Introduction
Myelodysplastic syndrome (MDS) is thought to arise
due to multiple alterations in a hematopoietic stem cell (HSC)
(1,2). The status of self-renewal,
proliferation and differentiation of HSCs depends on the
microenvironment in which the HSCs are located. Although little is
known with regard to the microenvironmental cues that govern HSC
self-renewal, mounting evidence has shown that stem cell
development requires a niche, which is defined as a subset of
tissue cells and extracellular substrates that can harbor HSCs and
control their proliferation, differentiation and function in
vivo (3–7). Recent advances in cancer biology
indicate the importance of the tumor environment during the
initiation and development of the cancer cell clone. The niche in
the bone marrow (BM) where the HSCs are located may play a pivotal
role in the initiation and progression of MDS. Thus far, two main
niche models have been identified: osteoblastic (8,9) and
perivascular cell niche (10–12).
Adhesion molecules located on the membrane within the niche execute
pivotal roles in hematopoiesis. Cadherins, a group of
Ca2+-dependent cell adhesion molecules, including
N-cadherin and E-cadherin, are typical representatives of these
adhesion molecules. N-cadherin is expressed in long-term (LT)-HSCs
and in a subpopulation of osteoblasts in BM (9,13,14).
N-cadherin also retains HSCs in the niche via homophilic adhesion,
which regulates quiescence and is required for HSCs to maintain in
an undifferentiated dormant status (9,15,16).
E-cadherin is expressed within human BM stromal cells,
CD34+ stem cells and perivascular niche cells (17,18).
E-cadherin uniquely regulates the self-renewal of human embryonic
stem cells (hESCs) via functional interactions between E-cadherin
and a small Ras family G protein, Ras-proximate-1 (Rap1) (19).
Adhesion molecules located on the cell membrane need
a downstream effector to transmit signals. Kirstetter et al
showed that β-catenin interacted directly with cadherins, playing a
key role in determining blood cell formation and determining their
fate prior to leaving the stem cell compartment in the BM (20). Recent evidence suggests that there
are two pools of β-catenin (21);
one is linked to the cadherin complex at cell-to-cell adherent
junctions (AJs) by a β-catenin binding domain in the cytoplasm
(22) and stabilizes the
interaction with the cytoskeleton (23,24),
which prevents β-catenin degradation. The other β-catenin pool is
regulated by the well-understood canonical Wnt signaling pathway
(25,26) (Fig.
1). In the absence of Wnt, cytoplasmic β-catenin forms a
complex with the scaffolding protein Axin, the tumor suppressor
adenomatous polyposis coli gene product (APC), casein kinase 1
(CK1) and glycogen synthase kinase 3 (GSK3). The complex is
phosphorylated by CK1 and subsequently by GSK3. Phosphorylated
β-catenin is recognized by an E3 ubiquitin ligase, resulting in its
ubiquitination and proteasomal degradation. In the presence of a
Wnt ligand, the Wnt ligand binds to the Frizzled (Fz or Fzd)
receptor and its coreceptor, low-density lipoprotein
receptor-related protein 5 or 6 (LRP5/6). The formation of a
Wnt-Fz-LRP complex, together with the recruitment of the
scaffolding protein Dishevelled (Dvl), results in LRP
phosphorylation and activation and the recruitment of the Axin
complex to the receptors. These events lead to the inhibition of
Axin-mediated β-catenin phosphorylation and thus to the
stabilization of β-catenin. The two pools of β-catenin maintain a
dynamic balance in normal cells and β-catenin may be transported to
the nucleus to play a pivotal role in regulating the key
developmental gene expression programs, including activating the
transcription of c-myc, a candidate of hematopoiesis (27).
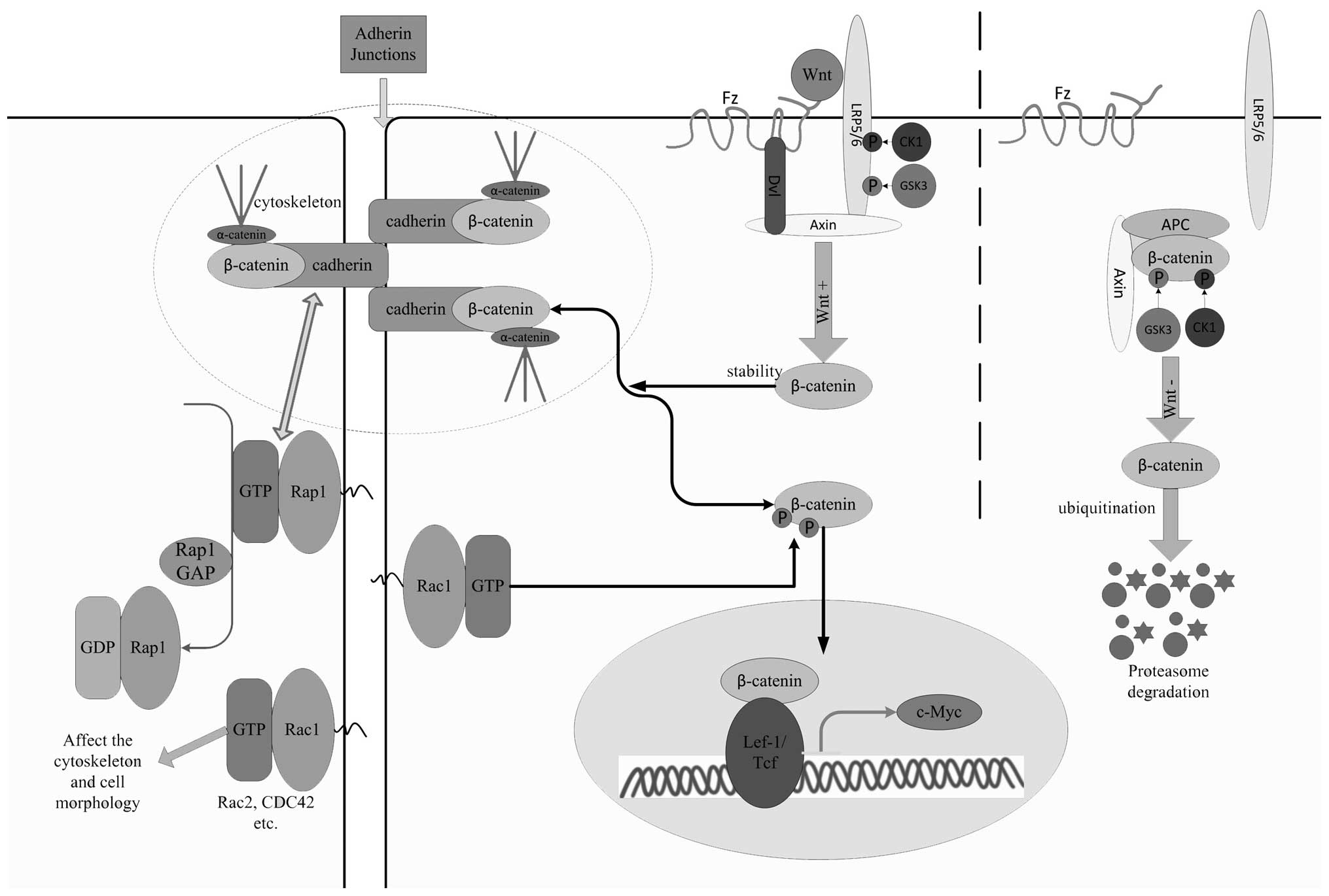 | Figure 1A possible mechanism for regulating
the hematopoiesis of HSCs in MDS is shown. Cadherin and Rap1 have a
positive feedback-like interplay with each other. Both are
downregulated in MDS. In the absence of Wnt, cytoplasmic β-catenin
forms a complex with Axin, APC, GSK3 and CK1, which is
phosphorylated by CK1 and GSK3. Phosphorylated β-catenin is
recognized by an E3 ubiquitin ligase, resulting in its
ubiquitination and proteasomal degradation. In the presence of a
Wnt ligand, a receptor complex forms between Fz and LRP5/6. Dvl
recruitment by Fz leads to LRP5/6 phosphorylation and Axin
recruitment. These events lead to the inhibition of Axin-mediated
β-catenin phosphorylation and therefore the stabilization of
β-catenin. β-catenin, which keeps a dynamic balance between the
cadherin-β-catenin complex and the Wnt signal-regulated
conformation, accumulates in the cytoplasm as the downregulation of
cadherins occurs and is transported to the nucleus. Continuous
activation of β-catenin through the interaction with T-cell
factor/Lef transcription factors results in the aberrant
transcription of c-myc, which promotes the differentiation of HSCs
at the expense of self-renewal and blocks before the terminal
maturation stage of blood cells, leading to the exhaustion of the
HSC pool and peripheral cytopenia. Rac1, a Rac subfamily member of
the Rho GTPase family, is indispensable for the nuclear
translocation of β-catenin via phosphorylation, and Rac1, Rac2 and
CDC42 affect cell morphology. Rap1, Ras-proximate-1; CK1, casein
kinase 1; GSK3, glycogen synthase 3; Dvl, dishevelled; LRP5/6,
low-density lipoprotein receptor-related protein 5 or 6; Fz,
frizzled; MDS, myelodysplastic syndrome; HSC, hematopoietic stem
cell; APC, adenomatous polyposis coli gene product. |
Dysplasia is another typical characteristic of MDS.
