Introduction
Gastric cancer is the fourth most common type of
cancer and the second leading cause of cancer-related mortality in
the world (1). Chemotherapy plays a
significant role in the treatment of gastric cancer in adjuvant and
advanced settings. 5-fluorouracil (5-FU) and its derivatives (e.g.,
S-1 and Xeloda) are significant, effective agents used to treat
gastric cancer. However, the efficacy of chemotherapy for gastric
cancer is limited due to insensitivity and the development of drug
resistance.
To reveal the mechanisms underlying 5-FU resistance,
the proteomic profiles of 5-FU-resistant and -sensitive colon
cancer cells have been previously compared by 2-D gel
electrophoresis. It was revealed that RhoGDI2 was upregulated in
5-FU-resistant colon cancer cells (LoVo/5-FU) and that the
knockdown of RhoGDI2 expression by transfection with
RhoGDI2-specific siRNA significantly increased the sensitivity to
5-FU in LoVo/5-FU. These data suggest that RhoGDI2 confers
resistance to 5-FU in colon cancer cells.
RhoGDI2 belongs to a family of Rho GTPase dissociate
inhibitors (GDIs). GDIs are pivotal regulators of Rho GTPase
function typified by forming a complex with Rho GTPase, modulating
their nucleotide exchange and membrane association. Therefore, they
play a significant role in regulating the actin cytoskeleton, cell
polarity, microtubule dynamics, membrane transport pathways and
transcription factor activity (2,3).
Unlike other members of the family (such as RhoGDI1 and RhoGDI3),
RhoGDI2 is preferentially expressed in hematopoietic cells and
appears to have a narrow selectivity and lower binding affinity for
Rho GTPases (4). RhoGDI2 associates
with and negatively regulates Rac1 and Rac3 in breast cancer cells,
but not RhoA, Cdc42 and RhoC (5),
whereas it positively regulates Rac1 in human bladder cancer cells
(6). RhoGDI2 is also a substrate
for caspases and becomes cleaved in various cell types during
apoptosis. The significant role of RhoGDI2 in cancer has been
previously noted by several lines of study. RhoGDI2 expression
appears to be inversely correlated with an invasive capacity in
bladder cancer cell lines (7). The
reduced expression of RhoGDI2 protein was associated with a poor
prognosis of patients with advanced bladder cancer (8). By contrast, RhoGDI2 is overexpressed
in human breast cancer cell lines and increases cancer cell
invasion and motility in vitro(9). Choi et al also revealed that
RhoGDI2 expression is positively correlated with tumor progression
and metastasis potential in gastric cancer (10). Therefore, exploring the role of
RhoGDI2 in gastric cancer is likely to aid our understanding of how
it contributes to 5-FU resistance.
Materials and methods
Cell culture
The human gastric cancer cell lines SNU16, SNU-1,
AGS and KATO-III were obtained from the American Type Culture
Collection (Rockville, MD, USA). The human gastric cancer cell
lines MKN-45, MKN-28 and SGC7901 were obtained from the Chinese
Academy of Sciences Cell Bank of Type Culture Collection (Shanghai,
China). All the gastric cancer cell lines were cultured in
RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with
10% FBS (PAA Laboratories GmbH, Morningside, Queensland,
Australia).
Patients and tissues
Human gastric cancer tissue arrays were obtained
from Outdo Biotech (Shanghai, China) with 88 individual cases of
gastric cancer and matched normal colon tissue. For the in
vitro chemosensitivity test, 12 tumor specimens were obtained
from patients during primary surgery from the Department of Surgery
at Ruijin Hospital (Shanghai, China). All tissues were obtained
from untreated patients following informed consent. The study was
approved by the Ethics Committee of Ruijin Hospital, Shanghai
JiaoTong University School of Medicine, Shanghai, China. Certain
sections of the specimens were transfered directly into tissue
culture medium for cell culture, while the others were fixed with
formalin for the IHC test.
