Introduction
Colorectal cancer (CRC) is one of the most common
malignant tumors, with 1,200,000 new cases and 608,700 mortalities
worldwide annually (1);
furthermore, it is the fourth leading cause of cancer-related
mortality in China. In recent decades, the incidence of CRC has
increased. Despite surgical resection coupled with systemic
chemotherapy, approximately half of newly diagnosed CRC patients
are likely to succumb to the disease due to tumor recurrence and
metastasis (2). The molecular
mechanism leading to the development, local invasion and distal
metastasis of CRC remains unclear. Thus, the delineation of the
mechanisms for colorectal carcinogenesis is of importance, as it
may provide novel strategies for diagnosis and prognosis.
Deleted in liver cancer 2 (DLC2), also called STARD
13, has been identified as a tumor suppressor gene and is closely
related to deleted in liver cancer 1 (DLC1). DLC2 is located at
chromosome 13q12.3 (3). DLC1 and
DLC2 proteins share 51% identity and 64% similarity at the level of
their amino acid sequences. DLC2 protein contains multiple domains,
including a sterile α motif (SAM), a RhoGAP and a lipid-binding
StAR-related lipid-transfer (START) domain. Previous studies have
shown that DLC2 suppressed cytoskeleton reorganization, cell
growth, cell migration and transformation in hepatocellular
carcinoma (HCC) (4); the START
domain of DLC2 has been reported to be involved in mitochondrial
localization in hepatoma cells (5)
and our previous study indicated that patients with DLC2
underexpression showed a significantly poorer prognosis than those
with high DLC2 expression in HCC (6). To date, all studies examining DLC2
expression and its suppressed function have been conducted on HCC.
However, DLC2 mRNA has been found to be downregulated in a
wide range of cancers, including CRC (7), but there have been no available data
on the expression levels of DLC2 mRNA and protein in
clinical CRC, especially in Chinese patients.
Therefore, in this study we examined the DLC2
mRNA and protein expression in Chinese patients with CRC and
evaluated the correlations between DLC2 expression levels and
clinicopathological parameters of CRC.
Materials and methods
Tissue specimens
The study protocol was approved by the Ethics
Committee of the Central South University (CSU). Fresh samples of
CRC tissues and pericarcinomatous intestine tissues (PCITs, 2.0 cm
away from the carcinoma) were obtained from 102 (66 males and 36
females) Chinese patients with primary cancer who underwent surgery
at The Third Affiliated Hospital of Central South University
between Octorber 2010 and February 2012. Each specimen was divided
into two parts; one was immediately frozen in liquid nitrogen and
stored at −80°C for real-time PCR and western blot analysis, the
other was fixed in 4% paraformaldehyde for immunohistochemistry.
The median age of the patients was 55.43 years, ranging from 26 to
78 years. All specimens obtained from surgical resection were
confirmed by pathological examination. Written informed consent was
obtained from each patient prior to the study.
Real-time PCR
Total RNA was extracted from specimens using TRIzol
reagent (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer’s instructions. Aliquots of 5 μg RNA from each
sample were reverse transcribed. Quantitative analysis of
DLC2 mRNA expression using the SYBR Green method was
performed using the ABI 7300 real-time PCR system. The sequences of
the primers were as follows: DLC2 5′-GTCAATGGGCTCCAAGAGGTAG-3′
(forward) and 5′-TGATTAGGAGATGGAAAGGTGGA-3′ (reverse); GAPDH
(endogenous control): 5′-CTGCACCACCAACTG CTTAG-3′ (forward);
5′-AGGTCCACCACTGACACGTT-3′ (reverse). A 25-μl reaction
mixture containing 1 μl cDNA template, 12.5 μl SYBR
Master mix and 0.20 μl of each primer was amplified using
the following thermal parameters: denaturing at 95˚C for 10 min and
45 cycles of the amplification step (denaturation at 95˚C for 15
sec, annealing and extension at 60˚C for 1 min). All amplification
reactions were performed in triplicate and the averages of the
threshold cycles (Cts) were used to interpolate curves using 7300
System SDS software. The results were expressed as the ratio of
DLC2 mRNA to GAPDH mRNA, and the results of each
sample was compared against one control sample to eliminate the
differences between the PCR plates.