Rho GTPases, belonging to a family of small G proteins and cycling
between an inactive GDP-bound and an active GTP-bound status, are
closely correlated with the cytoskeleton and the cell shape
(28) and are involved in
hematopoiesis and hemopathies (29). In the active conformation, G
proteins interact with effector proteins, which induce downstream
signaling events. The GDP-GTP cycle is regulated by guanine
nucleotide exchange factors (GEFs) and GTPase-activating proteins
(GAPs). GEFs induce the release of bound GDP which is subsequently
replaced by the more abundant GTP, whereas GAPs hydrolyze GTP to
switch back to the GDP-binding form. Only a small fraction of Rho
GTPases within the cell are in the active state and associated with
membranes at any given time. Rho-specific guanine nucleotide
dissociation inhibitors (RhoGDIs) associate with and maintain the
inactive pool in the cytosol.
Since the cellular component and the phenotype of
cell clones in the BM of MDS patients is extremely diverse and no
cell line or other model of MDS has been established thus far, it
is difficult to study the pathophysiology of MDS. Comprehensively
screening differentially expressed genes correlated with
hematopoiesis and morphological dysplasia in MDS patients using
whole genomic array and analyzing the internal correlation among
screened genes may reveal important clues concerning the dysplastic
hematopoiesis of MDS. As MDS is a multistep procedure, low-risk
MDS, including MDS-refractory anemia (RA), may be the initial step
of MDS. Analyzing certain differentially expressed genes in these
groups of patients may provide significant clues to understanding
the initial molecular mechanism of MDS. Therefore, we firstly
examined gene expression profiles from CD34+ cells with
MDS using oligonucleotide microarray. The adhesion molecules, Wnt
signaling pathway, Rho GTPases and the associated molecules were
analyzed. The selected differentially expressed genes were
validated using CD34+ cells or mononuclear cells (MNCs)
from patients with MDS-RA (resembling WHO 2008 classification as
indicated in Table I). The gene
expression data suggest that the cadherins-β-catenin-c-myc
signaling axis is closely correlated with the incidence of MDS and
that two small G proteins, Ras-proximate-1 GTPase-activating
protein (Rap1GAP) and Rac2, are crucial in the initial development
of MDS and act as new molecular markers for the diagnosis of
MDS.
 | Table IPatient clinical and laboratory
characteristics. |
Table I
Patient clinical and laboratory
characteristics.
| Samples | Age (years) | Gender | FAB | WHO (2008) | Karyotype | IPSS |
|---|
| Array group |
| 1 | 34 | M | RAEBt | AML (MDS) | 46, XY, tandem
duplication(1)(q12q24) | High |
| 2 | 70 | F | RAEBt | AML (MDS) | 47, XX, +8 | High |
| 3 | 43 | M | RAEB | RAEB-1 | 45, XY,
5q-,6p+,−7 | INT-2 |
| 4 | 40 | F | RAEB | RAEB-1 | 46, XX,
der(6)/47,idem,+8 | INT-2 |
| 5 | 53 | F | RA | RCMD | 47, XX,
+8,9q-[4]/48.idem,+der(1)[7] | INT-2 |
| 6 | 40 | M | RA | RCMD | 44, XY,
del(5)(q12q31),-7,-18 | INT-2 |
| 7 | 51 | M | RAS | RAS | 45, XY, -5,-6 | INT-1 |
| RQ-PCR group using
CD34+ cells |
| 1 | 35 | F | RA | RCMD | 46, XX | INT-1 |
| 2 | 39 | M | RA | RCUD | 46, XY | Low |
| 3 | 57 | M | RA | RCMD | 46, XY | INT-1 |
| 4 | 58 | M | RA | RCMD | 46, XY | INT-1 |
| 5 | 27 | F | RA | RCMD | 46, XX | INT-1 |
| 6 | 72 | M | RA | RCMD | 46, XY | Low |
| 7 | 56 | F | RA | RCUD | 46, XX | Low |
| 8 | 78 | M | RA | MDS-U | 46, XY | INT-1 |
| 9 | 57 | F | RA | RCUD | 46, XX | Low |
| 10 | 57 | M | RA | RCMD | 46, XY | INT-1 |
| 11 | 27 | M | RA | MDS-U | 46, XY | INT-1 |
| 12 | 46 | M | RA | RCMD | 46, XY | INT-1 |
Materials and methods
Patients
CD34+ cells isolated from eight samples
were analyzed by microarray hybridization for gene expression
profiling. These samples consisted of two RA; one RA with ringed
sideroblast (RAS); two RA with an excess of blasts (RAEB); two RA
with excess of blasts in transformation (RAEBt) in accordance with
FAB classification (Table I) and a
control which was mixed from several fracture patients with no
other diseases and a normal peripheral cell count. CD34+
cells or MNCs isolated from two paired groups were subjected to
real-time quantitative polymerase chain reaction (RQ-PCR). The
CD34+ cell group included 12 RA patients (Table I) and 12 bone fracture patients (5
male, 7 female, aged from 35 to 69 years old), and the MNC group
included 42 patients with RA (27 male, 15 female, aged from 27 to
85 years old) and 32 patients with bone fractures (17 male, 15
female, aged from 34 to 75 years old). The RA patients [WHO (2008)
RCUD, RCMD and MDS-U] had a normal karyotype and all the bone
fracture patients had normal peripheral cell counts and presented
no other diseases. The collection of samples was approved by the
Ethics Committee at The First Affiliated Hospital, Soochow
University, and informed consent was obtained from patients.
