Introduction
Head and neck cancer (HNC) is the 6th most common
type of cancer worldwide (1), and
comprises cancers of the aerodigestive tract, including the lips,
oral cavity, nasal cavity, paranasal sinuses, pharynx, larynx,
oropharynx, hypopharynx, salivary glands and local lymph nodes
(2). Approximately 90% of all HNCs
are squamous cell carcinomas (HNSCCs), which arise from the mucosal
lining in these regions (2). The
5-year survival rate for HNSCC is among the lowest of the most
common types of cancer; having remained at 64–67% for the last ten
years (3). Approximately 80–90% of
cases of HNSCC are attributed to prolonged tobacco and/or alcohol
use (2). However, only a small
fraction of those who consume tobacco or alcohol develop this
disease, suggesting that genetic factors are also important in its
pathogenesis (4–6). Fundamental to the genetic basis of all
cancers is the overexpression of oncogenes and/or silencing of
tumor suppressor genes. Aberrations, such as deletions or mutations
of these genes may lead to uncontrolled cell division and cancer
development.
Ras-associated binding (Rab)-GTPases are the largest
subfamily of small GTPases, and are involved in the regulation of
intracellular vesicle transport and protein trafficking (7–12). Rab
proteins associate with distinct membrane compartments within cells
and determine the specificity of vesicle trafficking pathways; the
nidi for the assembly of multiprotein complexes (13). More than 60 mammalian Rab proteins
have been identified to date, all of which exert their specific
function by cycling between a GTP-bound active conformation and,
following GTP hydrolysis, a GDP-bound inactive state (14).
Rab25 was identified in 1993 as a member of the Rab
family. Rab25 is also known as Rab11c, due to its sequence homology
with the Rab11a and Rab11b proteins (15). However, it is expressed only in
epithelial cells, in contrast to the ubiquitously expressed Rab11a
and Rab11b (15,16). Functional studies have demonstrated
that the Rab11 subfamily is involved in transcytosis, endocytic
sorting and transport across polarized epithelial cells, as well as
in apical vesicle recycling (17,18).
He et al (16)
reported that the expression of 9 human GTPases is upregulated in
liver cancer and that 6 of these were Rab proteins. This was the
first data that demonstrated an association between Rab25 and
epithelial cancer. Subsequently, overexpression of Rab25 has been
shown to be associated with numerous types of human cancer
(19–21). However, recent studies have identified
two opposing roles for Rab25 in tumor progression. Several groups
have suggested that Rab25 exerts a tumor promoting function, and
have proposed that its overexpression is associated with an
increase in tumor aggression and invasiveness (22–25).
Overexpression of Rab25 has been reported in ovarian (22–24),
prostate (26), invasive breast
(27), colon (15) and liver cancer (16), in addition to transitional cell
carcinoma of the bladder (27,28) and
Wilms' tumor (29), which indicates
that Rab25 acts as an oncogene in these situations. By contrast, a
documented association between loss of Rab25 expression and
increased tumor development, indicates that Rab25 may function as a
tumor suppressor in a number of types of cancer. Loss of Rab25
expression has been detected in certain breast and colon cancer
tissues, and in a novel breast cancer cell line, which was
generated from human mammary epithelial cells (30,31). More
recently, Zhang et al (25)
reported that Rab25 is upregulated in the majority of bladder
tumors and that this upregulation is associated with tumor invasion
and metastasis. The in vitro studies by that group have also
demonstrated that Rab25 exerts its oncogenic effect via the
Akt/glycogen synthase kinase-3β (GSK-3β)/Snail pathway. These
conflicting results indicate that Rab25 may exert different effects
in different types of tumor. To the best of our knowledge, only one
study has investigated the role of Rab25 in HNC cell lines
(32), the results of which suggested
that Rab25 is involved in tumor migration and metastasis in HNSCC.
The aim of the present study was to investigate the function of
Rab25 in human HNSCC tissue samples and to examine its association
with the Akt pathway. Therefore, the expression of Rab25 in HNSCC
tumors was measured using reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) and western blotting.