Western blot analysis
Protein was extracted from the specimens using the
fierce RIPA lysis buffer (Beyotime Biotech, Jiangsu, China)
following the manufacturer's instructions. Briefly, an equal amount
(35 μg) of the total protein was separated by
SDS-polyacrylamide gel electrophoresis (SDS-PAGE; Beyotime Biotech)
and then transferred onto a polyvinylidene difluoride (PVDF; Pall,
NY, USA) membrane using standard procedures. The membranes were
immunoblotted with antibodies against DLC2 (rabbit polyclonal
antibody; Sigma, St. Louis, MO, USA), followed by a horseradish
peroxidase-conjugated secondary antibody. The washes were repeated
before the membranes were detected using ECL system (KPL
Biosciences, Gaithesburg, MD, USA). Band intensities were analyzed
densitometrically using the Image J software.
Immunohistochemistry
All the specimens were immunohistochemically
evaluated. In brief, tissue sections of 3-μm thickness were
cut and baked at 60˚C, deparaffinized in xylene and rehydrated
through graded ethanol. Next, 3% hydrogen peroxide and methanol
were applied to block the endogenous peroxidases and sections were
subjected to heat-induced antigen retrieval in 0.01 M citrate
buffer (pH=6.0). The samples were incubated overnight with primary
antibodies against DLC2 at a 1:400 dilution (rabbit polyclonal
antibody; Sigma). Then, the samples were treated with MaxVision TM
HRP-Polymer anti-Mouse/Rabbit IHC kit (KIT-5020, MaxVision,
Shenzen, China) and 3,3′-diaminobenzidine tetrahydrochloride (DAB).
Counterstaining was performed with 0.5% hematoxylin.
Statistical analysis
Statistical analysis was performed using SPSS
software (version 17.0; SPSS, Inc., Chicago, IL, USA). Data are
expressed as mean ± SD and, for clarity, as mean ± SEM in figures.
The Student's t-test was used to analyze the correlation betwen the
expression of DLC2 in CRC and PCIT. The non-parametric Mann-Whitney
U test was used to analyze the correlations between the expression
of DLC2 and clinicopathological variables. P<0.05 was considered
to indicate a statistically significant result.
Results
Expression of DLC2 mRNA in CRC and
PCIT
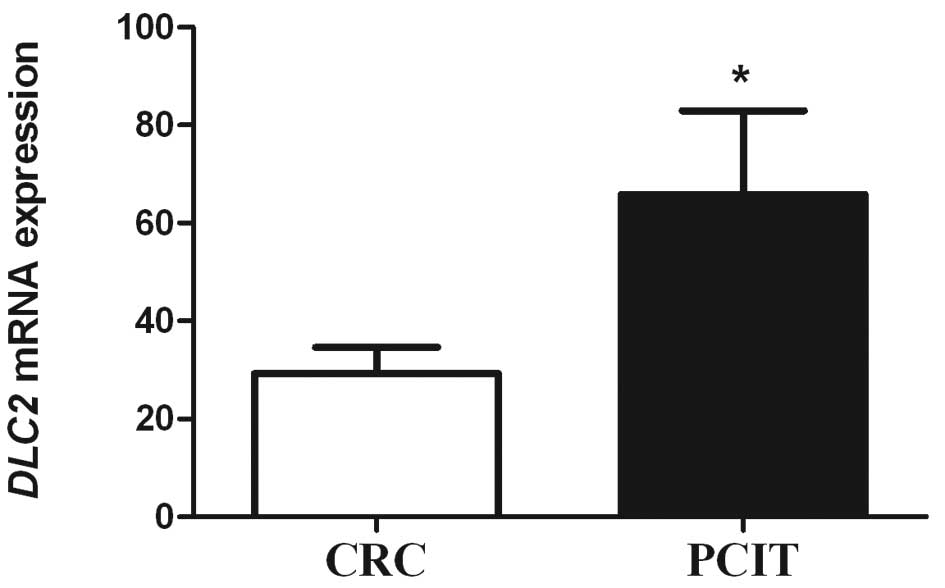
The expression of DLC2 mRNA was detected in
all CRC and PCIT specimens using real-time PCR. The relative
expression of DLC2 mRNA in CRC and PCIT was 29.16±5.36 and
65.87±17.05, respectively. The mRNA expression in CRC tissues was
significantly lower than that in PCIT (P=0.037; Fig. 1).