Sample preparation
Heparinized BM samples were obtained by aspiration
from the posterior iliac crest of the MDS patients and the normal
controls were obtained during surgery. To prevent the activation of
the cells by technical manipulation, fresh BM was processed
immediately following aspiration to select MNCs using lymphocyte
separation medium within the subsequent 4 h. The CD34+
cells analyzed by microarray were purified according to the
manufacturer's instructions (30)
and CD34+ cells subjected to RQ-PCR were sorted using
flow cytometry (BD, FACS). Total RNA was extracted using TRIzol
(Invitrogen, Carlsbad, CA, USA) according to the manufacturer's
instructions with minor modifications.
Oligonucleotide microarray and data
analysis
A total of eight pieces of Human Genome U133 Plus
2.0 Array were used, which were hybridized with the amplified
products from seven MDS patients and one normal control. Sample
preparation and microarray processing were conducted according to
the manufacturer's instructions (Affymetrix, Santa Clara, CA, USA).
Since there were few cells from which the RNAs could be extracted,
two rounds of in vitro transcript amplification were
necessary prior to hybridization on the oligonucleotide
microarray.
Data analysis
GeneChip image analysis was conducted using the
Microarray Analysis Suites version 5.0 (Affymetrix). Data analysis
was performed with the GeneSpring software version 6.0 (Silicon
Genetics, San Carlos, CA, USA). The samples, obtained from MDS
patients, were analyzed independently and compared to a normal
control. The samples were normalized for expression levels in each
chip to reference values. Statistical analyses of the average
expression level (analysis of variance, ANOVA) were performed for
each individual gene in the test samples from the MDS patients,
based on a comparison with the normal control. Fold changes for the
log ratios are shown (Fig. 2). The
signal Log2 ratio of ≥1 was considered as ‘up’, whereas the signal
Log2 ratio of ≤-1 as ‘down’. The adhesion molecules, Wnt signaling
pathway, Rho GTPases and the associated molecules were analyzed.
Only the genes were found to have a similar tendency to change when
no less than three samples from all the chips were analyzed.
RQ-PCR
RQ-PCR was used to validate the expression data for
the selected genes. The primers and TaqMan probes were designed
with Primer Express 3.0 and Beacon Designer 7.0. The expression
level of ABL (tyrosine-protein kinase ABL1 isoform a) was used to
normalize differences in input cDNA. Each sample reaction was
performed in triplicate and a reverse-transcriptase negative
control was also tested to exclude any contaminating DNA
amplification. The expression ratio was calculated as
2−n, where n is the CT value difference for each sample
(selected gene minus ABL). The difference between patients and
healthy control subjects was assessed using the Wilcoxon
non-parametric test.
Results
Gene expression profile analysis in
patients with MDS using microarray
Following normalization, the genes most involved
with hematopoiesis on the Affymetrix chips were selected for final
analysis, including adhesion molecules (cadherin and integrin),
molecules involved in the Wnt signaling pathway (APC, Axin, CK1,
Dvl, GSK3, LRP5 and LRP6 among others), Rho GTPase family members,
GAPs, GEFs, GDIs and c-myc-related molecules. Among these
molecules, the gene expression for N-cadherin, E-cadherin and c-myc
binding protein tended to be downregulated and the gene expression
for β-catenin, c-myc promoter binding protein, Rap1GAP, Rac1, Rac2
and CDC42 tended to be upregulated (Fig. 2). These nine genes were selected as
targets for further analysis.
Detection of the selected gene expression
level using RQ-PCR
To explore the expression profiles of the selected
genes in patients with an early stage of MDS, we prepared cDNA
using CD34+ cells from 12 MDS-RA patients with a normal
karyotype [WHO (2008) RCUD, RCMD and MDS-U]and 12 normal control
subjects. These cDNAs were applied to RQ-PCR analysis with
predeveloped and confirmed specific primers (Table II). A number of critical molecules
involved in the Wnt signaling pathway were also included in the
panel of RQ-PCR detection to exclude the impact of their effects,
including APC, Axin, CK1, Dvl, GSK3, LRP5 and LRP6, although no
change in expression was shown on the chips. The results showed
that β-catenin, c-myc promoter binding protein, Rap1GAP, Rac1, Rac2
and CDC42 were expressed at higher levels when compared with normal
controls (P<0.0001, P<0.0001, P<0.0001, P=0.0007, P=0.0001
and P=0.0086, respectively), while N-cadherin, E-cadherin and c-myc
binding protein were expressed at lower levels (P=0.0011, P=0.0001
and P<0.0001, respectively). As c-myc is a significant molecule
in the regulation of cell functions and was not present on the
chips, the c-myc binding protein and c-myc promoter binding protein
are indirect molecular substitutes that confirm the expression
changes of c-myc. We also detected the transcripts of c-myc with
RQ-PCR. The results did not show a significant difference between
the groups (P=0.6859), although the MDS-RA group had a relatively
higher median. Rap1GAP is a negative regulator of Rap1 and it was
observed that Rap1 had random expression changes on chips; thus, we
also detected the transcripts of Rap1 using RQ-PCR, but no
difference was observed (P=0.2598). Compensation may occur from
signal-induced proliferation-associated gene 1 (SPA-1), another
negative regulator of Rap1, which is secreted by only hematopoietic
progenitor cells and is gradually replaced by Rap1GAP as the cells
differentiated to mature cells (27). To exclude this compensation, RQ-PCRs
were also performed to detect the gene expression of SPA-1. The
results showed a significantly higher expression in the MDS-RA
group (P=0.0024; Fig. 3). As
CD34+ cells are extremely difficult to obtain from
patients diagnosed with the early stage of MDS, MNCs originating
from the MDS stem cell may also provide several critical clues to
understanding the pathophysiology of MDS. Although the MNCs in BM
are markedly heterogeneous, we also detected the same target gene
expression profiles as observed in the CD34+ cells using
RQ-PCR. A total of 42 patients with MDS-RA with a normal karyotype
[WHO (2008) RCUD, RCMD and MDS-U] and 32 normal control subjects
were used. The results show that Rap1GAP and Rac2 were expressed at
a higher level in the MDS-RA group (P<0.0001; Fig. 3).