Materials and methods
Tissue samples
Paired normal and tumor samples were obtained from
76 patients with HNSCC, who underwent surgery at the Department of
Otorhinolaryngology (Cerrahpasa Medical Faculty, Istanbul, Turkey)
following receipt of written informed consent. This study was
approved by the Cerrahpasa Medical Faculty Ethics Committee
(Approval no. 83045809/11295). Clinicopathological characteristics,
including patient age, gender, TNM staging, histological grade,
invasion status and tumor location, are presented in Table I.
 | Table I.Association of Rab25 protein and mRNA
expression with clinicopathological characteristics. |
Table I.
Association of Rab25 protein and mRNA
expression with clinicopathological characteristics.
|
|
| Rab25 protein, n
(%) |
| Rab25 mRNA, n
(%) |
|
|---|
|
|
|
|
|
|
|
|---|
| Characteristic | Patients, n
(%) | Decreased | Increased | P-value | Decreased | No change | Increased | P-value |
|---|
| Location |
|
|
|
| 0.492 |
|
|
| 0.976 |
|
Larynx | 59 (77.6) | 23 (52.3) | 21 (47.7) |
| 39 (66.1) | 3 (5.1) | 17 (28.8) |
|
| Oral
cavity | 9 (11.8) | 4 (57.1) | 3 (42.9) |
| 6 (66.7) | – | 3 (33.3) |
|
|
Parotitis | 5 (6.6) | 3 (75.0) | 1 (25.0) |
| 4 (80.0) | – | 1 (20.0) |
|
|
Paranasal sinus | 3 (3.9) | 2 (100.0) | 0 (0.0) |
| 2 (66.7) | – | 1 (33.3) |
|
| Stage |
|
|
| 0.547 |
|
|
| 0.411 |
| 1 | 2 (2.6) | – | – |
| 1 (50.0) | – | 1 (50.0) |
|
| 2 | 27 (27.6) | 12 (66.7) | 6 (33.3) |
| 14 (51.9) | 2 (7.4) | 4 (40.7) |
|
| 3 | 21 (35.5) | 9 (50.0) | 9 (50.0) |
| 17 (63.0) | – | 10 (37.0) |
|
| 4 | 26 (34.2) | 11 (52.4) | 10 (47.6) |
| 19 (73.1) | 1 (3.8) | 6 (23.1) |
|
| Histology |
|
|
| 0.384 |
|
|
| 0.692 |
|
SCC | 67 (88.2) | 27 (54.0) | 23 (46.0) |
| 44 (65.7) | 3 (4.5) | 20 (29.8) |
|
|
Other | 9 (11.8) | 5 (71.4) | 2 (28.6) |
| 7 (57.1) | – | 2 (42.9) |
|
| Local invasion |
|
|
| 0.126 |
|
|
| 0.532 |
|
Negative | 18 (23.7) | 7 (77.8) | 2 (22.2) |
| 11 (61.1) | 1 (5.6) | 6 (33.3) |
|
|
Positive | 55 (72.4) | 22 (50.0) | 22 (50.0) |
| 40 (72.7) | 1 (1.8) | 14 (25.5) |
|
| Age, years |
|
|
| 0.080 |
|
|
| 0.850 |
|
≥60 | 36 (47.4) | 13 (44.8) | 16 (55.2) |
| 25 (69.4) | 1 (2.8) | 10 (27.8) |
|
|
<60 | 40 (52.6) | 19 (67.9) | 9 (32.1) |
| 26 (65.0) | 2 (5.0) | 12 (30.0) |
|
| Gender |
|
|
| 0.450 |
|
|
| 0.692 |
|
Male | 67 (88.2) | 29 (58.0) | 21 (42.0) |
| 44 (65.7) | 3 (4.5) | 20 (29.8) |
|
|
Female | 9 (11.8) | 3 (42.9) | 4 (57.1) |
| 7 (77.8) |
| 2 (22.2) |
|
cDNA synthesis and RT-qPCR
Total RNA was extracted from tumor and adjacent
non-tumor samples using the PureLink™ RNA Mini kit (Ambion Life
Technologies, Carlsbad, CA, USA). Single-stranded cDNA was
synthesized using the Revert-Aid first strand cDNA synthesis kit
(Fermantas, Vilnius, Lithuania) and 300 ng of total RNA in a
reaction volume of 20 µl. Glucose-6-phosphate dehydrogenase (G6PD)