RhoGDI2-expressing plasmid
To generate a RhoGDI2 expression plasmid, the
full-length coding region of RhoGDI2 cDNA was amplified using the
primers RhoGDI2-forward 5′-CAT ACT CGA GCG GAC AGA GAC GTG AAG
CAC-3′ and RhoGDI2-reverse 3′-CAC TGG ATC CGA GTG ACA GGG TGG GAA
AAG-5′ (the restriction sites of XhoI and BamHI are
underlined) and inserted into pIRES2-EGFP (Clontech Laboratories,
Mountain View, CA, USA) at XhoI and BamHI sites.
MKN-45 cells were transfected with pIRES2-EGFP-RhoGDI2 or
pIRES2-EGFP using Lipofectamine 2000 reagent (Invitrogen) according
to the manufacturer’s instructions. Stable plasmid-transfected
clones were selected using 800 μg/ml G418 (Invitrogen) for 2
weeks, isolated colonies were picked up with tips and the cells
were further cultured in the presence of 400 μg/ml G418.
MKN-45 cells transfected with pIRES2-EGFP-RhoGDI2 were named
MKN-45/RhoGDI2 cells. MKN-45 cells transfected with pIRES2-EGFP
were named MKN-45/GFP cells.
In vitro cytotoxicity assay
The MTT assay was used to determine the relative
sensitivity of cell lines to 5-FU (Xudonghaipu Pharmaceutical Co.,
Shanghai, China), as described previously (11). For cell lines, cells plated in
96-well microplates were cultured with growth medium or treated
with serial dilutions of 5-FU for 72 h. Viable cells were measured
with MTT (Sigma, St. Louis, MO, USA) and the results were expressed
relative to the absorbance of cells grown in the absence of the
drug. IC50 values were calculated by nonlinear regression analysis
from triplicate independent experiments.
For patient samples, tissues were minced by scissors
into RPMI-1640 medium. The tumor cells were then incubated at 37°C
for 30 min in an enzyme cocktail containing 0.02% deoxyribonuclease
I (Sigma), 0.05% pronase (Calbiochem, Dormstadt, Germany) and 0.02%
collagenase. The tumor cell suspension (5×105 cells/ml)
was then strained through a 150 μm stainless steel mesh. The
cells were centrifuged at 1,000 rpm for 5 min and, following
rinsing twice, the gastric carcinoma cells were verified by 0.25%
trypan blue dye exclusion (Sigma). The cell number was adjusted to
1×105 cells/ml and a 180 μl aliquot of the tumor
cell suspension was plated into each well of a 96-well cell culture
plate (Nunc Inc., Rochester, NY, USA), followed by the addition of
20 μl of each dose of 5-FU (1 mg/ml). The cells were then
incubated at 37°C for 72 h in a 5% CO2 incubator. The
cells were washed with phosphate-buffered saline and subsequently
25 μl of MTT (2 mg/ml; Sigma) was added and measured as
described previously (11). The
tumor inhibition rate (IR) was calculated from the following
equation: IR (%) = (1−T/C) × 100, where T=OD540 of the
treated cells and C=OD540 of the control cells.
Western blot analysis
Western blot analysis was performed as previously
described (12). Briefly, cell
lysates were separated by SDS-PAGE and transferred to a PVDF
membrane. The blot was then probed with anti-RhoGDI2 (LabVision,
Fremont, CA, USA) with a dilution of 1:1,000, followed by an
incubation with a horseradish peroxidase-conjugated secondary
antibody. The signal was detected using enhanced chemiluminescence
(Millipore, Billerica, MA, USA). The expression level was
quantified using the Image J programme (NIH).
Immunohistochemical staining
Immunohistochemical staining was performed as
described previously using a DAKO EnVision+ System HRP (13). RhoGDI2 polyclonal antibody from
LabVision was applied at a 1:2,000 dilution overnight at 4°C.