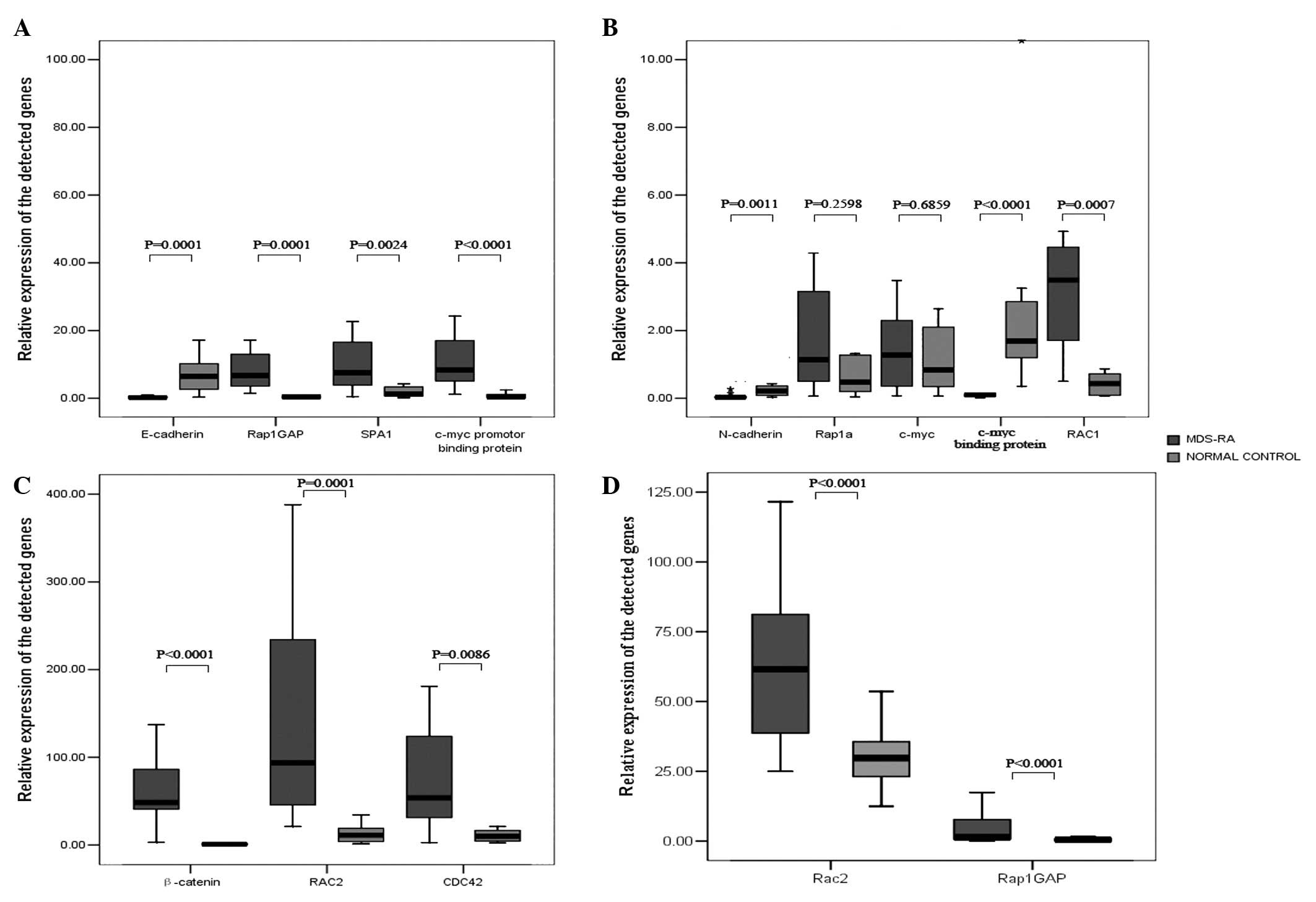 | Figure 3Box graphs showing the expression
levels of β-catenin, E-cadherin, N-cadherin, c-myc, c-myc binding
protein, c-myc promoter binding protein, Rap1GAP, SPA-1, Rap1,
Rac2, Rac1 and CDC42 in patients with MDS-RA. (A-C) When
CD34+ cells were used from 12 MDS-RA patients, gene
expression levels of β-catenin, c-myc promoter binding protein,
Rap1GAP, SPA-1, Rac1, Rac2 and CDC42 were higher than those of the
normal control subjects (P<0.0001, P<0.0001, P<0.0001,
P=0.0024, P=0.0007, P=0.0001 and P=0.0086, respectively), while the
gene expression levels of N-cadherin, E-cadherin and c-myc binding
protein were lower than those of the normal control subjects
(P=0.0011, P=0.0001 and P<0.0001, respectively), and no
significant difference was found with Rap1 and c-myc (P=0.2598,
P=0.6859). (D) When MNCs were used from 42 MDS-RA patients, only
Rap1GAP and Rac2 showed higher expression levels than the normal
controls (P<0.0001 and P<0.0001, respectively). Rap1GAP,
Ras-proximate-1 GTPase-activating protein; SPA-1, signal-induced
proliferation-associated gene 1; MDS, myelodysplastic syndrome; RA,
refractory anemia. |
| Table IIPrimer sequences and probes for
RQ-PCR. |
Table II
Primer sequences and probes for
RQ-PCR.
| Gene | Forward (5′ to
3′) | Reverse (5′ to
3′) | Probe (5′ FAM to 3′
TAMRA) |
|---|
| β-catenin |
ACCTATACTTACGAAAAACTAC |
CCACCAGCTTCTACAATA |
CTGAAGGTGCTATCTGTCTGCTC |
| E-cadherin |
CAAGTGACCACCTTAGAG |
GAATTTGCAATCCTGCTT |
CCTTCCTACAGACGCCAGC |
| N-cadherin |
CAGTGTACAGAATCAGTG |
CAACAGTAAGGACAAACA |
CCTACTGGACGGTTCGCC |
| Rap1GAP |
CAAGGTGGGAGAGAGAGTTTGAG |
CCCCACATATCACATCCTCCTTG |
CCAGGGGCTAAGCTCTCCACACGG |
| MYCBP |
CCATCTAACTCATTACCTTAA |
CTAAACTGTCTTGGCTAAA |
ACCAGTGCCATCATTCCTAATCAG |
| MYCPBP |
AGAATTGTACTTGAGTCATT |
TTTAGTCGTTACCTTTAAGT |
CTTGTCACCTCTTGTCACCTGG |
| RAC2 |
GAAGCATCTACCCGTTCACTCC |
CAAGTTGTGGCAGCAACCATC |
CCACCCCACGCCTGACTCCCCTC |
| RAC1 |
CAGATTACCGACACTGTCACTTG |
CAGACCCAAAGGAACATCAATAGG |
TGACCCTCTTTACCTCGCCCACGC |
| CDC42 |
GCCTTAATTATACATTTGAACTT |
TGAATTATCCTAATCATATGCTTA |
ACCATTCCTCGTCACCACAAT |
| c-myc |
GTATTTCTACTGCGACGA |
CAGCTCGAATTTCTTCCA |
CTACCAGCAGCAGCAGCA |
| Rap1a |
AGAATTTAGATCTTATATTGGTTTG |
AAAGGAGAAATACTATTGTCTTAT |
TTCCAAGAGATATACACAGAGCAAT |
| SPA1 |
GCAGCTCTCTGTCGGATGAG |
TGTCTCACTGTCAGCACTGG |
CCCAGTCCTGCCCAACACCACCC |
| APC |
AATCAGAGTTGCGATGGAAGAAC |
GAAGTATGTCCTTTTCGATTTGCTG |
TCTGGCTATTCTTCGCTGTGCTCGT |
| Axin |
TATTATGTCAATGCCGGCTATGC |
CCATCCACGCTGCTGTCC |
CCAACGACAGCGAGCAGCAGAGCC |
| CNSK1D |
GACGTCACAGCGCGATGG |
TCAATACGGGGCGGATGGG |
CGCCGCCGCCGCTGCTCC |
| Dvl |
AGGGTGCTCACTCGGATGC |
GCCACATTTGGGTGGAAGGAG |
CCGATGCCGCCTGTCCGCTCAAG |
| GSK3B |
CCTGGCGCAATGAGGAGAG |
GGAGATGCGACGGGAAACG |
CCGCCGCCACCGCCACCG |
| LRP5 |
GACCTGCATCGTGCCTGAG |
TCTGTCCAGTAGATGTGGTTGTTG |
CTTCACCAGCAGAGCCGCCATCCAC |
| LRP6 |
GGTTATGAATACTGATGGCACTGG |
ATCACTTCCCTCTCTGCACTTC |
TTCAATGCTACGCCTCTGCCAGTCA |
| ABL |
GATACGAAGGGAGGGTGTACCA |
CTCGGCCAGGGTGTTGAA |
TGCTTCTGATGGCAAGCTCTACGTCTCCT |
Discussion
Features of MDS include refractory cytopenia,
dysplastic cell morphology and a propensity towards malignant
transformation. Although the underlying causes of primary MDS have
yet to be defined, the dysplastic hematopoiesis triggered by the
defection of HSCs themselves and the defection of a reciprocal
interaction between HSCs and their specific microenvironment is
widely accepted (9). The malignant
transformation in MDS has been suggested to be a multistep
procedure; the general evolution model progresses from MDS-RA to
RAEB, RAEBt and acute leukemia, when using FAB classification.