was used as a reference gene for normalization. The resultant cDNA
(2 µl) was used for the Real-Time assay, which was performed in 20
µl reaction volume, containing 1X LightCycler 480 Probes Master Mix
(Roche Diagnostics, Mannheim, Germany), Rab25 (Intergrated DNA
Technologies, Inc., Toronto, ON, Canada) and G6PD gene-specific
primers (Roche Diagnostics), and Universal Probe Library hydrolysis
probes (Roche Diagnostics) labeled with 6-Carboxyflourescein or VIC
at the 5′-end, and with Non-flourescent Dark Quencer dye or HEX at
the 3′-end, for Rab25 and G6PD, respectively. The following primer
sequences were used: Forward, 5′-GCTGCTGTCAAGGCTCAGAT-3′ and
reverse, 5′-CCCACTGCACCACGATAGTA-3′ for Rab25; forward,
5′-GAGCCAGATGCACTTCGTG-3′ and reverse, 5′-GGGCTTCTCCAGCTCAATC-3′
for G6PD. The target and reference genes were co-amplified in the
same reaction. Amplification was performed at 95°C for 20 sec, 60°C
for 15 sec and 72°C for 20 sec for 45 cycles, with an initial
denaturation at 95°C for 10 min, using the Light Cycler 480 II
platform (Roche Diagnostics). Changes in the expression levels were
calculated using the 2−ΔΔCt method (33).
Western blotting
Tissue samples were rinsed with ice-cold
phosphate-buffered saline and homogenized in a protease inhibitor
(PMSF), containing protein extraction buffer (1X lysis buffer, 1X
TBS, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% SDS, 0.004%
sodium azide), in order to obtain whole cell lysate. Equal
quantities of protein (30 µg) from each sample were loaded onto 12%
SDS-PAGE gels and electrophoresed at 165 V for 50 min at room
temperature. Resolved protein samples were transferred to
nitrocellulose membranes (Invitrogen Life Technologies, Carlsbad,
CA, USA) using the iBlot® Dry Blotting system (Invitrogen Life
Technologies) for western blot analysis. Following transfer,
immunoblots were blocked overnight in 5% non-fat dry milk (w/v) in
Tris-buffered saline (TBS), containing 20 mM Tris-HCl (pH 7.6), 150
mM NaCl and 0.1% Tween-20, and probed with the mouse anti-Rab25
monoclonal antibody, clone 12C3 (cat no. 05–1574; EMD Millipore,
Billerica, MA, USA), which was diluted to 1/1,000 with 1% non-fat
dry milk in TBS with Tween-20 (TBST; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA). In order to measure Akt1 or phosphorylated
Akt1 protein levels, the blots were incubated with primary
monoclonal mouse Akt1 (B1; cat. no. sc-5298; 1/2,000 dilution;
Santa Cruz Biotechnology, Inc.) and mouse p-Akt1 (5.Ser 473; cat
no. sc-293125; 1/2,000 dilution; Santa Cruz Biotechnology, Inc.)
antibodies for 90 min at room temperature, respectively. The blots
were washed with TBST three times and the protein content was
visualized using HRP-conjugated goat anti-mouse secondary
antibodies for Akt1 and p-Akt-1 (cat. no. sc-2005; 1/1,000
dilution; Santa Cruz Biotechnology, Inc.) and Rab25 (cat. no.
