Introduction
Apoptosis is a physiologically essential mechanism
of the cell and is important in the development and progression of
tumors. Regulated inhibition of apoptosis preserves normal
homeostasis and tissue and organ morphogenesis. The clear strategy
for cancer therapy is to target the lesions that suppress apoptosis
in tumor cells (1). Diverse
molecular mechanisms are involved in the induction of apoptosis
(2,3). Survivin, the smallest member of the
mammalian inhibitors of the apoptosis protein (IAP) family, is
upregulated in various malignancies to protect cells from apoptosis
(4). Overexpression of survivin in
cancer overcomes the apoptotic checkpoint and favors the aberrant
progression through mitosis (5).
Gene silencing by RNA interference (RNAi) using
synthetic small interfering RNA (siRNA) or expressed stem-loop RNA
[short-hairpin RNA (shRNA) or artificial microRNA (miRNA)] has been
shown to be effective against survivin in vitro and in
vivo (6–8). However, due to the short half-life and
low in vivo transfection efficiency of synthetic siRNA, the
application of shRNA expression vectors, under the direction of RNA
polymerase III promoters such as U6 and H1, may be a powerful tool
for anticancer therapy (9,10). shRNA was significantly more potent
than siRNA at mediating knockdown, and the difference resulted from
the less efficient delivery of siRNA to the cytosol compared with
shRNA delivery to the nucleus (11). Moreover, shRNA was more effective
than the artificial miRNA in mediating gene silencing,
independently of the target sequence and experimental setting
(12).
However, the utilization of shRNA expression vectors
has been limited by the inefficient delivery technique,
particularly in vivo (13).
Currently, techniques that have been considered for gene delivery
of shRNA expression vectors include cationic lipids and liposomes,
viruses and physical methods. However, a number of aspects limit
the applicability of these methods in humans. The use of a viral
vector has been developed as a highly efficient method for gene
delivery to a variety of tissues, although it evokes specific
immune responses that may limit clinical application. Among
non-viral techniques, ultrasound-targeted microbubble destruction
(UTMD) has evolved as a new, promising tool for site-specific drug
and gene delivery in vitro and in vivo, targeting
delivery via a process called sonoporation, allowing for direct
transfer into the cells (14–16).
Significant efforts have been made to demonstrate the application
of siRNA mediated by UTMD to block gene expression in vitro
and in vivo (17–20). Wang et al (21) found that UTMD was capable of
delivering survivin siRNA into SKOV-3 cells, which inhibited
survivin expression and induced apoptosis. This technology provided
a new promising approach for siRNA delivery in vitro. In our
previous study (22),
shRNA-survivin was added to HeLa cells followed by UTMD, and the
downregulation of survivin expression was assessed. We suggested
that this technique effectively inhibited the expression of the
target gene and induced cell apoptosis. Moreover, in a previous
in vitro experimental study (23), we attempted to solve an important
problem arising from the application of the non-viral gene transfer
system of UTMD (combination of ultrasound exposure and liposome
microbubbles) and PEI, particularly in the transfection of shRNA
targeting survivin. However, the UTMD technique for the delivery of
shRNA in vitro had not yet been optimized, and such methods
of apoptosis induction and the efficacy of using UTMD technique and
shRNA expression vectors had not been studied.
In the present study, we investigated whether or not
the different shRNAs targeting survivin were capable of being
transfected by the UTMD technique. Notably, UTMD para- meters for
the delivery system of shRNA in vitro were optimized.
Furthermore, we investigated the effects of gene inhibition and
apoptosis induction, which was not performed previously. The
results revealed that the optimal irradiation parameters obtained
higher transfection efficiency and did not affect the integrity of
plasmid DNA. UTMD mediated survivin gene mRNA and protein knockdown
significantly, and caused marked cell apoptosis.
Materials and methods
Cell culture
Human cervical cancer cell lines (HeLa) were
obtained from the American Type Culture Collection (ATCC) and
cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented
with 10% heat-inactivated fetal bovine serum (Invitrogen
Biotechnology, Shanghai, China). Cultures were grown at 37°C in a
humidified atmosphere, containing 5% CO2.
Construction of shRNA expression vectors
targeting survivin
DNA template oligonucleotides corresponding to the
human survivin gene (GenBank accession no. NM_001168) were designed
and synthesized as in our previous study [23]: survivin-shRNA1
(sense, 5′-GATCCGGAACTGGCCCTT CTTGGAGTTCAAGAGACTCCAAGAAGGGCCAGTTCT
TTTTTGGAAG-3′); survivin-shRNA2 (sense, 5′-GATCCG
ACTGGACAAGAGAAAGAGCCTTCAAGAGAGGCTCTT TCTCTGTCCAGTTTTTTTGGAAG-3′);
survivin-shRNA3 (sense, 5′-GATCCGGGACCACCGCATCTCTACATTCAA
GAGATGTAGAGATGCGGTGGTCCTTTTTTGGAAG-3′). These double strand
oligonucleotides were subcloned into a linearized U6
promoter-driven pSIREN-DNR-DsRed-Express vector (BD Biosciences
Clontech, USA) at the BamHI and EcoRI sites. The
selected reconstructed plasmids for transfection were extracted and
purified using a Qiaquick kit (Qiagen, Crawley, UK). Those specific
recombinant shRNA vectors were termed pSIREN/S1, pSIREN/S2 and
pSIREN/S3, respectively. Similarly, a non-specific control vector
was constructed (pSIREN/con). Transient transfections were
performed using Lipofectamine™ 2000 (Invitrogen Biotechnology,
Shanghai, China), as described previously (24,25).