According to the ‘two hit’ theory, a second molecular event
generates a distinct HSC clone with increased proliferation, which
may gradually become a malignant clone and eventually transform
into acute myeloid leukemia. Low-risk MDS, such as MDS-RA,
especially with normal karyotype, may indicate the early stages of
MDS. Therefore, further study focusing on this group of patients
with the disease to explore several gene expression profiles
correlated with hematopoiesis should provide clues and insight into
the potential pathophysiology of MDS and the initial molecular
event.
While investigating an interaction between HSCs and
the microenvironment, we initially found that N-cadherin and
E-cadherin were expressed at lower levels in the MDS-RA group [WHO
(2008) RCUD, RCMD and MDS-U] using CD34+ cells.
Cadherins are cell adhesion molecules responsible for
Ca2+-dependent cell-to-cell interaction, which are
involved in the formation of AJs and are involved in the
maintenance of the continuous hematopoiesis of HSCs via the
regulation of the interaction between HSCs and the microenvironment
(9). N-cadherin acts as a signal
that triggers a subpopulation of HSCs to transit between ‘reserved’
(dormant) and ‘primed’ (active) states by the regulation of its
expression (16). The enhanced cell
adhesion induced by N-cadherin overexpression results in slower HSC
division (14), while the
downregulation of N-cadherin was observed in HSCs prior to
detachment from the niche and reentry into the cell cycle (31). The impairment of cadherin-mediated
anchorage, characteristic of HSCs, via AJs results in HSCs that are
able to undergo apoptosis (32,33).
Therefore, the downregulation of N-cadherin may be in concordance
with an elevated ratio of apoptosis to proliferation (34) and a faster senescence rate (35) in early MDS.
A model has not yet been suggested to study the
contribution of E-cadherin to the maintenance of HSCs, with the
exception of the recently reported functional interactions between
E-cadherin and Rap1 that uniquely regulate the self-renewal of
hESCs (19). In contrast to
epithelial cells (36), hESCs lack
a negative feedback mechanism between Rap1 and E-cadherin, but
exhibit a positive feedback, i.e., the downregulation of E-cadherin
is accompanied by the same expression change of Rap1 and vice
versa. Rap1 is essential during early embryo development (37–39)
and it is crucial in the formation and maintenance of
cadherin-mediated cell-to-cell junctions (40–44).
Therefore, Rap1 is required for the maintenance of hESCs in an
undifferentiated state via Rap1-mediated interaction with the
microenvironment. The cells exhibited a loss of undifferentiated
characteristics with reduced levels of Rap1, but the inhibition of
Rap1 suppressed colony formation, self-renewal of hESCs and
resulted in an increase in cell death due to the defect of
interaction with the microenvironment (19). The present study showed that
E-cadherin was significantly downregulated in MDS-RA, however, the
status of Rap1 expression may be critical. Using the chips and
RQ-PCR, direct evidence for the downregulation of Rap1 at the
transcriptional level was not found. However, Rap1GAP and SPA-1,
two negative regulators of Rap1, which may convert Rap1 back to its
inactive GDP-bound state (45–47),
were upregulated in MDS-RA samples and we previously reported that
MDS samples expressed more Rap1GAP at the mRNA and protein levels
(48,49), thus the activity of Rap1 should be
downregulated. Therefore a positive feedback mechanism between
E-cadherin and Rap1 may also exist in HSCs; combined with the
downregulation of N-cadherin, the decreased expression would
further enhance the error in the interaction with the
microenvironment, resulting in an accelerated cell cycle and
apoptosis, and eventually, the exhausted reservoir of HSCs and the
clinical manifestation of cytopenia.
The cadherins on the membrane must regulate
hematopoiesis through certain molecular pathways in which various
molecules are involved. We have shown that β-catenin expression was
higher in patients with MDS-RA. Constitutively active β-catenin in
transgenic mouse HSCs showed blocked multilineage differentiation
and reduced colony formation (20,50).
The analysis of genes involved in the Wnt signaling (including Wnt
ligand, Axin complex or Fz receptor complex and others) by chips,
and APC, Axin, CK1, DVL, GSK3, LRP5, LRP6 using RQ-PCR, did not
show changes in expression. No Wnt pathway mutations have been
detected in hematological malignancies thus far. Therefore,
cadherin may modulate β-catenin activity in HSCs with MDS as the
expression of cadherin decreased. N-cadherin shRNA and a weakened
interplay between β-catenin and cadherin by partially dismantled
AJs enhanced the nuclear accumulation of β-catenin (51–53),
which may be responsible for the accelerated cell division, reduced
long-term repopulation activity of HSCs and resulted in the final
exhaustion of the stem cell pool (20,50,54). A
combination of the downregulation of N-cadherin and E-cadherin and
the upregulation of β-catenin in MDS-RA samples suggests that more
β-catenin would accumulate in the cytoplasm and then be transported
to the nuclei of the HSCs. Continuous activation of β-catenin is
reported to block LT-HSC differentiation with the exception of
inducing the exhaustion of the stem cell pool (20,50),
resulting in the absence of mature terminal blood cells and the
final manifestation of peripheral cytopenia in MDS-RA.
In the nucleus, β-catenin interacts with T-cell
factor/Lef transcription factors and controls target gene
expression (25,55,56).
Among these genes, β-catenin is important in activating the
transcription of a proto-oncogene, c-myc (27) (Fig.
1). The activity of c-myc protein is required for primitive
hematopoiesis and for maintaining the proliferation of lineage
committed cell types in BM (15,57).
c-myc regulates the balance between self-renewal and
differentiation. The elimination of c-myc shifts the balance
towards self-renewal at the expense of differentiation, resulting
in the accumulation of LT-HSC, whereas committed progenitors and
differentiated cell types are lost. N-cadherin is upregulated in
stem cells in the absence of c-myc and this suggests that the
mutants fail to differentiate due to their inability to detach from
the differentiation preventive niche. The deregulated
overexpression of c-myc in HSCs promotes differentiation at the
expense of self-renewal and causes the repression of N-cadherin,
resulting in stem cell exhaustion and the absence of mature
terminal blood cells (15). In a
previous study, the c-myc protein was expressed at a higher level
in CD34+ cells with MDS (58). The results of the present study do
not show a difference in direct transcripts of c-myc in MDS, which
may be due to limited samples; however, two regulatory molecules
showed some notable changes. The c-myc binding protein, which
promotes the expression of c-myc, was downregulated, while the
c-myc promoter binding protein, which inhibits the expression of
c-myc, was upregulated. These findings may reflect that the
transcription level of c-myc tended to increase, resulting in a
relatively higher median value in patients with MDS-RA (Fig. 3). If the downregulation of cadherins
inhibits the self-renewal activity of HSCs, the balance may shift
towards differentiation, but may be blocked at terminal
differentiation, resulting in the loss of all hematopoietic cell
types over time due to a normal cell turnover in MDS. These
mechanisms may explain the major features of MDS, i.e., the
hypercellularity in BM, the ineffective hematopoiesis caused by an
accelerated apoptosis and the cytopenia in peripheral blood.