AP181P; 1/1,000 dilution; EMD Millipore) for 60 min at room
temperature. The protein bands were detected by the Western
Lightening Chemiluminescence reagent (Invitrogen Life
Technologies). The intensity of luminescence was quantified using
the ImageJ 1.47 bundle with the Java analyzing program (http://rsb.info.nih.gov/ij/download.html).
Statistical analysis
The χ2 test was performed to analyze the
differences in expression levels between tumor and normal tissues.
The correlation between expression levels and clinicopathological
parameters was analyzed using the Spearman's Rho test. Statistical
analysis was performed using IBM SPSS software (version 20.0.0; IBM
SPSS, Armonk, NY, USA). For all statistical tests, P<0.05 was
considered to indicate a statistically significant difference.
Results
Rab25 mRNA expression is decreased in
HNSCC, although its expression is not correlated with tumor
invasion
Rab25 mRNA expression was analyzed using RT-qPCR in
samples from patients with HNSCC and compared with matched adjacent
non-malignant tissues. Complete loss of Rab25 mRNA expression was
observed in 9 out of 76 (11.8%) tumor samples. When the mean ΔCt
level of the remaining 67 samples was calculated, a 3.44-fold
decrease in Rab25 expression was observed in the tumor samples
compared with that in the non-malignant tissue samples, following
normalization to G6PD (Table II).
Decreased Rab25 expression was observed in 69.1% of cases. The
majority of patients were male (88.2%), and had laryngeal cancer
(77.6%) and SCC (88.2%). Fifty-five of the patients had evidence of
vascular (26%), lymphatic (57.5%), perineural (39.7%) and/or
cartilage invasion (43.7%). However, when the association between
Rab25 expression and invasion was analyzed, no significant
correlation was detected between Rab25 expression and tumor
invasion (P=0.529). Similarly, no significant correlation was
identified between Rab25 expression and other clinicopathological
variables (Table I). None of the
patients had evidence of metastasis to any other site.
| Table II.Median expression of Rab25 mRNA
expression in tumor and normal tissues. |
Table II.
Median expression of Rab25 mRNA
expression in tumor and normal tissues.
| Tissue | Rab25 Ct
(median) | G6PD Ct
(median) | ΔCt | ΔΔCt |
2−ΔΔCt |
|---|
| HNSCC | 29.45 | 26.96 | 2.50 | 1.77 | 0.29 |
| Normal | 28.77 | 28.03 | 0.73 | 0 | 1 |
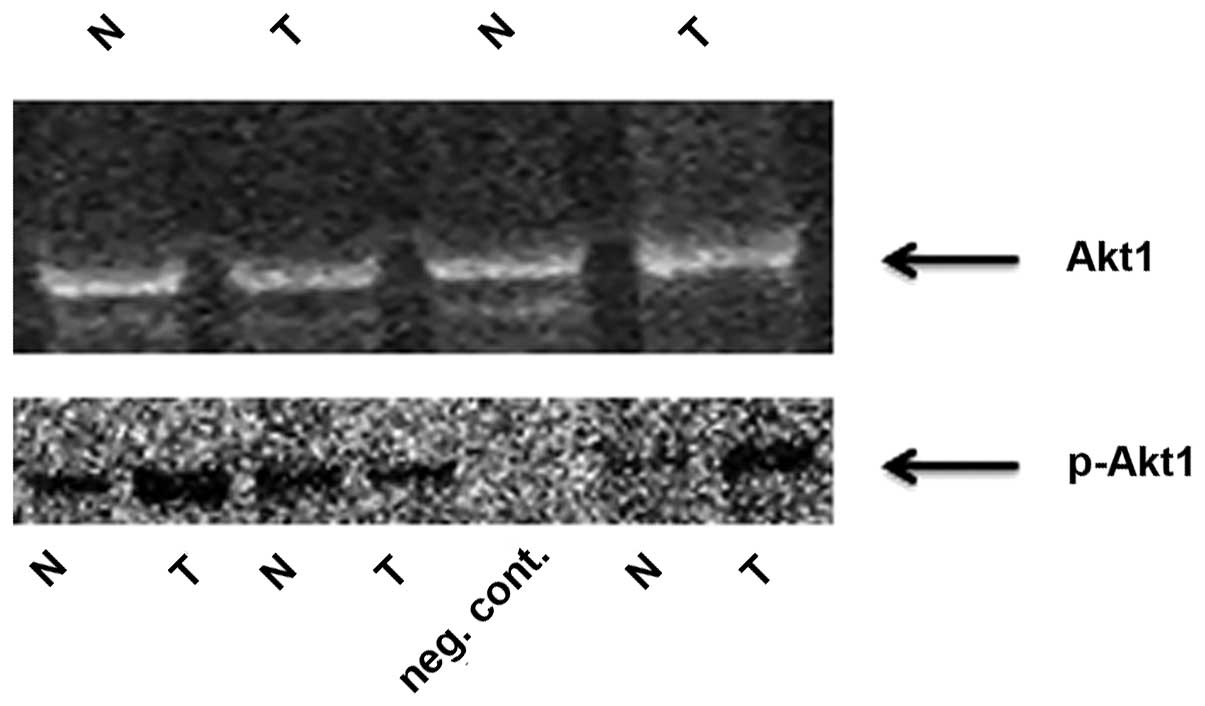
Rab25 protein is downregulated and
p-Akt1 is upregulated in HNSCC
The Rab25 protein was also downregulated in 56.1% of
tumor tissues and this downregulation at the protein level was
positively correlated with Rab25 mRNA expression. By contrast,
p-Akt1 expression was increased in tumor tissues, compared with
that in non-cancerous tissues (Fig.
1), and this upregulation in p-Akt1 levels was inversely