Optimization of survivin-shRNA with the
UTMD technique
Briefly, cells were harvested by 0.25% trypsin
digestion after reaching 80-90% confluence and were resuspended in
DMEM media (300 μl/well) at a concentration of 1×107
cell/ml (26). SonoVue microbubbles
(Bracco, Milan, Italy) were used to promote cavitation and were
reconstituted in saline solution at a concentration of 25–30
particles/cell, and slowly added to the cell suspension prior to
ultrasound exposure (25). The UTMD
experiments were performed in an exposure tank and the transducer
(Accusonic, Metron Medical Australia Pty., Ltd.) was held in a
positioning device at the base of the tank, as previously described
(27).
Experiments were divided into the groups: negative
control group, Lipofectamine-transfected groups (pSIREN/CON+L,
pSIREN/S1+L, pSIREN/S2+L, pSIREN/S3+L) and UTMD-treated groups
(pSIREN/S1+UTMD, pSIREN/S2+UTMD, pSIREN/S3+UTMD).
The study was approved by the Ethics Committee of
The Third Affiliated Hospital of Guangzhou Medical College,
Guangzhou, China.
Optimization of the UTMD protocol
Plasmid (final concentration 25 μg/ml) was added to
the cell suspension prior to ultrasound exposure. The cells were
exposed to various ultrasound intensities (0.4, 1.0, 1.6 and 2.2
W/cm2) for various durations (1 and 3 min), with a 10 or
20% of duty cycle (DC) and frequency of 1 MHz, n=6 for each
parameter.
Analysis of transfection efficiency
The transfected cell expression of red fluorescence
protein (RFP) was visualized by an inverse fluorescence microscope
(Olympus IX71, Japan) after 48 h (28). The cells were then harvested,
collected and resuspended in phosphate-buffered saline (PBS).
Approximately 1×105 cells were obtained from each sample
for transfection efficiency analysis, with a 488-nm wavelength
excitation light and a 585±42 nm wavelength emission light to
detect the red fluorescence using flow cytometry (FACSCalibur,
Becton-Dickinson, USA). The transfection efficiency was assessed as
the number of cells expressing RFP per total number of surviving
cells.
Stability of plasmid DNA following
ultrasound exposure
Plasmid DNA (100 μl) or SonoVue/DNA complexes were
added into 6 wells of 96-well culture plates, as described
previously (15), and exposed to
ultrasound irradiation (0.4, 1.0, 1.6 and 2.2 W/cm2; DC,
20%; exposure time, 3 min) with or without the addition of SonoVue
microbubbles. Gel electrophoresis analysis was performed
immediately after ultrasound irradiation.
Reverse transcription PCR (RT-PCR)
Cells were harvested 48 h following transfection,
and total RNA was extracted with TRIzol reagent (Invitrogen)
according to the manufacturer's instructions. The RNA was
reverse-transcribed into cDNA with a reverse transcription kit
(Promega, Madison, WI, USA). Specific primers were designed on the
basis of human cDNA sequences using Primer Premier 5.0 software
(Premier Biosoft International, Palo Alto, CA, USA) and synthesized
by Invitrogen. The sequences of the primers were: forward,
5′-ACCACCGCATCTCTACATTC-3′; reverse, 5′-AGTCTG GCTCGTTCTCAGTG-3′;
and the predicted product was 113 bp. GAPDH was used as an internal
standard and its mRNA was amplified with primers: forward,
5′-GAAGGTGAA GGTCGGAGTC-3′; reverse, 5′-GAAGATGGTGATGGG ATTTC-3′;
and the predicted product was 226 bp. PCR amplification of the
sequences was performed and the products obtained were
electrophoresed through a 1% agarose gel with ethidium bromide. The
intensity of survivin and GAPDH were evaluated using Quantity One
software (Bio-Rad, Hercules, CA, USA). The relative expression
levels of survivin were expressed as the ratio of
survivin/GAPDH.
Western blotting
The cells were washed twice with cold PBS prior to
being lysed in cell lysis buffer. Cell extracts were separated by
10% sodium dodecyl sulfate polyacrylamide gel electrophoresis
(SDS-PAGE), and transferred onto nitrocellulose membranes
(Millipore, Billerica, MA, USA). Immunoblot analysis was performed
according to standard protocol (24). The blots were developed using
enhanced chemiluminescence. The relative levels of survivin were
expressed as the ratio of survivin/β-actin.
DNA ladder formation
DNA was extracted from detached and adherent cells,
and was subjected to electrophoresis using agarose gel as
previously described (15).
Flow cytometry assays of apoptosis
Cells were assessed by flow cytometry (FACSCalibur,
BectonDickinson) following the double staining of cells with
annexin V and 7-AAD (Jinmei Biotech Co. Ltd., Wuhan, China). Early
apoptotic cells were stained with FITC-labelled annexin-V alone,
whereas necrotic and late apoptotic cells were stained with annexin
V and 7-AAD (29). Each assay was
performed three times.
Flow cytometric analysis of the cell
cycle
A total of 48 h after transfection, HeLa cells were
collected by trypsinization, pooled with non-attached cells, and
fixed in 70% ethanol on ice for 30 min. Fixed cells were pelleted
by centrifugation and suspended in 10 mg/ml propidium iodide, 100
mg/ml RNase A and 0.05% Triton X-100 in PBS (pH 7.4), and finally
stained with 20 mg/ml propidium iodide (PI; 300 ml) for 20 min.