Purified rabbit-IgG was used as an isotype control. The stained
sections were reviewed by two independent observers (Z.Z. and
X.Y.H.) who had no prior knowledge of the clinical pathological
data of the patients. A scoring method was used as reported
previously based on the evidence that the specimens clearly
demonstrated varying degrees of staining intensity and percentage
of cell staining (14). Briefly,
strong-intensity staining was scored as 3, moderate as 2, weak as 1
and negative as 0. For each intensity score, the percentage of
cells with that score was estimated visually. A combined weighted
score consisting of the sum of the percentage of cells staining at
each intensity level was calculated for each sample. The
immunolabelling was categorised as negative (score>30) or
positive (score ≤30) for all the tissues.
Cell cycle analysis
Cells were plated in triplicate in 6-well plates.
After 24 h, cells adherent to plates were exposed to 0.5 or 5
μg/ml 5-FU for 72 h. The percentage of viable cells was
determined by counting trypan blue exclusive cells by a
hemocytometer. For cycle analysis, cells were harvested, fixed in
70% ice-cold ethanol and incubated with propidium iodide (20
μg/ml) and RNase A (100 μg/ml) for 1 h. Propidium
iodide-stained DNA content was then assessed using a FACSCalibur
flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA) and the
percentage of cells in each phase of the cell cycle was analysed
using Flowjo 8.0 (Tree Star, Ashland, OR, USA).
Statistical analysis
Statistical analyses of data were performed using
the Student’s t-test or one-way analysis of variance, depending on
the number of groups in comparison. Data that failed the test for
normal distribution or homogeneous variance were analysed using the
Mann-Whitney U or the Kruskal-Wallis tests. Correlations between
RhoGDI2 and 5-FU sensitivity were calculated using the Pearson
coefficient test. The statistical software SPSS version 14.0 (SPSS,
Inc., Chicago, IL, USA) was used for analysis. P<0.05 was
considered to indicate a statistically significant result.
Results
Negative correlation between RhoGDI2
expression and 5-FU sensitivity in human gastric cancer
Western blot analysis was used to determine the
level of RhoGDI2 in seven human gastric cancer cell lines. As shown
in Fig. 1A, RhoGDI2 was expressed
in all seven gastric cancer cell lines. The poorly differentiated
cell line KATO-III derived from signet ring carcinoma had the
highest level of RhoGDI2, which was 1.87-fold higher than that of
the well-differentiated cell line, SGC7901. The sensitivity to 5-FU
of each gastric cancer cell line was measured by the MTT assay and
the IC50 values are indicated in Fig.
1B. Of note, the IC50 of each cell line positively correlates
with the RhoGDI2 level (P=0.006, Fig.
1C). These results indicate that RhoGDI2 is associated with
resistance to 5-FU in gastric cancer cells.
Subsequently, the 5-FU sensitivity of tumor cells
isolated from gastric cancer tissues was tested and compared with
the level of RhoGDI2 expression. Since the RhoGDI2 was markedly
expressed in stroma, IHC was used to measure RhoGDI2 expression
specifically in cancer cells. Consistent with the results obtained
using the cell lines, the 5-FU sensitivity of patient cancer cells
expressing RhoGDI2 was 30% less than that of the negative controls
(P<0.05, Mann-Whitney U test), as shown in Fig. 1D.
