacle C: ND3 and ND4L subunits of mitochondrial
complex I, both nucleus encoded in Chlamydomonas reinhardtii, are
required for activity and assembly of the enzyme. Eukaryot Cell.
5:1460–1467. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang C, Liu T, Luo P, Gao L, Liao X, Ma
L, Jiang Z, Liu D, Yang Z, Jiang Q, et al: Near-infrared oxidative
phosphorylation inhibitor integrates acute myeloid
leukemia-targeted imaging and therapy. Sci Adv. 7:eabb61042021.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Carter JL, Hege K, Kalpage HA, Edwards H,
Hüttemann M, Taub JW and Ge Y: Targeting mitochondrial respiration
for the treatment of acute myeloid leukemia. Biochem Pharmacol.
182:1142532020. View Article : Google Scholar : PubMed/NCBI
|