correlated with Rab25 protein expression (P=0.032).
Discussion
Malfunction of the vesicular traffic system is an
abnormal biological behavior that occurs in cancer cells. It has
been shown that the Ras superfamily proteins regulate various steps
of vesicular trafficking in eukaryotic cells (34). Rab25, a member of the Ras superfamily,
has been demonstrated to be involved in cell proliferation and in
protection of cells from apoptosis (22,23–35).
Initially, Rab25 was hypothesized to be associated with epithelial
cancer, following reports that Rab25 is one of 6 Rab proteins that
are upregulated in hepatocellular carcinoma (17). Subsequently, Rab25 was identified as a
driver of the 1q22 amplicon, which is frequently amplified in
ovarian cancer (22–35). In a separate study, Rab25 expression
was also shown to be correlated with histological grade and it was
defined as an androgen response gene in ovarian cancer (36). Increased expression of Rab25 was
observed in breast, ovarian and bladder cancer, and was associated
with aggressive tumor features (22,23,28). By
contrast, a tumor suppressor role for Rab25 has also been
demonstrated in colon cancer, triple-negative breast cancer and
esophageal SCC (ESCC) (31,37,38). These
results indicate that changes in the expression of Rab25 may depend
on the cell type and, therefore, the mechanism underlying the
effect of this protein on cancer progression may be tumor-type
specific. Although data on the involvement of the Rab25 in
proliferation, survival, apoptotic signaling, angiogenesis,
invasion and metastasis are available, to the best of our knowledge
there is no evidence of the precise molecular mechanisms underlying
its effects. One study has suggested that it affects
TIMP1-associated tumor growth and angiogenesis (39), while another reported that Rab25 is a
regulator of cellular bioenergetics in cancer (40). Caswell et al (41) hypothesized that Rab25 promotes α5β1
integrin-containing vesicle trafficking to the membrane of
pseudopodia tips, thereby facilitating cell invasion in ovarian
cancer. A further study demonstrated that overexpression of Rab25
in rat intestinal cells may lead to microtubule-dependent
transformation in vitro and tumor formation in vivo
(42). Multiple studies have
implicated Rab25 in invasion and cell migration. However, the
results have been contradictory, with certain studies demonstrating
an association between upregulation of Rab25 gene expression, and
metastasis and invasion, while other studies have produced the
opposite result. In order to evaluate the mechanisms underlying its
effects on invasion and metastasis, functional experiments have
been performed. Zhang et al (25) reported that Rab25 contributes to
metastasis through induction of epithelial mesenchymal transition
and activation of the Akt/GSK-3β/Snail pathway in bladder cancer.