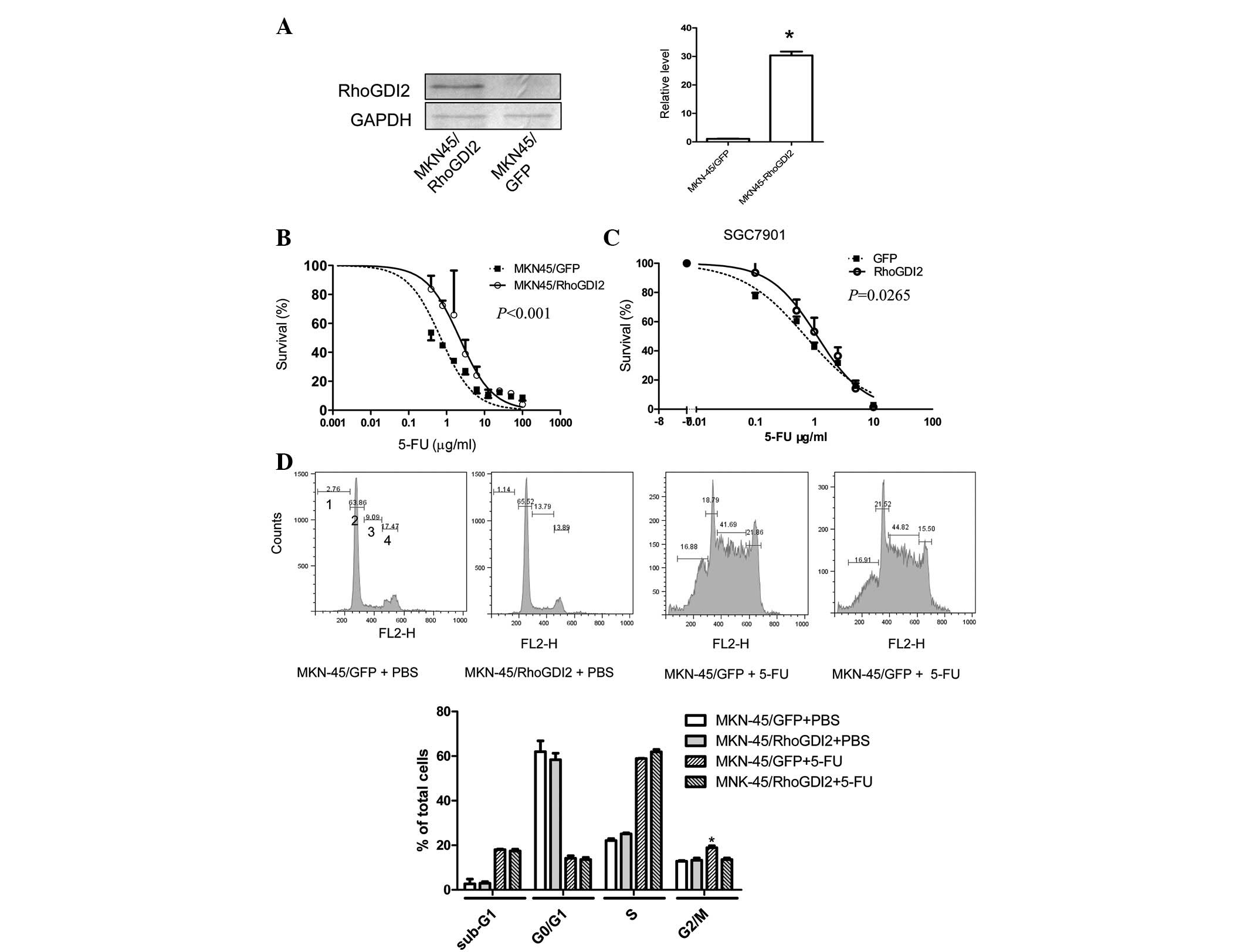
Ectopic expression of RhoGDI2 in gastric
cancer cells induces resistance to 5-FU and reverts 5-FU induced
G2/M phase arrest
To determine whether increased levels of RhoGDI2
induced 5-FU resistance, a gastric cancer cell line
(MKN-45/RhoGDI2) was established that had been stably transfected
with a RhoGDI2 expression vector. The RhoGDI2 level was markedly
increased in MKN-45/RhoGDI2, compared with the control cell line
MKN-45/GFP (Fig. 2A), while the
sensitivity to 5-FU was decreased (Fig.
2B, P<0.0001). To verify that this effect was not a cell
type-specific phenomenon, another gastric cancer cell line,
SGC7901, was transiently transfected with the RhoGDI2 coding
vector. Following 24 h of transfection, cells were replated for the
5-FU sensitivity test. The IC50 of 5-FU in cells transfected with
RhoGDI2 vector was increased by 60% compared with cells transfected
with a GFP vector (P=0.0265, Fig.