Samples were analyzed for DNA content by flow cytometry. The
samples were assayed in triplicate, and the fraction of every cell
cycle phase was calculated.
Statistical analysis
Statistical analysis was performed using the SPSS
13.0 package (SPSS, Chicago, IL, USA). The variables were shown as
the mean ± standard deviation (SD). P<0.05 was considered to
indicate a statistically significant difference.
Results
Optimization of the UTMD-mediated
delivery system in vitro
First, we investigated whether the UTMD technique
was suitable for the delivery of shRNA into cells. As shown in
Fig. 1A, transfection efficiency
was affected by transfection parameters, such as ultrasound
intensity and exposure time. The optimal conditions of ultrasound
exposure achieved the highest transfection efficiency (Fig. 1A-D). Moreover, our results showed
that the appearance of DNA fragments was correlated with the
transfection parameters (Fig. 1C).
No significant effect was noted on the structure of vectors under
suitable exposure conditions. The optimized parameters (1.0
W/cm2 for 3 min with 20% DC pulse mode) were used in the
subsequent UTMD-treated groups.
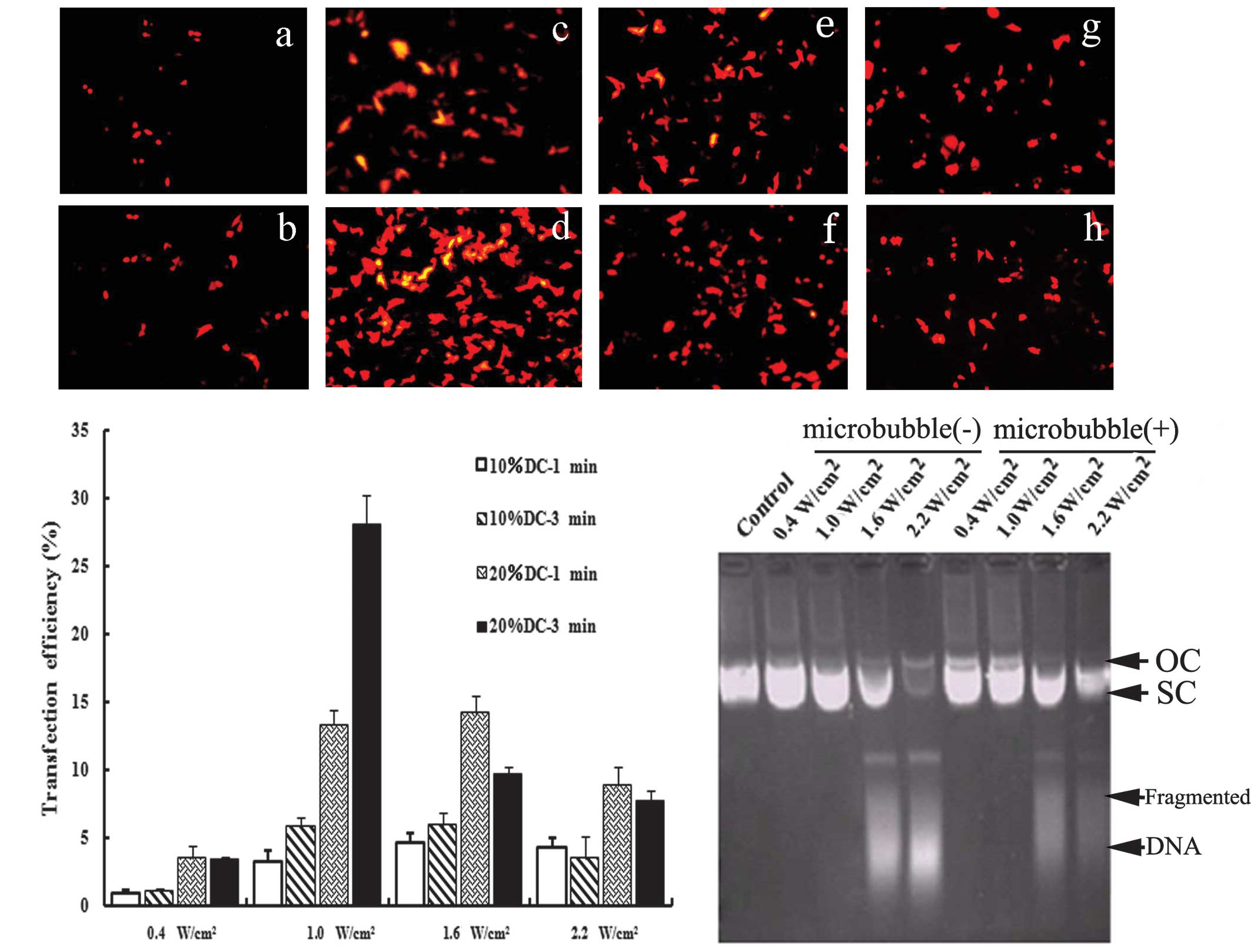 | Figure 1Optimization of the UTMD-mediated
delivery system in vitro. (A) Fluorescence microscopy views
under various ultrasound intensities (0.4, 1.0, 1.6 and 2.2
W/cm2) and exposure times (1 and 3 min), DC=20%. With
the optimal ultrasound parameters (Fig. 1a-d), the transfection
efficiency was increased significantly (28.04±2.27%). (B) Effects
of ultrasound parameters on the transfection efficiency. When cells
were irradiated using 10% DC with 0.4 W/cm2, only a few
expressed RFP. When the ultrasound intensity was increased to 1.0
W/cm2 or when the DC was 20%, the transfection
efficiency increased significantly (P<0.05). However, when the
ultrasound intensity was further increased to 1.6 and 2.2