Increased Akt phosphorylation has also been associated with
overexpression of Rab25 in aggressive breast and ovarian cancer
in vitro (22). Tong et
al (37) reported that Rab25
mediates tumorigenicity, metastasis and angiogenesis in ESCC, via
deregulation of FAK-Raf-MEK1/2-ERK signaling. By contrast,
Amornphimolthan et al (32)
demonstrated that Rab25 regulated cell migration and metastasis by
modulating the epidermal growth factor receptor-mediated pathway in
oral cancer. Until recently there were no reports on the expression
level of Rab25 in HNSCC. However, in 2013, Amornphimolthan et
al (32) reported that Rab25
functions as a tumor suppressor in HNSCC. In their study, the
expression of Rab25 in vitro and in vivo was
measured, in association with invasion and metastasis in HNSCC, and
it was concluded that downregulation of Rab25 is associated with
invasion and metastasis in this disease. However, to date that have
been no studies investigating the role of Rab25 in human HNSCC.
Therefore, in order to examine the effect of Rab25 in HNSCC the
present study was conducted using tissue samples from patients with
this disease. In accordance with the results reported by
Amornphimolthan et al (32),
the present study demonstrated that Rab25 was significantly
downregulated in 69.1% and 56.1% of the HNSCC tumor tissue samples
at the mRNA and protein levels, respectively. However, in contrast
to the results of the study by Amornphimolthan et al
(32), Rab25 downregulation was not
associated with tumor invasion in the current study (P=0.529). In
accordance with in vitro studies reported by Zhang et
al (25), downregulation of Rab25
was associated with increased Akt1 phosphorylation, which is an
indicator of Akt1 activation. The results of a study by Tang et
al (43) suggested a correlation
between loss of Rab25 expression, and H-ras or K-ras mutations in
breast cancer cell lines. In the present study group all of the
patients were K-ras-negative. Therefore, the current results do not
support the hypothesis proposed by Tang et al.
The majority of studies published on this subject
have been performed using cell lines and animal models to
investigate gene expression profiles. The regulation of gene
expression in vivo or under physiological conditions may be
distinct from that observed in vitro. To the best of our
knowledge, this is the first study to investigate Rab25 expression
levels in human HNSCC tissue samples. The results indicate that
Rab25 functions as a tumor suppressor in HNSCC. However, its
downregulation was not found to be associated with lymphatic,
vascular or cartilage invasion. All patients in the current study
population were metastasis-free, and in the absence of distant
metastasis, Rab25 was shown to be downregulated at the mRNA and
protein level in HNSCC tumor tissues. This downregulation was
associated with an increase in Akt1 phosphorylation. The current
results indicate that Rab25 primarily functions as a tumor
suppressor in HNSCC and exerts its effect by preventing Akt1
phosphorylation, which is a molecule that is known to activate the
mTOR pathway. In conclusion, Rab25 may have exert tumor suppressor
or oncogenic effects during tumor progression, depending on the
cell type; suppressing tumor progression by blocking Akt1
phosphorylation in particular types of tumor and promoting
progression by phosphorylating Akt1 in others. However, further
functional studies are required to elucidate the precise mechanisms
underlying these effects.
Acknowledgements
This study was supported by the Research Fund of
Istanbul University (grant nos. 13732 and 34503).
References
|
1
|
Warnakulasuriya S: Global epidemiology of
oral and oropharyngeal cancer. Oral Oncol. 45:309–316. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hashibe M, Brennan P, Chuang SC, et al:
Interaction between tobacco and alcohol use and the risk of head
and neck cancer: Pooled analysis in the International Head and Neck
Cancer Epidemiology Consortium. Cancer Epidemiol Biomarkers Prev.