2C).
To determine whether RhoGDI2 influences the cell
cycle following 5-FU treatment, PI staining followed by flow
cytometry was used. It is reported that the dual antitumor effects
of 5-FU on the cell cycle is dependent on 5-FU concentration:
exposure to low dose 5-FU resulted in G2-phase arrest and mitotic
catastrophe, whereas a high dose of 5-FU induced G1-phase arrest
and apoptosis (15). Ectopic
expression of RhoGDI2 alone did not influence the cell cycle
distribution (Fig. 2D). Following
0.5 μg/ml (low dose) 5-FU treatment for 72 h, the percentage
of MKN-45/GFP cells in G1 decreased, whereas the proportion of
cells in S and G2 phase increased. Compared with MKN-45/GFP cells
treated with 5-FU, there were fewer MKN-45/RhoGDI2 cells in G2/M
(P<0.05) following 5-FU treatment, but no difference was
observed in PBS-treated MKN-45/RhoGDI2 cells. In addition, the
sub-G1 proportion, known as apoptotic cells, remained unchanged
between MKN-45/GFP and MKN-45/RhoGDI2 cells following exposure to
5-FU. Exposure to a high dose of 5-FU (5 μg/ml) resulted in
G1/arrest in MKN-45/GFP and MKN-45/RhoGDI2 and there was no
difference in sub-G1, G1, S, G2/M phase proportion between them
(data not shown). These results suggest that RhoGDI2 reverts
5-FU-induced G2/M phase arrest without influencing apoptosis and G1
phase arrest.
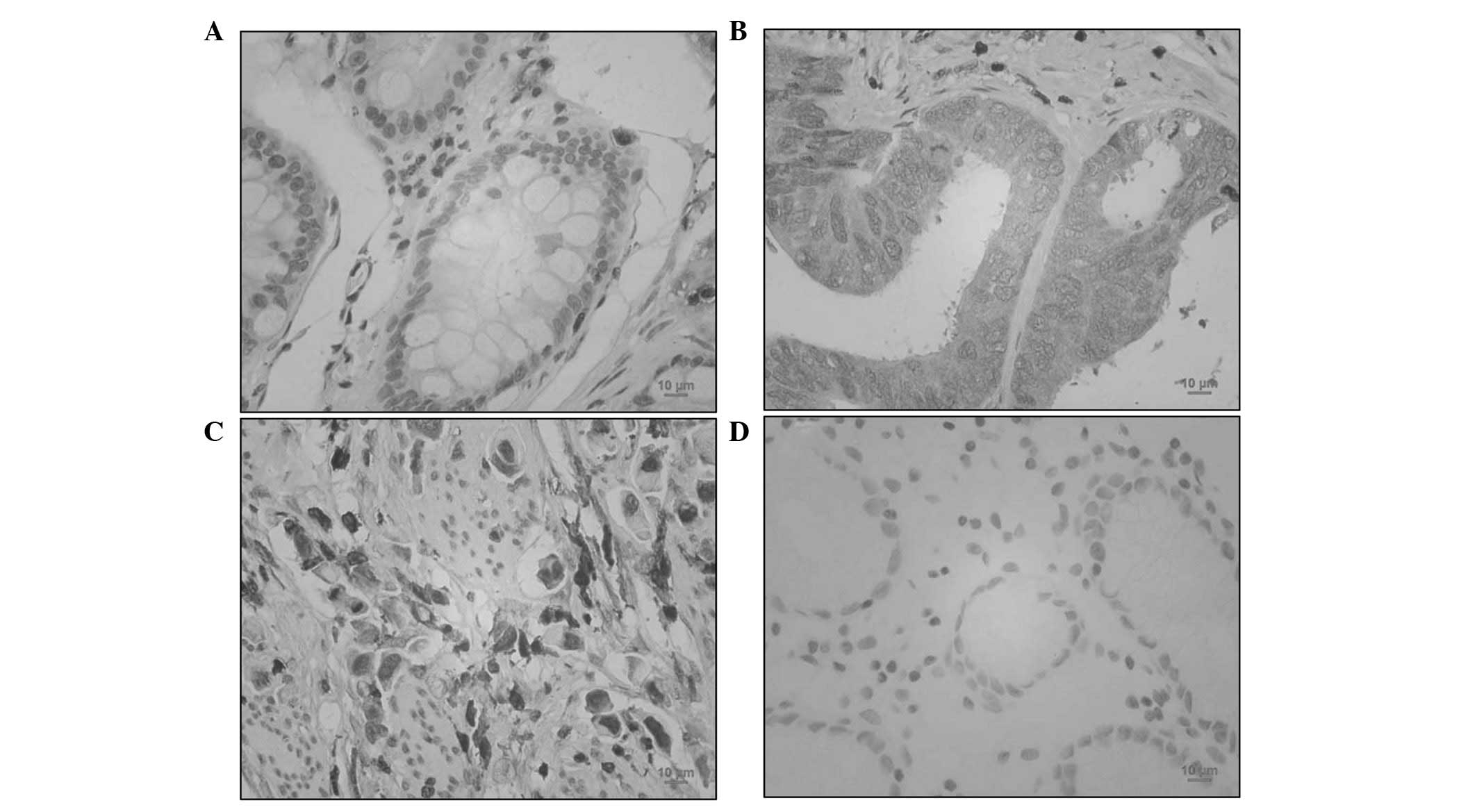
RhoGDI2 was upregulated in human gastric
cancer
To determine the expression of RhoGDI2 in patients
with gastric cancer, RhoGDI2 was analysed by IHC. As shown in
Fig. 3, benign gastric epithelial
cells were negatively or weakly (3 out of 88 cases) stained with