Expression of DLC2 protein in CRC and
PCIT
The protein expression was detected in all CRC and
PCIT specimens using western blotting. The relative expression of
DLC2 protein in CRC and PCIT was 2.29±0.61 and 1.66±0.65,
respectively. There was no significant difference between CRC and
PCIT. In addition, to identify the DLC2 protein expression status,
immunohistochemical staining was also employed. The result was
consistent that of the western blot analysis (Fig. 2).
Correlations between DLC2 expression
levels and clinicopathological parameters of CRC
The distribution pattern of DLC2 was determined by
dividing the expression levels of DLC2 mRNA and protein in
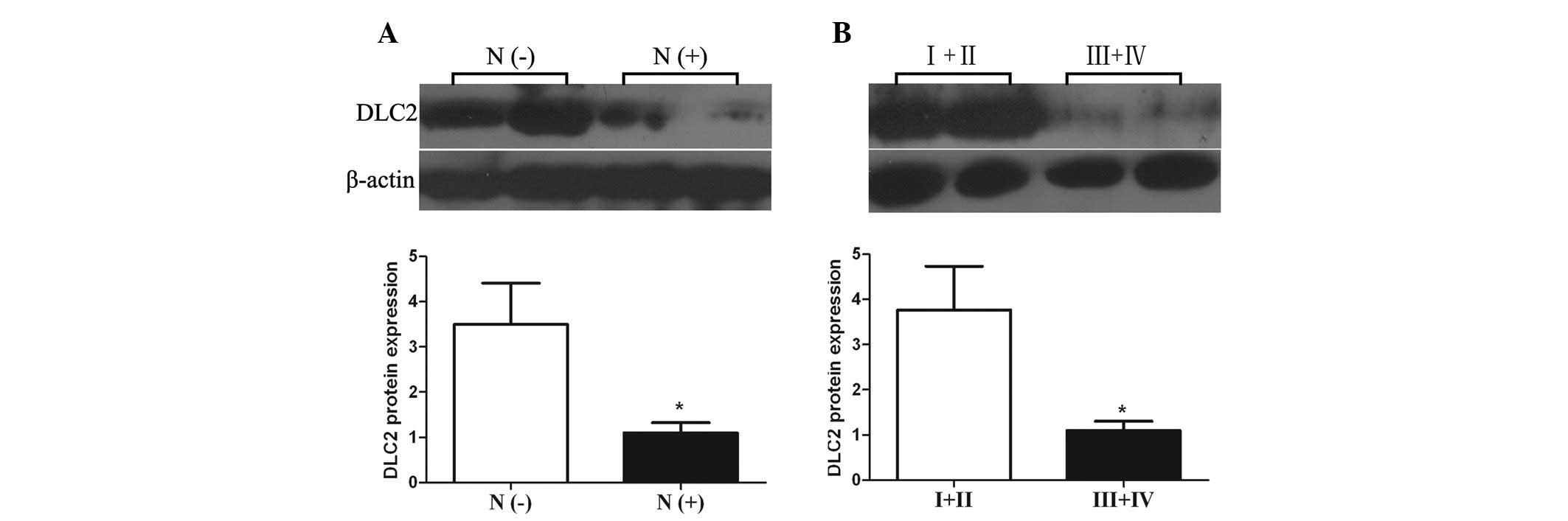
the subgroups by clinicopathological parameters. A significant
difference was found between DLC2 expression levels in CRC with or
without lymph node metastasis (P= 0.021 and 0.028, respectively);
both DLC2 mRNA and protein expression levels in CRC with
lymph node metastasis were lower than those in CRC cases without
lymph node metastasis (Figs. 3A and
4A). DLC2 mRNA and protein
expression levels of CRC of tumor TNM stage III and IV also showed
lower expression levels than in CRC cases of stage I and II
(P=0.020 and 0.026, respectively; Figs.