18:541–550. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Howlader N, Noone AM, Krapcho M, et al:
SEER Cancer Statistics Review, 1975–2010. National Cancer
Institute; Bethesda, MD: http://seer.cancer.gov/csr/1975_2010/based on
November 2012 SEER data submission, posted to the SEER web site.
April. 2013
|
|
4
|
Báez A: Genetic and enviromental factors
in head and neck cancer genesis. J Environ Sci Health C Environ
Carcinog Ecotoxicol Rev. 26:174–200. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gleich LL and Salamone FN: Molecular
genetics of head and neck cancer. Cancer Control. 9:369–378.
2002.PubMed/NCBI
|
|
6
|
Demokan S, Suoglu Y, Demir D, et al:
Microsatellite instability and methylation of the DNA mismatch
repair genes in head and neck cancer. Ann Oncol. 17:995–999. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pereira-Leal JB and Seabra MC: Evolution
of the Rab family of small GTP-binding proteins. J Mol Biol.
313:889–901. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zerial M and McBride H: Rab proteins as
membrane organizers. Nat Rev Mol Cell Biol. 2:107–117. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Deneka M, Neeft M and van der Sluijs P:
Regulation of membrane transport by rab GTPases. Crit Rev Biochem
Mol Biol. 38:121–142. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pfeffer S and Aivazian D: Targeting Rab
GTPases to distinct membrane compartments. Nat Rev Mol Cell Biol.
5:886–896. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jordens I, Marsman M, Kuijl C and Neefjes
J: Rab proteins, connecting transport and vesicle fusion. Traffic.
6:1070–1077. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schwartz SL, Cao C, Pylypenko O, Rak A and
Wandinger-Ness A: Rab GTPases at a glance. J Cell Sci.
120:3905–3910. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Somsel Rodman J and Wandinger-Ness A: Rab
GTPases coordinate endocytosis. J Cell Sci 113 Pt. 2:183–92.
2000.
|
|
14
|
Vetter IR and Wittinghofer A: The guanine
nucleotide-binding switch in three dimensions. Science.
294:1299–1304. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Goldenring JR, Shen KR, Vaughan HD and
Modlin IM: Identification of a small GTP-binding protein, Rab25,
expressed in the gastrointestinal mucosa, kidney and lung. J Biol
Chem. 268:18419–18422. 1993.PubMed/NCBI
|
|
16
|
He H, Dai F, Yu L, et al: Identification
and characterization of nine novel human small GTPases showing
variable expressions in liver cancer tissues. Gene Expr.
10:231–242. 2002.PubMed/NCBI
|
|
17
|
Casanova JE, Wang X, Kumar R, et al:
Association of Rab25 and Rab11a with the apical recycling system of
polarized Madin-Darby canine kidney cells. Mol Biol Cell. 10:47–61.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang X, Kumar R, Navarre J, et al:
Regulation of vesicle trafficking in madin-darby canine kidney
cells by Rab11a and Rab25. J Biol Chem. 275:29138–29146. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Agarwal R, Jurisica I, Mills GB and Cheng
KW: The emerging role of the RAB25 small GTPase in cancer. Traffic.
10:1561–1568. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cheng KW, Lahad JP, Gray JW and Mills GB:
Emerging role of RAB GTPases in cancer and human disease. Cancer
Res. 65:2516–2519. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mitra S, Cheng KW and Mills GB: Rab25 in
cancer: A brief update. Biochem Soc Trans. 40:1404–1408. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cheng KW, Lahad JP, Kuo WL, et al: The
RAB25 small GTPase determines aggressiveness of ovarian and breast
cancers. Nat Med. 10:1251–1256. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fan Y, Xin XY, Chen BL and Ma X: Knockdown
of RAB25 expression by RNAi inhibits growth of human epithelial
ovarian cancer cells in vitro and in vivo. Pathology. 38:561–567.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Y, Tao X, Jia L, et al: Knockdown of
RAB25 promotes autophagy and inhibits cell growth in ovarian cancer
cells. Mol Med Rep. 6:1006–1012. 2012.PubMed/NCBI
|
|
25
|
Zhang J, Wei J, Lu J, et al:
Overexpression of Rab25 contributes to metastasis of bladder cancer
through induction of epithelial-mesenchymal transition and
activation of Akt/GSK-3β/Snail signaling. Carcinogenesis.