The nuclear localization of β-catenin is
indispensable for regulating target gene expression programs. Wu
et al reported that the nuclear accumulation of β-catenin
depends on its phosphorylation at Ser191 and Ser605, which requires
Rac1 activation (Fig. 1) (59). Rac1, a member gene of the Rho GTPase
family, was expressed at higher levels in the MDS-RA group of
CD34+ cells. Another two Rho GTPase genes, Rac2 and
CDC42, were also expressed at higher levels within the same study
group. Rac1 and Rac2 belong to the same subfamily Rac and increased
expression was in agreement with the suggestion of nuclear
accumulation of β-catenin in patients with MDS-RA. Members of the
Rho GTPase family are also involved with the cytoskeleton and cell
shape (28), for example, Rac1
plays significant roles in neutrophil shape and tail retraction
(60,61); Rac2 mainly contributes to actin
polymerization (60,62–65)
and CDC42 is involved in neutrophil polarity (66,67).
We believe that the aberrant expression of these RhoGTPase family
members in MDS-RA is relevant to the morphological dysplasia of BM
cells. The cytoskeleton, cadherins and catenins are closely linked
to affect the interaction between HSCs and their microenvironment
(Fig. 1). Rho GTPases, as key
mediators of the cytoskeleton, also affect hematopoiesis and
hemopathies. For example, Rac1 is essential for the engraftment of
HSCs into the BM (60), whereas
Rac2 is required for retention of HSCs in the BM (68,69)
and CDC42 uniquely regulates HSC trafficking and residence in the
BM niche (70,71). Deletion of Rac1, Rac2 and CDC42
produces similar effects to those observed in the downregulation of
cadherin and upregulation of β-catenin and c-myc (60,69,71).
As the Rac1, Rac2 and CDC42 genes were all highly expressed in
MDS-RA, their involvement in a compensation mechanism needs further
investigation. The function of these proteins should be elucidated
at the protein level, especially the active GTP-binding
molecules.
Therefore, based on the gene expression profiles
using CD34+ cells, we propose that the
cadherin-β-catenin-c-myc signaling axis may affect hematopoiesis in
early MDS (Fig. 1). We drew this
conclusion based on the interaction between HSCs and the
microenvironment. Raaijmakers et al (72) previously demonstrated that
genetically impaired mouse osteo-progenitor cells, as the component
of the niche, are likely to induce BM dysfunction similar to MDS.
These findings suggest that the perturbation of certain cells in
the microenvironment is not a passive event, but may be an early
event that triggers MDS initiation. Epigenetics has been considered
to be a unifying principle in the etiology of human complex traits
and diseases (73). Epigenetics
links the genotype of a cell to the phenotype under certain
environmental conditions. The epigenetic changes in a cell reflect
the crosstalk between the cells within their environment. In
comparison with genetic alterations, the epigenetic changes are
usually slow, small in size, dynamic and reversible and require
numerous cell divisions to be maintained. The early stage in the
formation of MDS clones may represent the procedure by which a
number of epigenetic changes occur in the healthy hematopoietic
stem/progenitor cells. This may be due to the dysfunctional change
of the niche or an impaired interaction between HSCs and the
microenvironment for unknown reasons. The abnormal HSCs with
epigenetic alterations, which may be viewed as MDS stem cells or
preleukemia stem cells, together with subsequent additional genetic
changes, progress and eventually become leukemia stem cells.
Accordingly, in the early stage of MDS, the abnormal MNCs with the
exception of CD34+ cells may all originate from the MDS
stem cells. As the CD34+ cells are extremely difficult
to obtain in early stage MDS, exploring the same gene expression
profiles based on MNCs may also provide further insight. Notably,
only Rap1GAP and Rac2 were expressed at higher levels in MNCs with
MDS-RA. There was no evidence of changes in gene expression
involved in the canonical Wnt signaling pathway and an increased
expression of Rap1GAP was observed not only in CD34+
cells, but also in MNCs. Therefore, it is likely that Rap1 acts as
an initiator for the cadherin-β-catenin-c-myc signaling axis via
the downregulated interplay between cadherin and Rap1 which has a
positive feedback mechanism. Rac2, which belongs to the same
subfamily of Rac1 of the Rho GTPase family, has the ability to
increase the nuclearization of β-catenin via its increased
activity. Therefore, the two small G proteins, Rap1GAP and Rac2,
may play pivotal roles in the initial development of MDS.
In conclusion, our expression data with regard to
the initiation and development of MDS may provide clues for the
elucidation of aberrant hematopoiesis in MDS. We also suggest early
events that may trigger the epigenetic alteration observed in HSCs.
This study provides new and significant insight into the
pathophysiology of MDS. In addition, Rap1GAP and Rac2 may act as
new molecular markers for the diagnosis of MDS.
Acknowledgements
This study was supported by the grant no. 81070402
from The National Natural Scientific Foundation of China and A
Project Funded by the Priority Academic Program Development of
Jiangsu Higher Education Institutions PAPD.
References
|
1
|
Corey SJ, Minden MD, Barber DL, Kantarjian
H, Wang JC and Schimmer AD: Myelodysplastic syndromes: the
complexity of stem-cell diseases. Nat Rev Cancer. 7:118–129. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nolte F and Hofmann WK: Myelodysplastic
syndromes: molecular pathogenesis and genomic changes. Ann Hematol.
87:777–795. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Spradling A, Drummond-Barbosa D and Kai T:
Stem cells find their niche. Nature. 414:98–104. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fuchs E, Tumbar T and Guasch G:
Socializing with the neighbors: Stem cells and their niche. Cell.
116:769–778. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Scadden DT: The stem-cell niche as an
entity of action. Nature. 441:1075–1079. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Muguruma Y, Yahata T, Miyatake H, Sato T,
Uno T, Itoh J, Kato S, Ito M, Hotta T and Ando K: Reconstitution of
the functional human hematopoietic microenvironment derived from
human mesenchymal stem cells in the murine bone marrow compartment.
Blood. 107:1878–1887. 2006. View Article : Google Scholar
|
|
7
|
Bendall SC, Stewart MH and Bhatia M: Human
embryonic stem cells: lessons from stem cell niches in vivo. Regen
Med. 3:365–376. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Calvi LM, Adams GB, Weibrecht KW, Weber
JM, Olson DP, Knight MC, Martin RP, Schipani E, Divieti P,
Bringhurst FR, et al: Osteoblastic cells regulate the
haematopoietic stem cell niche. Nature. 425:841–846. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang J, Niu C, Ye L, Huang H, He X, Tong
WG, Ross J, Haug J, Johnson T, Feng JQ, et al: Identification of
the haematopoietic stem cell niche and control of the niche size.
Nature. 425:836–841. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kiel MJ, Radice GL and Morrison SJ: Lack
of evidence that hematopoietic stem cells depend on
N-cadherin-mediated adhesion to osteoblasts for their maintenance.