3B and 4B). In addition, the
expression status of DLC2 mRNA was also correlated with
tumor histopathological degree; poorly differentiated CRC cases had
a lower expression level of DLC2 than well- and moderately
differentiated CRC (P= 0.004; Fig.
5). However, there was no significant association between the
expression levels of DLC2 and other clinicopathological parameters,
including gender, age, tumor size, tumor site, depth of cancer
invasion, distant metastasis and blood levels of carcinoembryonic
antigen (CEA; Table I).
 | Table ICorrelation between expression levels
of DLC2 and clinicopathological parameters. |
Table I
Correlation between expression levels
of DLC2 and clinicopathological parameters.
| Variable | No. of
patients | P-value
|
|---|
| DLC2
mRNA | DLC2 protein |
|---|
| Gender | | 0.429 | 0.288 |
| Male | 66 | | |
| Female | 36 | | |
| Age (years) | | 0.625 | 0.420 |
| <60 | 60 | | |
| ≥60 | 42 | | |
| Tumor site | | 0.528 | 0.918 |
| Colon | 45 | | |
| Rectum | 57 | | |
| Tumor size
(cm) | | 0.457 | 0.425 |
| <5.0 | 52 | | |
| ≥5.0 | 50 | | |
| Tumor depth | | 0.876 | 0.354 |
| Limited to
muscular layer | 66 | | |
| Serosa and
adjacent fat | 36 | | |
| Lymph node
metastasis | | 0.021a | 0.028a |
| Negative | 50 | | |
| Positive | 52 | | |
| Distant
metastasis | | 0.338 | 0.233 |
| Absent | 94 | | |
| Present | 8 | | |
| Tumor TNM
stage | | 0.020a | 0.026a |
| I and II | 49 | | |
| III and IV | 53 | | |
| Tumor
differentiation | | 0.004a | 0.080 |
|
Well-moderate | 76 | | |
| Poor | 26 | | |
| CEA (ng/ml) | | 0.903 | 0.271 |
| ≥5 | 41 | | |
| <5 | 61 | | |
Discussion
Although DLC2 has been identified as a tumor
suppressor gene, the expression levels of DLC2 mRNA and
protein, especially in clinical CRC of Chinese patients, have not
been studied. The present study indicates that the expression level
of DLC2 mRNA was significantly lower in CRC than in PCIT;
however, the protein expression level of DLC2 was not significantly
different in CRC compared with PCIT in Chinese patients. This
difference between mRNA and protein may be due to
post-transcriptional control. With regard to human neoplasia,
Ullmannova and Popescu reported that the DLC2 mRNA level was
downregulated in lung, ovarian, renal, breast, uterine, gastric,
colon and rectal tumors and that the expression ratios of
DLC2 in colon and rectal cancer were 46 and 61%,
respectively (7). Our real-time PCR
results showed that DLC2 mRNA was also underexpressed and
the expression ratios of DLC2 mRNA were 70 and 62% in colon
and rectal cancer, respectively. This result is higher than that
reported by Ullmannova and Popescu; the reason for this difference
may be differences between ethnicities. Also, the low expression of
DLC2 has been confirmed to be involved in the invasion and
migration of HCC cells (4). DLC2
protein has been reported to inhibit the activity of
Raf-1-ERK1/2-p70S6K via its RhoGAP function, resulting in the
suppression of cell growth (8).