W/cm2 or the exposure time was extended to 3 min, the
expression of RFP decreased. As compared with 10% DC, irradiating
for 1 and 3 min with 20% DC improved the expression rate
significantly (P<0.05). (C) Effects of ultrasound parameters on
the integrity of plasmid DNA. Ultrasound parameters, 1.0
W/cm2; DC=20%; exposure time, 3 min. The structures and
migration rates of vectors did not markedly change following
irradiation with 0.4 and 1.0 W/cm2. Following treatment
with higher ultrasound intensity, the structures of the vectors
were markedly destroyed and DNA fragments could be observed. Under
the same ultrasound parameters, the addition of SonoVue had no
effect on the integrity of plasmid DNA. UTMD, ultrasound-targeted
microbubble destruction; DC, duty cycle; RFP, red fluorescence
protein; OC, open circular DNA; SC, supercoiled DNA. |
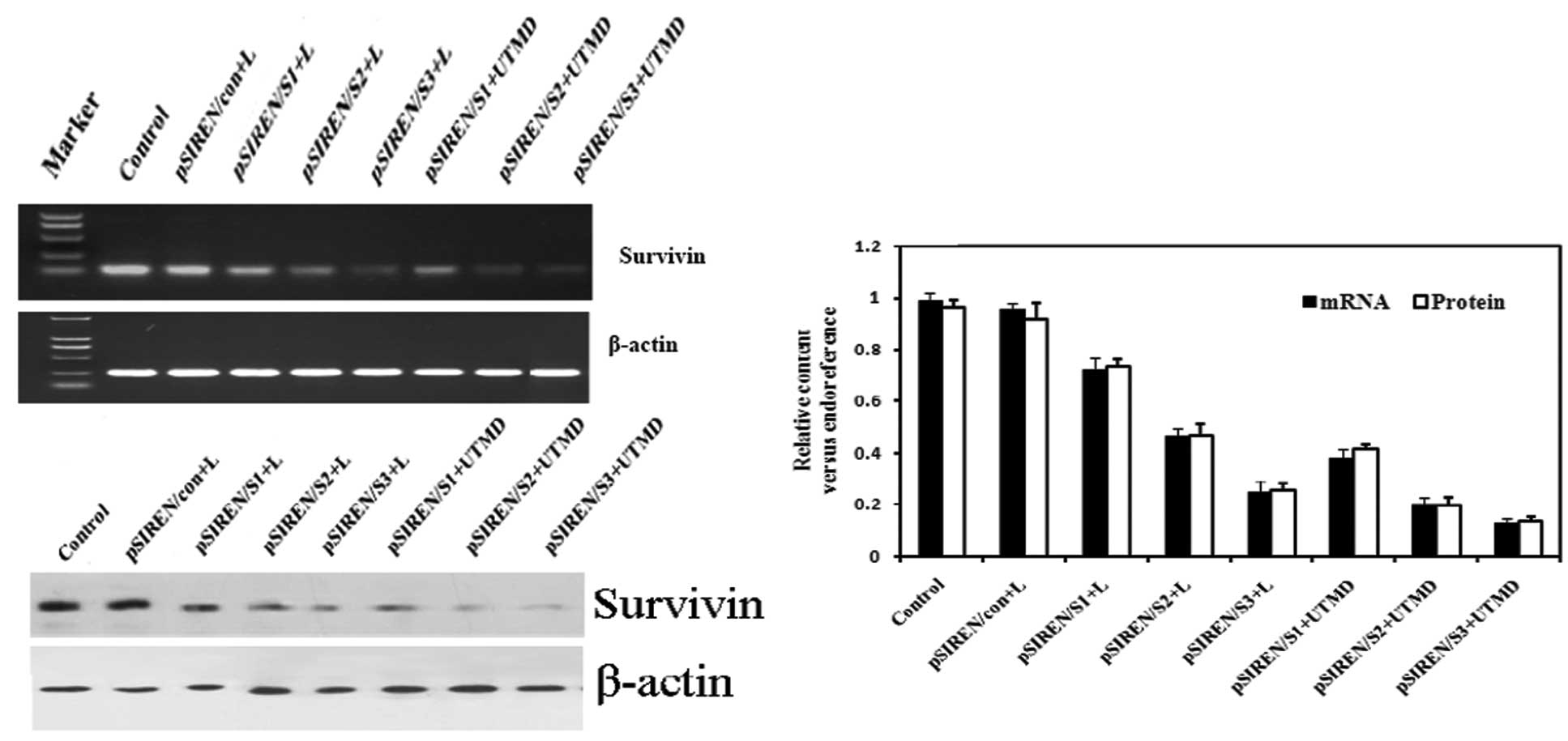
Vector-based shRNAs decreased endogenous
survivin expression in cells
We examined whether delivery of survivin-shRNA by
our UTMD system was capable of efficiently knocking down the
expression of survivin. When shRNA expression vectors were
introduced into HeLa cells, the results of RT-PCR showed that
survivin mRNA expression was markedly knocked down, whereas the
control or pSIREN/con groups did not have this effect (Fig. 2A and C). Furthermore, the expression
levels of survivin mRNA declined markedly in the UTMD-treated
groups as compared with the Lipofectamine- transfected groups
(P<0.05 for all). Western blot analysis also revealed that the
expression of survivin protein was further suppressed in the groups
transfected with pSIREN/S1, pSIREN/S2 and pSIREN/S3 vectors
(Fig. 2B and C). Transfection in
the UTMD group was found to induce the greatest inhibition of
survivin protein expression, as compared with the
Lipofectamine-treated groups (P<0.05 for all).