RhoGDI2 antibody. By contrast, 53.4% gastric cancer specimens (47
out of 88 cases) were positively stained with anti-RhoGDI2
(P<0.01 vs. normal gastric mucosa). Thus, RhoGDI2 was
overexpressed in gastric cancer cells. In intestinal-type gastric
cancer cells, RhoGDI2 was mainly expressed in the plasma (Fig. 3B). However, diffuse-type gastric
cancer cells were also markedly stained with anti-RhoGDI2 in the
nucleus (Fig. 3C).
Discussion
In our previous study using 2D electrophoresis-mass
spectrometry, it was revealed that RhoGDI2 is a contributor to 5-FU
resistance in colon cancer (11).
The current study demonstrated that RhoGDI2 also confers resistance
to 5-FU in gastric cancer cells. Firstly, the endogenous level of
RhoGDI2 was correlated with 5-FU resistance. Secondly, the ectopic
expression of RhoGDI2 induced resistance to 5-FU. In addition,
gastric cancer cells isolated from patients with higher levels of
RhoGDI2 were more resistant to 5-FU. Other groups have shown that
RhoGDI2 was overexpressed in chemotherapy-resistant fibrosarcoma
cells and paclitaxel-resistant ovarian cancers, respectively
(16,17). However, Lee et al(18) recently revealed contradictory
results suggesting that the chemoresistance in ovarian serous
carcinomas is associated with the downregulation of RhoGDI2
expression. Hee et al(19)
reported that RhoGDI2 confers resistance against cisplatin-induced
apoptosis in gastric cancer cells. The results of the current study
lead to the conclusion that high levels of RhoGDI2 expression are
associated with chemotherapy resistance in certain types of
cancers, including gastroenterological cancer.
The role of RhoGDI2 in apoptosis is contradictory.