DLC2 has been found to modulate angiogenic responses in vascular
endothelial cells by regulating cell attachment and migration
(9). Consistent with previous
studies, the present study showed that DLC2 mRNA was
downregulated in CRC, which indicates that DLC2 participates in
colorectal carcinogenesis and plays an important role in the
invasion and metastasis of CRC.
To investigate how DLC2 affects colorectal
carcinogenesis, we evaluated the correlations between DLC2
expression levels, including mRNA and protein, and
clinicopathological parameters of CRC. Our results revealed that
the expression levels of DLC2 mRNA and protein were
frequently with lymph node metastasis and the DLC2 expression
levels in CRC with lymph node metastasis were lower than those in
CRC without lymph node metastasis. This result indicates that DLC2
is a tumor suppressor, whose downregulated expression stimulates
the lymph node metastasis of CRC. This implies that DLC2 has a role
in suppressing CRC lymph node metastasis. Our previous study showed
that a lower expression level of DLC2 was associated with higher
expression level of RhoA in HCC (6). In addition, other studies have shown
that DLC2 exhibited its tumor suppressor functions by means of the
inhibition of RhoA activity (4); a
higher level expression of RhoA has been detected in several types
of cancer, including bladder, testicular, ovarian, colon, breast
and lung cancer (10–13); Takami et al demonstrated that
the activity of RhoA was correlated with lymph node metastasis in
human CRC (14). Accordingly, we
hypothesized that the downregulation of DLC2 and upregulation of
RhoA may be essential in CRC progression, especially in the process
of CRC lymph node metastasis. To confirm this hypothesis,
investigations into the correlation between DLC2 and RhoA are
anticipated.
Notably, in the present study, the mRNA and protein
expression levels of DLC2 were also correlated with CRC tumor TNM
stage. Our results revealed that the DLC2 mRNA and protein
expression levels in CRC of TNM stages III and IV were lower than
those in CRCs of stage I and II. This result implies that
downregulated mRNA and protein expression levels of DLC2 were
correlated with advanced stage of CRC. Additionally, the mRNA
expression level was correlated with tumor histopathological degree
of CRC. It is well known that the TNM staging system of tumors is
the gold standard for evaluating prognosis in patients with CRC. It
has been reported that the five-year survival rates of CRC patients
who underwent surgical resections were 94.1, 80.2, 61.7 and 23.2%
with tumors of TNM stages I to IV, respectively (15); additionally, low tumor
histopathological differentiation was also associated with poor
survival (16). Thus, it is
reasonable to assume that the downregulated expression of DLC2 is
correlated with a poor prognosis of CRC patients. To test this
assumption, we intend to continue the follow-up of these patients
in our further research.
To the best of our knowledge, this is the first
study to report DLC2 mRNA and protein expression levels in
Chinese CRC patients and the downregulated expression levels of
DLC2 mRNA and protein are associated with lymph node
metastasis and tumor TNM stage in CRC. The DLC2 mRNA
expression level is also correlated with tumor histopathological
degree. Although a number of diagnostic advances have been made, it
remains difficult to accurately diagnose lymph nodes metastasis and
assess the prognosis of CRC. In this study, we found that the
downregulated expression levels of DLC2 mRNA and protein
were correlated with lymph node metastasis and tumor TNM stage.
Furthermore, the underexpression of DLC2 mRNA was also
associated with tumor histopathological degree of CRC. Therefore,
the present study demonstrates that DLC2 may not only participate
in CRC carcinogenesis, invasion and lymph node metastasis, but may
also be a potential indicator for the presence of lymph nodes
metastasis in CRC and a prognostic marker for CRC patients.
Acknowledgements
This study was supported by the
Planned Science and Technology Project of Hunan Province, China
(no. 22010FJ3087) and the Chinese National Science Foundation (no.