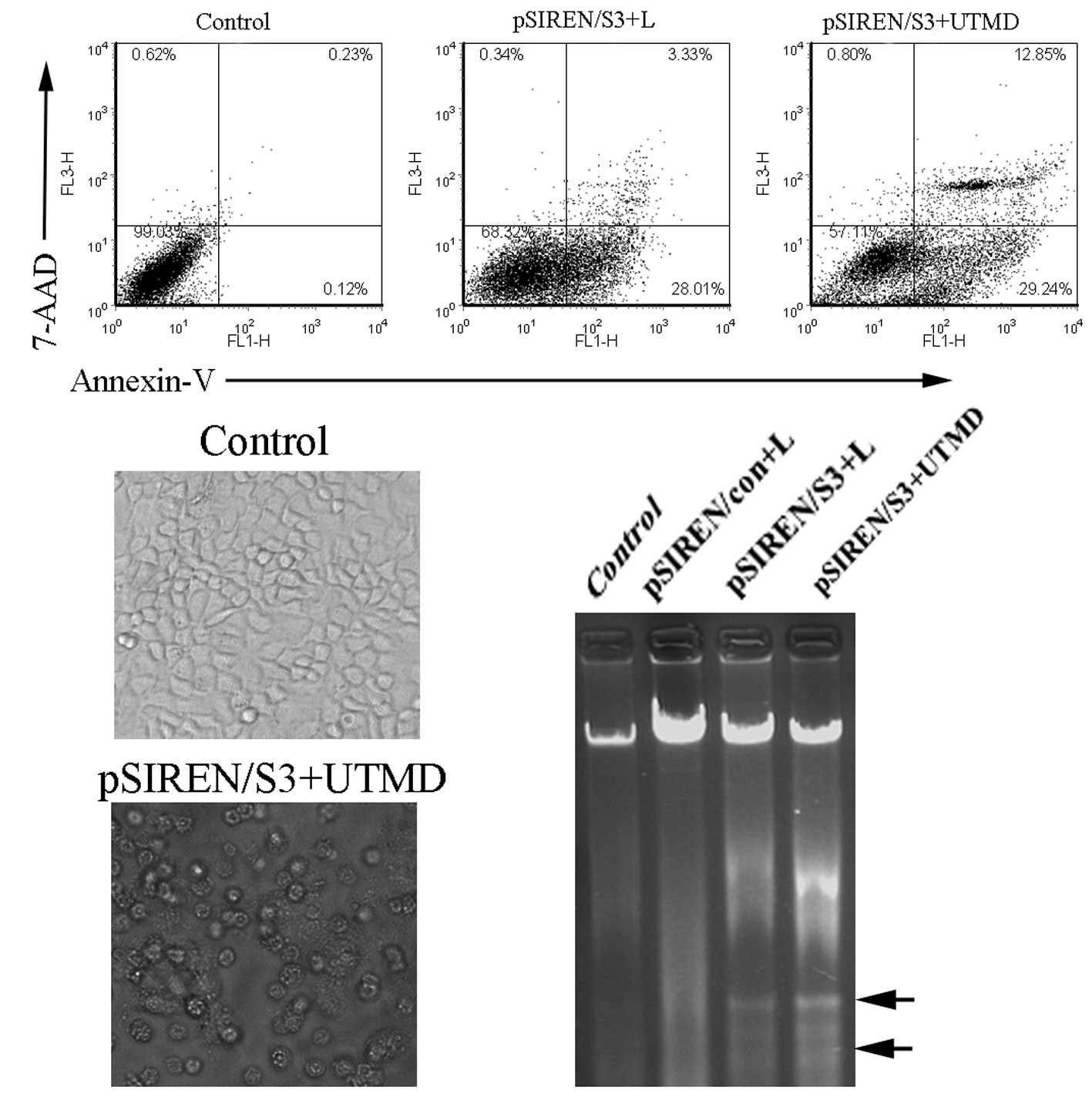
RNAi-targeting survivin inhibited
apoptosis induction
To evaluate the effect of survivin depletion on the
proliferation and apoptosis of HeLa cells in vitro, we used
the flow cytometry assay to detect the effect of loss of survivin
on cell apoptosis. The survivin shRNA construct-transfected HeLa
cells showed a robust induction in cell apoptosis compared to the
control (Fig. 3). Our data
demonstrated that the apoptosis ratio in the pSIREN/S3+UTMD group
(42.03±1.52%) was significantly higher than that of the pSIREN/S3+L
group (31.46±2.78%) (Fig. 3A,
P<0.05). To further determine the impact of survivin shRNA on
cell growth, we detected the morphological changes in HeLa
transfectants and the DNA ladder. The apoptotic cells in the
pSIREN/S3+UTMD group presented typical morphological changes of
cell apoptosis, such as cell shrinkage, round shape and bubbled
cell membranes (Fig. 3B). Moreover,
the apparent DNA ladder was detected in the pSIREN/S3+UTMD or
pSIREN/S3+L groups, whereas there was no marked change in the
control or non-specific vector groups (Fig. 3C).