RhoGDI2 is proteolysed by caspase-3 during Fas-induced apoptosis
(20) or in Burkitt-like lymphoma
cells with taxol or epirubicin (21). However, the function of RhoGDI2
cleavage has only recently been elucidated. Based on the
observation that the expression of a non-cleavable RhoGDI2 D19A
mutant did not protect CHO cells against apoptosis induction, it
was concluded that RhoGDI2 cleavage does not contribute to cell
death (22). By contrast, another
study suggested that the cleavage of RhoGDI2 by caspase-3 is not a
functionally irrelevant effect of caspase activation during
apoptosis but rather expedites the progression of the apoptotic
process (23). Unlike bladder
cancer and lymphoma cells, RhoGDI2 was reported as an oncogene in
breast, colon and gastric cancer cells (9,10,24). A
Korean group reported that RhoGDI2 confers resistance to
cisplatin-induced apoptosis in gastric cancer cells by upregulating
Bcl-2 expression and activating PLCγ (19,25).
Our results also demonstrated that RhoGDI2 is involved in the
resistance to 5-FU in gastric cancer cells. However, these data
suggest that the resistance was not due to preventing apoptosis,
but reverting G2/M phase arrest by RhoGDI2. RhoGDI2 may exert its
chemo-resistant function through distinct mechanisms in response to
various drugs, depending on the cell type.
It has been reported that there are dual antitumor
effects of 5-FU on the cell cycle depending on 5-FU concentration:
exposure to low dose 5-FU resulted in G2-phase arrest and mitotic
catastrophe, whereas a high dose of 5-FU induced G1-phase arrest
and apoptosis (15). In MKN-45
cells, similar dual effects were observed at 0.5 and 5 μg/ml
5-FU, respectively. However, RhoGDI2 only affected the low dose
5-FU-induced G2/M arrest, but not G1 arrest induced by high dose
5-FU. This is consistent with the results shown in Fig. 2B, which demonstrate that the
resistance to 5-FU conferred by RhoGDI2 expression is mainly
present at low doses of 5-FU and disappeared at higher doses of
5-FU.
In conclusion, it is suggested that RhoGDI2 is
aberrantly overexpressed and is also a direct contributor to the
resistance to chemotherapeutic agents, including 5-FU, in gastric
cancer cells. The potential mechanism involves inhibiting
5-FU-induced G2/M phase arrest. In this context, RhoGDI2 may be
used as a molecular target for the sensitisation of
gastrointestinal cancer cells during chemotherapy and as a
predictor of chemotherapy treatment outcomes.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Heasman SJ and Ridley AJ: Mammalian Rho
GTPases: new insights into their functions from in vivo studies.
Nat Rev Mol Cell Biol. 9:690–701. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jaffe AB and Hall A: Rho GTPases:
biochemistry and biology. Annu Rev Cell Dev Biol. 21:247–269. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dovas A and Couchman JR: RhoGDI: multiple
functions in the regulation of Rho family GTPase activities.
Biochem J. 390:1–9. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang Y, Rivera Rosado LA, Moon SY and
Zhang B: Silencing of D4-GDI inhibits growth and invasive behavior
in MDA-MB-231 cells by activation of Rac-dependent p38 and JNK
signaling. J Biol Chem. 284:12956–12965. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moissoglu K, McRoberts KS, Meier JA,
Theodorescu D and Schwartz MA: Rho GDP dissociation inhibitor 2
suppresses metastasis via unconventional regulation of RhoGTPases.
Cancer Res. 69:2838–2844. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gildea JJ, Seraj MJ, Oxford G, et al:
RhoGDI2 is an invasion and metastasis suppressor gene in human
cancer. Cancer Res. 62:6418–6423. 2002.PubMed/NCBI
|
|
8
|
Theodorescu D, Sapinoso LM, Conaway MR,
Oxford G, Hampton GM and Frierson HF Jr: Reduced expression of
metastasis suppressor RhoGDI2 is associated with decreased survival
for patients with bladder cancer. Clin Cancer Res. 10:3800–3806.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Y and Zhang B: D4-GDI, a Rho GTPase
regulator, promotes breast cancer cell invasiveness. Cancer Res.