81172298). We would like to thank Dr Ren Guo for assistance with
experiments and manuscript reviewing and formatting.
References
|
1.
|
A JemalF BrayMM CenterJ FerlayE WardD
FormanGlobal cancer statisticsCA Cancer J
Clin616990201110.3322/caac.20107
|
|
2.
|
HJ WilkeE Van CutsemCurrent treatments and
future perspectives in colorectal and gastric cancerAnn
Oncol14Suppl 2ii49ii55200310.1093/annonc/mdg73012810459
|
|
3.
|
YP ChingCM WongSF ChanDeleted in liver
cancer (DLC) 2 encodes a RhoGAP protein with growth suppressor
function and is underexpressed in hepatocellular carcinomaJ Biol
Chem2781082410830200310.1074/jbc.M20831020012531887
|
|
4.
|
TH LeungYP ChingJW YamDeleted in liver
cancer 2 (DLC2) suppresses cell transformation by means of
inhibition of RhoA activityProc Natl Acad Sci
USA1021520715212200510.1073/pnas.050450110216217026
|
|
5.
|
DC NgSF ChanKH KokMitochondrial targeting
of growth suppressor protein DLC2 through the START domainFEBS
Lett580191198200610.1016/j.febslet.2005.11.07316364308
|
|
6.
|
L XiaorongW WeiQ LiyuanY
KaiyanUnderexpression of deleted in liver cancer 2 (DLC2) is
associated with overexpression of RhoA and poor prognosis in
hepatocellular carcinomaBMC
Cancer8205200810.1186/1471-2407-8-20518651974
|
|
7.
|
V UllmannovaNC PopescuExpression profile
of the tumor suppressor genes DLC-1 and DLC-2 in solid tumorsInt J
Oncol2911271132200617016643
|
|
8.
|
TH LeungJW YamLK ChanYP ChingIO NgDeleted
in liver cancer 2 suppresses cell growth via the regulation of the
Raf-1-ERK1/2-p70S6K signalling pathwayLiver
Int3013151323201010.1111/j.1478-3231.2010.02307.x20629949
|
|
9.
|
Y LinNT ChenYP ShihYC LiaoL XueSH LoDLC2
modulates angiogenic responses in vascular endothelial cells by
regulating cell attachment and
migrationOncogene2930103016201010.1038/onc.2010.5420208559
|
|
10.
|
AB JaffeA HallRho GTPases in
transformation and metastasisAdv Cancer
Res845780200210.1016/S0065-230X(02)84003-911883532
|
|
11.
|
T KamaiK AraiT TsujiiM HondaK
YoshidaOverexpression of RhoA mRNA is associated with advanced
stage in testicular germ cell tumourBJU
Int87227231200110.1046/j.1464-410x.2001.02030.x11167647
|
|
12.
|
G FritzI JustB KainaRho GTPases are
over-expressed in human tumorsInt J
Cancer81682687199910.1002/(SICI)1097-0215(19990531)81:5%3C682::AID-IJC2%3E3.0.CO;2-B10328216
|
|
13.
|
T KamaiT TsujiiK AraiSignificant
association of Rho/ROCK pathway with invasion and metastasis of
bladder cancerClin Cancer Res926322641200312855641
|
|
14.
|
Y TakamiM HigashiS KumagaiThe activity of
RhoA is correlated with lymph node metastasis in human colorectal
cancerDig Dis Sci53467473200810.1007/s10620-007-9887-017597401
|
|
15.
|
Y LuoJ CuiC ChenClinical outcomes after
surgical resection of colorectal cancer in 1,294
patientsHepatogastroenterology5913981402201122115801
|
|
16.
|
S GillCL LoprinziDJ SargentPooled analysis
of fluorouracil-based adjuvant therapy for stage II and III colon
cancer: who benefits and by how much?J Clin
Oncol2217971806200410.1200/JCO.2004.09.05915067028
|