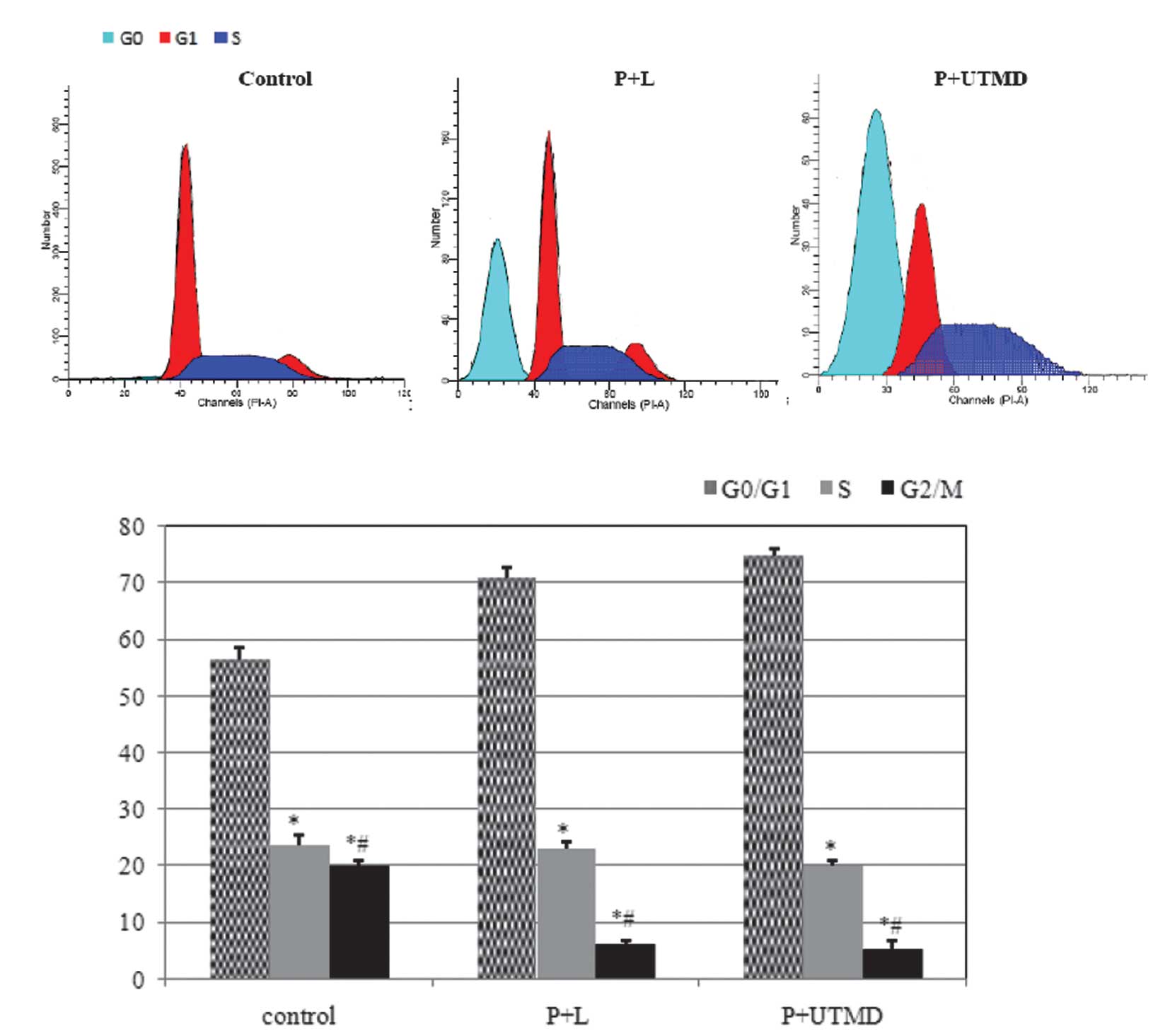
Changes in the cell cycle apoptotic
rate
Cell cycle distribution analysis by flow cytometry
indicated that, as compared with the control group, there were
marked changes in the P+L and P+UTMD groups; a number of cells were
blocked in the G0/G1 phase (70.81±1.78 vs.
74.9±1.21 vs. 56.54±2.01%, P<0.01), reduced sharply in the
G2/M phase (6.11±0.67 vs. 5.20±1.71 vs. 19.89±1.77%,
P<0.01) and S phase (23.08±1.29 vs. 19.89±1.11 vs. 23.56±1.99%,
P<0.01), and an apoptosis peak was observed prior to the
G1 phase (Fig. 4A).
Discussion
Cervical carcinoma is a common high-risk malignancy
in gynecology and its main therapeutic method is radiotherapy.
Apoptosis induction is an effective and promising cancer treatment.
Among the regulators of apoptosis, IAP proteins, such as survivin,
have recently attracted considerable attention for their ability to
suppress an evolutionarily conserved step in apoptosis, potentially
involving direct caspase inhibition. Therefore, survivin has become
a significant target for tumor therapy. In this study, we inhibited
the expression of survivin to promote cell apoptosis and inhibit
cell proliferation.
In a previous study (22), pSIREN/S3 was added to HeLa cells
followed by exposure to ultrasound and SonoVue microbubbles. The
study was designed to evaluate the non-viral gene transfer system
of UTMD compared with other transfection methods. We suggested that
shRNA targeting the survivin gene mediated by the UTMD technique
was capable of effectively inhibiting the expression of target
genes and induce cell apoptosis. In a previous study (23), we hypothesized that three shRNAs
targeting the survivin gene could be transfected by ultrasound with
liposome microbubbles. In this study, we investigated whether the
shRNA expression vectors targeting survivin (pSIREN/S1, pSIREN/S2,
pSIREN/S3 and pSIREN/CON) were capable of being transfected using
the UTMD technique. Following transfection of the vectors into the
human cervical cancer cells, we measured their inhibitory effects
on survivin mRNA and protein expression by semi-quantitative RT-PCR
and western blotting. Transfection efficiency with flow cytometry
assays and the stability of plasmid DNA was detected. The results
indicated that shRNA targeting survivin mediated by the UTMD
technique markedly inhibited the expression of survivin mRNA and
its corresponding protein to a certain degree in vitro.