34:2401–2408. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Calvo A, Xiao N, Kang J, et al:
Alterations in gene expression profiles during prostate cancer
progression: Functional correlations to tumorigenicity and
down-regulation of selenoprotein-P in mouse and human tumors.
Cancer Res. 62:5325–5335. 2002.PubMed/NCBI
|
|
27
|
Wang W, Goswami S, Lapidus K, et al:
Identification and testing of a gene expression signature of
invasive carcinoma cells within primary mammary tumors. Cancer Res.
64:8585–8594. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mor O, Nativ O, Stein A, et al: Molecular
analysis of transitional cell carcinoma using cDNA microarray.
Oncogene. 22:7702–7710. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Natrajan R, Williams RD, Hing SN, et al:
Array CGH profiling of favourable histology Wilms tumours reveals
novel gains and losses associated with relapse. J Pathol.
210:49–58. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng JM, Ding M, Aribi A, et al: Loss of
RAB25 expression in breast cancer. Int J Cancer. 118:2957–2964.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Goldenring JR and Nam KT: Rab25 as a
tumour suppressor in colon carcinogenesis. Br J Cancer. 104:33–36.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Amornphimoltham P, Rechache K, Thompson J,
et al: Rab25 regulates invasion and metastasis in head and neck
cancer. Clin Cancer Res. 19:1375–1388. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Grosshans BL, Ortiz D and Novick P: Rabs
and their effectors: Achieving specificity in membrane traffic.
Proc Natl Acad Sci USA. 103:11821–11827. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cheng KW, Lu Y and Mills GB: Assay of
Rab25 function in ovarian and breast cancers. Methods Enzymol.
403:202–215. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sheach LA, Adeney EM, Kucukmetin A, et al:
Androgen-related expression of G-proteins in ovarian cancer. Br J
Cancer. 101:498–503. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tong M, Chan KW, Bao JY, et al: Rab25 is a
tumor suppressor gene with antiangiogenic and anti-invasive
activities in esophageal squamous cell carcinoma. Cancer Res.
72:6024–6035. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cheng JM, Volk L, Janaki DK, Vyakaranam S,
Ran S and Rao KA: Tumor suppressor function of Rab25 in
triple-negative breast cancer. Int J Cancer. 126:2799–2812.
2010.PubMed/NCBI
|
|
39
|
Bigelow RL, Williams BJ, Carroll JL, et
al: TIMP-1 overexpression promotes tumorigenesis of MDA-MB-231
breast cancer cells and alters expression of a subset of cancer
promoting genes in vivo distinct from those observed in vitro.
Breast Cancer Res Treat. 117:31–44. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cheng KW, Agarwal R, Mitra S, et al: Rab25
increases cellular ATP and glycogen stores protecting cancer cells
from bioenergetic stress. EMBO Mol Med. 4:125–141. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Caswell PT, Spence HJ, Parsons M, White
DP, Clark K, Cheng KW, Mills GB, Humphries MJ, Messent AJ, Anderson
KI, et al: Rab25 associates with alpha5beta1 integrin to promote
invasive migration in 3D microenvironments. Dev Cell. 13:496–510.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lapierre LA, Caldwell CM, Higginbotham JN,
Avant KM, Hall J, Beauchamp RD and Goldenring JR: Transformation of
rat intestinal epithelial cells by overexpression of Rab25 is
microtubule dependent. Cytoskeleton (Hoboken). 68:97–111. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tang BL: Is Rab25 a tumor promoter or
suppressor-context dependency on RCP status? Tumor Biol.
31:359–361. 2010. View Article : Google Scholar
|