Cell Stem Cell. 1:204–217. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lo Celso C, Fleming HE, Wu JW, Zhao CX,
Miake-Lye S, Fujisaki J, Côté D, Rowe DW, Lin CP and Scadden DT:
Live-animal tracking of individual haematopoietic stem/progenitor
cells in their niche. Nature. 457:92–96. 2009.PubMed/NCBI
|
|
12
|
Xie Y, Yin T, Wiegraebe W, He XC, Miller
D, Stark D, Perko K, Alexander R, Schwartz J, Grindley JC, et al:
Detection of functional haematopoietic stem cell niche using
real-time imaging. Nature. 457:97–101. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee H, Gaughan JP and Tsygankov AY: c-Cbl
facilitates cytoskeletal effects in v-Abl transformed fibroblast
through Rac1- and Rap1-mediated signaling. Int J Biochem Cell Biol.
40:1930–1943. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hosokawa K, Arai F, Yoshihara H, Iwasaki
H, Hembree M, Yin T, Nakamura Y, Gomei Y, Takubo K, Shiama H, et
al: Cadherin based adhesion is a potential target for niche
manipulation to protect hematopoietic stem cells in adult bone
marrow. Cell Stem Cell. 6:194–198. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wilson A, Murphy MJ, Oskarsson T, Kaloulis
K, Bettess MD, Oser GM, Pasche AC, Knabenhans C, Macdonald HR and
Trumpp A: c-Myc controls the balance between hematopoietic stem
cell self-renewal and differentiation. Genes Dev. 18:2747–2763.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Haug JS, He XC, Grindley JC, Wunderlich
JP, Gaudenz K, Ross JT, Paulson A, Wagner KP, Xie Y, Zhu R, et al:
N-cadherin expression level distinguishes reserved versus primed
states of hematopoietic stem cells. Cell Stem Cell. 2:367–379.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Turel KR and Rao SG: Expression of the
cell adhesion molecule E-cadherin by the human bone marrow stromal
cells and its probable role in CD34(+) stem cell adhesion. Cell
Biol Int. 22:641–648. 1998.PubMed/NCBI
|
|
18
|
Wang L, Menendez P, Cerdan C and Bhatia M:
Hematopoietic development from human embryonic stem cell lines. Exp
Hematol. 33:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li L, Wang S, Jezierski A, Moalim-Nour L,
Mohib K, Parks RJ, Retta SF and Wang L: A unique interplay between
Rap1 and E-cadherin in the endocytic pathway regulates self-renewal
of human embryonic stem cells. Stem Cells. 28:247–257.
2010.PubMed/NCBI
|
|
20
|
Kirstetter P, Anderson K, Porse BT,
Jacobsen SE and Nerlov C: Activation of the canonical Wnt pathway
leads to loss of hematopoietic stem cell repopulation and
multilineage differentiation block. Nat Immunol. 7:1048–1056. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Maher MT, Flozak AS, Stocker AM, Chenn A
and Gottardi CJ: Activity of the beta-catenin phosphodestruction
complex at cell-cell contacts is enhanced by cadherin-based
adhesion. J Cell Biol. 186:219–228. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nagafuchi A and Takeichi M: Cell binding
function of E-cadherin is regulated by the cytoplasmic domain. EMBO
J. 7:3679–3684. 1988.PubMed/NCBI
|
|
23
|
Meng W and Takeichi M: Adherens junction:
molecular architecture and regulation. Cold Spring Harb Perspect
Biol. 1:a0028992009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Stepniak E, Radice GL and Vasioukhin V:
Adhesive and signaling functions of cadherins and catenins in
vertebrate development. Cold Spring Harb Perspect Biol.
1:a0029492009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: components, mechanisms, and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Doucas H, Garcea G, Neal CP, Manson MM and
Berry DP: Changes in the Wnt signaling pathway in gastrointestinal
cancers and their prognostic significance. Eur J Cancer.
41:365–379. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Machacek M, Hodgson L, Welch C, Elliott H,
Pertz O, Nalbant P, Abell A, Johnson GL, Hahn KM and Danuser G:
Coordination of Rho GTPase activities during cell protrusion.
Nature. 461:99–103. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mulloy JC, Cancelas JA, Filippi MD, Kalfa
TA, Guo F and Zheng Y: Rho GTPases in hematopoiesis and
hemopathies. Blood. 115:936–947. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hofmann WK, de Vos S, Komor M, Hoelzer D,
Wachsman W and Koeffler HP: Characterization of gene expression of
CD34+ cells from normal and myelodysplastic bone marrow.
Blood. 100:3553–3560. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hosokawa K, Arai F, Yoshihara H, Nakamura
Y, Gomei Y, Iwasaki H, Miyamoto K, Shima H, Ito K and Suda T:
Function of oxidative stress in the regulation of hematopoietic
stem cell-niche interaction. Biochem Biophys Res Commun.
363:578–583. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pyle AD, Lock LF and Donovan PJ:
Neurotrophins mediate human embryonic stem cell survival. Nat
Biotechnol. 24:344–350. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Watanabe K, Ueno M, Kamiya D, Nishiyama A,
Matsumura M, Wataya T, Takahashi JB, Nishikawa S, Nishikawa S,
Muguruma K and Sasai Y: A ROCK inhibitor permits survival of
dissociated human embryonic stem cells. Nat Biotechnol. 25:681–686.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li X, Bryant CE and Deeg HJ: Simultaneous
demonstration of clonal chromosome abnormalities and apoptosis in
individual marrow cells in myelodysplastic syndrome. Int J Hematol.
80:140–145. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang YY, Cen JN, He J, Shen HJ, Liu DD,
Yao L, Qi XF and Chen ZX: Accelerated cellular senescence in
myelodysplastic syndrome. Exp Hematol. 37:1310–1317. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Balzac F, Avolio M, Degani S, Kaverina I,
Torti M, Silengo L, Small JV and Retta SF: E-cadherin endocytosis
regulates the activity of Rap1: a traffic light GTPase at the
crossroads between cadherin and integrin function. J Cell Sci.
118:4765–4783. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Asha H, de Ruiter ND, Wang MG and
Hariharan IK: The Rap1 GTPase functions as a regulator of
morphogenesis in vivo. EMBO J. 18:605–615. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hariharan IK, Carthew RW and Rubin GM: The
Drosophila roughened mutation: activation of a rap homolog
disrupts eye development and interferes with cell determination.
Cell. 67:717–722. 1991.
|
|
39
|
Ohba Y, Ikuta K, Ogura A, Matsuda J,
Mochizuki N, Nagashima K, Kurokawa K, Mayer BJ, Maki K, Miyazaki J
and Matsuda M: Requirement for C3G-dependent Rap1 activation for
cell adhesion and embryogenesis. EMBO J. 20:3333–3341. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bos JL: Linking Rap to cell adhesion. Curr
Opin Cell Biol. 17:123–128. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Knox AL and Brown NH: Rap1 GTPase
regulation of adherens junction positioning and cell adhesion.
Science. 295:1285–1288. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sebzda E, Bracke M, Tugal T, Hogg N and
Cantrell DA: Rap1A positively regulates T cells via integrin
activation rather than inhibiting lymphocyte signaling. Nat
Immunol. 3:251–258. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
43
|
Price LS, Hajdo-Milasinovic A, Zhao J,
Zwartkruis FJ, Collard JG and Bos JL: Rap1 regulates
E-cadherin-mediated cell-cell adhesion. J Biol Chem.