Moreover, pSIREN/S3 has a higher inhibitory power not only in the
Lipofectamine-transfected group but also in the UTMD-treated group,
and the pSIREN/S3 vector combined with the UTMD method had a better
interfering effect on survivin expression in the HeLa cells, and
was selected for further analysis. The novel data reported in this
study included the combination of ultrasound exposure and SonoVue
microbubbles and optimization of the UTMD technique for the
delivery system of shRNA in vitro, which has not been
performed in previous studies.
The results of our study and other reports (16,30)
have shown that, prior to achieving the optimal parameters, such as
increasing ultrasound intensity, extending irradiation time or DC,
UTMD may improve transfection efficiency. However, when plasmid DNA
was treated with high intensity or low frequency ultrasound, shear
stress mediated by UTMD led to the structural degradation of
plasmid DNA (31). Certain measures
should be taken to protect vectors, to further enhance the
transfection efficiency. Our results reveal that UTMD-mediated
survivin gene mRNA and protein knockdown in HeLa cell lines caused
marked cell apoptosis and that the appearance of DNA fragments was
correlated with the transfection parameters. The optimal
irradiation parameters would achieved higher transfection
efficiency, but exhibited no effect on the integrity of plasmid
DNA.
We hypothesized that the downregulation of survivin
was capable of inducing apoptosis in cancer cells. To explore this
possibility, we measured levels of apoptosis in transfected and
control cells. Experimental data indicated that degradation of
survivin was the main trigger for apoptosis induction. After the
expression of survivin was downregulated, a marked block in the
G0/G1 phase of the cell cycle was observed
and cell apoptosis induction was examined, which was similar to
other reports that did not use this technique (32). These data have laid a foundation for
further investigation into cervical carcinoma therapeutics.
In conclusion, our results showed that survivin
downregulation with shRNA expression vectors mediated by the
optimal UTMD parameters markedly induced cell apoptosis and caused
cell cycle arrest. This study suggests that a combination treatment
may lead to an additive apoptosis-inducing effect on cervical
cancer cells. More studies are required to promote this efficient,
promising novel technique for gene delivery, which is likely to
accelerate the development of therapeutic approaches for human
cervical cancer.
Acknowledgements
This study was supported by the Medical Research
Foundation of Guangdong Province (No. A2010270), and by research
projects of the Guangzhou Education Bureau (No. 10A242), and by
research projects of the Science and Technology Bureau of Guangzhou
Liwan District (20111213067), and by youth research support
projects of the Third Affiliated Hospital of Guangzhou Medical
University (2011Y02).
References
|
1
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lakhani SA, Masud A, Kuida K, Porter GA
Jr, Booth CJ, Mehal WZ, Inayat I and Flavell RA: Caspases 3 and 7:
key mediators of mitochondrial events of apoptosis. Science.
311:847–851. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chipuk JE, Kuwana T, Bouchier-Hayes L,
Droin NM, Newmeyer DD, Schuler M and Green DR: Direct activation of
Bax by p53 mediates mitochondrial membrane permeabilization and
apoptosis. Science. 303:1010–1014. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Knauer SK, Krämer OH, Knösel T, Engels K,
Rödel F, Kovács AF, Dietmaier W, Klein-Hitpass L, Habtemichael N,
Schweitzer A, Brieger J, Rödel C, Mann W, Petersen I, Heinzel T and
Stauber RH: Nuclear export is essential for the tumor-promoting
activity of survivin. FASEB J. 21:207–216. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Carvalho A, Carmena M, Sambade C, Earnshaw
WC and Wheatley SP: Survivin is required for stable checkpoint
activation in taxol-treated HeLa cells. J Cell Sci. 116:2987–2998.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Caldas H, Holloway MP, Hall BM, Qualman SJ
and Altura RA: Survivin-directed RNA interference cocktail is a
potent suppressor of tumour growth in vivo. J Med Genet.
43:119–128. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kappler M, Bache M, Bartel F, Kotzsch M,
Panian M, Würl P, Blümke K, Schmidt H, Meye A and Taubert H:
Knockdown of survivin expression by small interfering RNA reduces
the clonogenic survival of human sarcoma cell lines independently
of p53. Cancer Gene Ther. 11:186–193. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu Q, Fu H, Xing R, Tie Y, Zhu J, Sun Z
and Zheng X: Survivin knockdown combined with apoptin
overexpression inhibits cell growth significantly. Cancer Biol
Ther. 7:1053–1060. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vlassov AV, Korba B, Farrar K, Mukerjee S,
Seyhan AA, Ilves H, Kaspar RL, Leake D, Kazakov SA and Johnston BH:
shRNAs targeting hepatitis C: effects of sequence and structural
features, and comparision with siRNA. Oligonucleotides. 17:223–236.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sui G, Soohoo C, Affar el B, Gay F and Shi
Y, Forrester WC and Shi Y: A DNA vector-based RNAi technology to
suppress gene expression in mammalian cells. Proc Natl Acad Sci
USA. 99:5515–5520. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
McAnuff MA, Rettig GR and Rice KG: Potency
of siRNA versus shRNA mediated knockdown in vivo. J Pharm Sci.