66:5592–5598. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cho HJ, Baek KE, Park SM, et al: RhoGDI2
expression is associated with tumor growth and malignant
progression of gastric cancer. Clin Cancer Res. 15:2612–2619. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zheng Z, Li J, He X, et al: Involvement of
RhoGDI2 in the resistance of colon cancer cells to 5-fluorouracil.
Hepatogastroenterology. 57:1106–1112. 2010.PubMed/NCBI
|
|
12
|
Wang YW, Qu Y, Li JF, et al: In vitro and
in vivo evidence of metallopanstimulin-1 in gastric cancer
progression and tumorigenicity. Clin Cancer Res. 12:4965–4973.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yuen HF, Chan YP, Chan KK, et al: Id-1 and
Id-2 are markers for metastasis and prognosis in oesophageal
squamous cell carcinoma. Br J Cancer. 97:1409–1415. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ben QW, Wang JC, Liu J, et al: Positive
expression of L1-CAM is associated with perineural invasion and
poor outcome in pancreatic ductal adenocarcinoma. Ann Surg Oncol.
17:2213–2221. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yoshikawa R, Kusunoki M, Yanagi H, et al:
Dual antitumor effects of 5-fluorouracil on the cell cycle in
colorectal carcinoma cells: a novel target mechanism concept for
pharmacokinetic modulating chemotherapy. Cancer Res. 61:1029–1037.
2001.
|
|
16
|
Sinha P, Hütter G, Köttgen E, Dietel M,
Schadendorf D and Lage H: Search for novel proteins involved in the
development of chemoresistance in colorectal cancer and
fibrosarcoma cells in vitro using two-dimensional electrophoresis,
mass spectrometry and microsequencing. Electrophoresis.
20:2961–2969. 1999. View Article : Google Scholar
|
|
17
|
Goto T, Takano M, Sakamoto M, et al: Gene
expression profiles with cDNA microarray reveal RhoGDI as a
predictive marker for paclitaxel resistance in ovarian cancers.
Oncol Rep. 15:1265–1271. 2006.PubMed/NCBI
|
|
18
|
Lee DH, Chung K, Song JA, et al: Proteomic
identification of paclitaxel-resistance associated hnRNP A2 and GDI
2 proteins in human ovarian cancer cells. J Proteome Res.
9:5668–5676. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cho HJ, Baek KE, Park SM, et al: RhoGDI2
confers gastric cancer cells resistance against cisplatin-induced
apoptosis by upregulation of Bcl-2 expression. Cancer Lett.
311:48–56. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Na S, Chuang TH, Cunningham A, et al:
D4-GDI, a substrate of CPP32, is proteolyzed during Fas-induced
apoptosis. J Biol Chem. 271:11209–11213. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Essmann F, Wieder T, Otto A, Müller EC,
Dörken B and Daniel PT: GDP dissociation inhibitor D4-GDI (Rho-GDI
2), but not the homologous rho-GDI 1, is cleaved by caspase-3
during drug-induced apoptosis. Biochem J. 346:777–783. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Krieser RJ and Eastman A: Cleavage and
nuclear translocation of the caspase 3 substrate Rho
GDP-dissociation inhibitor, D4-GDI, during apoptosis. Cell Death
Differ. 6:412–419. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Choi MR, Groot M and Drexler HC:
Functional implications of caspase-mediated RhoGDI2 processing
during apoptosis of HL60 and K562 leukemia cells. Apoptosis.
12:2025–2035. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li X, Wang J, Zhang X, Zeng Y, Liang L and
Ding Y: Overexpression of RhoGDI2 correlates with tumor progression
and poor prognosis in colorectal carcinoma. Ann Surg Oncol.
19:145–153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cho HJ, Baek KE, Nam IK, et al: PLCgamma
is required for RhoGDI2-mediated cisplatin resistance in gastric
cancer. Biochem Biophys Res Commun. 414:575–580. 2011. View Article : Google Scholar : PubMed/NCBI
|