279:35127–35132. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hogan C, Serpente N, Cogram P, Hosking CR,
Bialucha CU, Feller SM, Braga VM, Birchmeier W and Fujita Y: Rap1
regulates the formation of E-cadherin-based cell-cell contacts. Mol
Cell Biol. 24:6690–6700. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bos JL: All in the family? New insights
and questions regarding interconnectivity of Ras, Rap1 and Ral.
EMBO J. 17:6776–6782. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Quilliam LA, Rebhun JF and Castro AF: A
growing family of guanine nucleotide exchange factors is
responsible for activation of Ras-family GTPases. Prog Nucleic Acid
Res Mol Biol. 71:391–444. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bos JL, Rehmann H and Wittinghofer A: GEFs
and GAPs: critical elements in the control of small G proteins.
Cell. 129:865–877. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Qi X, Chen Z, Qian J, Cen J and Gu M:
Expression of Rap1GAP in human myeloid disease following microarray
selection. Genet Mol Res. 7:379–387. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ika SA, Qi XF and Chen ZX: Protein RAP1GAP
in human myelodysplastic syndrome detected by flow cytometry and
its clinical relevance. Zhongguo Shi Yan Xue Ye Xue Za Zhi.
17:612–617. 2009.PubMed/NCBI
|
|
50
|
Scheller M, Huelsken J, Rosenbauer F,
Taketo MM, Birchmeier W, Tenen DG and Leutz A: Hematopoietic stem
cell and multilineage defects generated by constitutive
beta-catenin activation. Nat Immunol. 7:1037–1047. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hosokawa K, Arai F, Yoshihara H, Iwasaki
H, Nakamura Y, Gomei Y and Suda T: Knockdown of N-cadherin
suppresses the long-term engraftment of hematopoietic stem cells.
Blood. 116:554–563. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Taddei A, Giampietro C, Conti A, Orsenigo
F, Breviario F, Pirazzoli V, Potente M, Daly C, Dimmeler S and
Dejana E: Endothelial adherens junctions control tight junctions by
VE-cadherin-mediated upregulation of claudin-5. Nat Cell Biol.
10:923–934. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Heuberger J and Birchmeier W: Interplay of
cadherin-mediated cell adhesion and canonical Wnt signaling. Cold
Spring Harb Perspect Biol. 2:a0029152010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kiel MJ, Yilmaz OH, Iwashita T, Terhorst C
and Morrison SJ: SLAM family receptors distinguish hematopoietic
stem and progenitor cells and reveal endothelial niches for stem
cells. Cell. 121:1109–1121. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Frank SR, Schroeder M, Fernandez P,
Taubert S and Amati B: Binding of c-Myc to chromatin mediates
mitogen-induced acetylation of histone H4 and gene activation.
Genes Dev. 15:2069–2082. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Staller P, Peukert K, Kiermaier A, Seoane
J, Lukas J, Karsunky H, Möröy T, Bartek J, Massagué J, Hänel F and
Eilers M: Repression of p15INK4b expression by Myc through
association with Miz-1. Nat Cell Biol. 3:392–399. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Trumpp A, Refaeli Y, Oskarsson T, Gasser
S, Murphy M, Martin GR and Bishop JM: c-Myc regulates mammalian
body size by controlling cell number but not cell size. Nature.
414:768–773. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Rajapaksa R, Ginzton N, Rott LS and
Greenberg PL: Altered oncoprotein expression and apoptosis in
myelodysplastic syndrome marrow cells. Blood. 88:4275–4287.
1996.PubMed/NCBI
|
|
59
|
Wu X, Tu X, Joeng KS, Hilton MJ, Williams
DA and Long F: Rac1 activation controls nuclear localization of
beta-catenin during canonical Wnt signaling. Cell. 133:340–353.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Gu Y, Filippi MD, Cancelas JA, Siefring
JE, Williams EP, Jasti AC, Harris CE, Lee AW, Prabhakar R, Atkinson
SJ, et al: Hematopoietic cell regulation by Rac1 and Rac2 guanosine
triphosphatases. Science. 302:445–449. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Filippi MD, Szczur K, Harris CE and
Berclaz PY: Rho GTPase Rac1 is critical for neutrophil migration
into the lung. Blood. 109:1257–1264. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Roberts AW, Kim C, Zhen L, Lowe JB, Kapur
R, Petryniak B, Spaetti A, Pollock JD, Borneo JB, Bradford GB, et
al: Deficiency of the hematopoietic cell-specific Rho family GTPase
Rac2 is characterized by abnormalities in neutrophil function and
host defense. Immunity. 10:183–196. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Sun CX, Downey GP, Zhu F, Koh AL, Thang H
and Glogauer M: Rac1 is the small GTPase responsible for regulating
the neutrophil chemotaxis compass. Blood. 104:3758–3765. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Szczur K, Xu H, Atkinson S, Zheng Y and
Filippi MD: Rho GTPase CDC42 regulates directionality and random
movement via distinct MAPK pathways in neutrophils. Blood.
108:4205–4213. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Szczur K, Zheng Y and Filippi MD: The
small Rho GTPase Cdc42 regulates neutrophil polarity via CD11b
integrin signaling. Blood. 114:4527–4537. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Pestonjamasp KN, Forster C, Sun C,
Gardiner EM, Bohl B, Weiner O, Bokoch GM and Glogauer M: Rac1 links
leading edge and uropod events through Rho and myosin activation
during chemotaxis. Blood. 108:2814–2820. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ferguson GJ, Milne L, Kulkarni S, Sasaki
T, Walker S, Andrews S, Crabbe T, Finan P, Jones G, Jackson S, et
al: PI(3)Kgamma has an important context-dependent role in
neutrophil chemokinesis. Nat Cell Biol. 9:86–91. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yang FC, Atkinson SJ, Gu Y, Borneo JB,
Roberts AW, Zheng Y, Pennington J and Williams DA: Rac and Cdc42
GTPases control hematopoietic stem cell shape, adhesion, migration,
and mobilization. Proc Natl Acad Sci USA. 98:5614–5618. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Cancelas JA, Lee AW, Prabhakar R, Stringer
KF, Zheng Y and Williams DA: Rac GTPases differentially integrate
signals regulating hematopoietic stem cell localization. Nat Med.
11:886–891. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wang L, Yang L, Filippi M, Williams D and
Zheng Y: Genetic deletion of Cdc42GAP reveals a role of Cdc42 in
erythropoiesis and hematopoietic stem/progenitor cell survival,
adhesion, and engraftment. Blood. 107:98–105. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Yang L, Wang L, Geiger H, Cancelas JA, Mo
J and Zheng Y: Rho GTPase Cdc42 coordinates hematopoietic stem cell
quiescence and niche interaction in the bone marrow. Proc Natl Acad
Sci USA. 104:5091–5096. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Raaijmakers MH, Mukherjee S, Guo S, Zhang
S, Kobayashi T, Schoonmaker JA, Ebert BL, Al-Shahrour F, Hasserjian
RP, Scadden EO, et al: Bone progenitor dysfunction induces
myelodysplasia and secondary leukemia. Nature. 464:852–857. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Petronis A: Epigenetics as a unifying
principle in the aetiology of complex traits and diseases. Nature.
465:721–727. 2010. View Article : Google Scholar : PubMed/NCBI
|