96:2922–2930. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Boudreau RL, Monteys AM and Davidson BL:
Minimizing variables among hairpin-based RNAi vectors reveals the
potency of shRNAs. RNA. 14:1834–1844. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pardridge WM: shRNA and siRNA delivery to
the brain. Adv Drug Deliv Rev. 59:141–152. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo DP, Li XY, Sun P, Tang YB, Chen XY,
Chen Q, Fan LM, Zang B, Shao LZ and Li XR: Utrasound-targeted
microbubble destruction improves the low density lipoprotein
receptor gene expression in HepG2 cells. Biochem Biophys Res
Commun. 343:470–474. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen ZY, Xie MX, Wang XF, Lü Q and Ding
SW: Efficient gene delivery to myocardium with ultrasound targeted
microbubble destruction and polyethylenimine. J Huazhong Univ Sci
Technolog Med Sci. 28:613–617. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen ZY, Xie MX, Wang XF and Lü Q: Effects
of lipid shell microbubble on ultrasound mediated EGFP gene
delivery to transplanted tumors: initial experience. Chin-Ger J
Clin Oncol. 7:424–428. 2008. View Article : Google Scholar
|
|
17
|
Kinoshita M and Hynynen K: A novel method
for the intracellular delivery of siRNA using microbubble-enhanced
focused ultrasound. Biochem Biophys Res Commun. 335:393–399. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saito M, Mazda O, Takahashi KA, Arai Y,
Kishida T, Shin-Ya M, Inoue A, Tonomura H, Sakao K, Morihara T,
Imanishi J, Kawata M and Kubo T: Sonoporation mediated transduction
of pDNA/siRNA into joint synovium in vivo. J Orthop Res.
25:1308–1316. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tran MA, Gowda R, Sharma A, Park EJ, Adair
J, Kester M, Smith NB and Robertson GP: Targeting V600EB-Raf and
Akt3 using nanoliposomal-small interfering RNA inhibits cutaneous
melanocytic lesion development. Cancer Res. 68:7638–7649. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vandenbroucke RE, Lentacker I, Demeester
J, De Smedt SC and Sanders NN: Ultrasound assisted siRNA delivery
using PEG-siPlex loaded microbubbles. J Control Release.
126:265–273. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Zheng Y, Yang F, Zhao P and Li H:
Survivin small interfering RNA transfected with a microbubble and
ultrasound exposure inducing apoptosis in ovarian carcinoma cells.
Int J Gynecol Cancer. 20:500–506. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen ZY, Liang K, Xie MX, Wang XF, Lü Q
and Zhang J: Novel ultrasound-targeted microbubble destruction
mediated short hairpin RNA plasmid transfection targeting survivin
inhibits gene expression and induces apoptosis of HeLa cells. Mol
Biol Rep. 36:2059–2067. 2009. View Article : Google Scholar
|
|
23
|
Chen ZY, Liang K, Liu JH, Xie MX, Wang XF,
Lü Q, Zhang J and Fang LY: Enhancement of survivin gene
downregulation and cell apoptosis by a novel combination: liposome
microbubbles and ultrasound exposure. Med Oncol. 26:491–500. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ling X and Li F: Silencing of
antiapoptotic survivin gene by multiple approaches of RNA
interference technology. Biotechniques. 36:450–454. 2004.PubMed/NCBI
|
|
25
|
Mehier-Humbert S, Bettinger T, Yan F and
Guy RH: Ultrasound- mediated gene delivery: kinetics of plasmid
internalization and gene expression. J Control Release.
104:203–211. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zarnitsyn VG and Prausnitz MR: Physical
parameters influencing optimization of ultrasound-mediated DNA
transfection. Ultrasound Med Biol. 30:527–538. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kodama T, Tomita Y, Koshiyama K and
Blomley MJ: Transfection effect of microbubbles on cells in
superposed ultrasound waves and behavior of cavitation bubble.
Ultrasound Med Biol. 32:905–914. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li H, Wang S, Zhu T, Zhou J, Xu Q, Lu Y
and Ma D: Pin1 contributes to cervical tumorigenesis by regulating
cyclin D1 expression. Oncol Rep. 16:491–496. 2006.PubMed/NCBI
|
|
29
|
Ochiai N, Uchida R, Fuchida S, Okano A,
Okamoto M, Ashihara E, Inaba T, Fujita N, Matsubara H and Shimazaki
C: Effect of farnesyl transferase inhibitor R115777 on the growth
of fresh and cloned myeloma cells in vitro. Blood. 102:3349–3353.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liang HD, Lu QL, Xue SA, Halliwell M,
Kodama T, Cosgrove DO, Stauss HJ, Partridge TA and Blomley MJ:
Optimisation of ultrasound-mediated gene transfer (sonoporation) in
skeletal muscle cells. Ultrasound Med Biol. 30:1523–1529. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kuo JH, Jan MS and Sung KC: Evaluation of
the stability of polymer-based plasmid DNA delivery systems after
ultrasound exposure. Int J Pharm. 257:75–84. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang R, Ma L, Zheng M, Ren J, Wang T,
Meng Y, Zhao J, Jia L, Yao L, Han H, Li K and Yang A: Survivin
knockdown by short hairpin RNA abrogates the growth of human
hepatocellular carcinoma xenografts in nude mice. Cancer Gene Ther.
17:275–288. 2010. View Article : Google Scholar : PubMed/NCBI
|