Introduction
Gastric cancer is the fourth most common type of
cancer and the second-highest cause of cancer-related mortality,
worldwide (1). Approximately 8% of
newly diagnosed malignant tumors originate in the stomach, and
>700,000 people succumb to gastric cancer annually (2). Epidemiological studies have indicated
that the risk factors for gastric cancer include family history,
age, gender, consumption of salt-preserved foods and dietary
nitrites, gastrectomy, Helicobacter pylori (H.
pylori) infection, smoking and alcohol. Several treatment
modalities for gastric cancer are available, including surgery,
chemotherapy, radiotherapy and immunotherapy. However, the
prognosis for patients with gastric cancer remains relatively poor.
The global five-year survival rate is ~20%, which is likely to be
due to diagnosis at an advanced stage of disease and the limited
treatment options that are available. Therefore, there is a
requirement to identify novel biomarkers to aid with the early
diagnosis and treatment of gastric cancer.
Previous studies have demonstrated that aberrant
cellular metabolism is a hallmark of tumorigenesis and cancer
progression (3–5). Accumulating evidence indicates that
in vivo tumors and tumor cell lines undergo abnormal changes
in cholesterol metabolism (6,7). In theory, rapidly dividing cancer cells
utilize two major mechanisms, in order to fulfill their cholesterol
requirements (8). Cellular
requirements may be met by either de novo cholesterol
biosynthesis, or by uptake of exogenous lipoprotein-associated
cholesterol and cholesteryl esters. De novo cholesterol
biosynthesis is under tight feedback regulation in normal cells and
tissues (9). However, during
tumorigenesis, this mechanism is altered, which is likely to be a
reflection of the increased cholesterol requirements of actively
dividing tumor cells. Cholesterol metabolism shifts during
neoplasia, and absorption of exogenous cholesterol increases
because it is important for neoplasia due to the increased demand
of the tumor cells (10).
Apolipoprotein E (ApoE), secreted by hepatic and
extrahepatic cells, affects cholesterol transport, lipid metabolism
and protein synthesis, by binding to the low-density lipoprotein
receptor and the ApoE receptor on lipid particles. ApoE
participates in other cellular functions, including tissue repair,
immune response and regulation, and cell growth and differentiation
(11,12). Numerous studies have shown that ApoE
expression is associated with a number of types of tumors and tumor
cell lines (13). Recently, ApoE has
been identified as a potential tumor-associated marker for gastric
cancer, due to its elevated protein expression relative to that of
normal controls (14).
Although ApoE is known to be overexpressed in
gastric cancer, its associated genes and transcription factors
remain to be identified. The present study used Affymetrix Exon
Arrays to identify differential gene expression profiles in gastric
cancer tissues and adjacent normal tissues. Transcription
factor-gene regulatory networks were constructed through
integration of the transcriptional regulatory element database
(TRED) (15), the BioGRID database
and gene expression profiling, using Cytoscape software version
3.0.1 (Ontario Genomics Institute, Toronto, Canada). ApoE
associated genes and its transcription factor-gene regulatory
network were systematically identified. The aim of the study was to
provide insight into the pathogenesis of gastric cancer, and to
identify biomarkers in order to improve the diagnosis and treatment
of this disease.
Materials and methods
Sample collection
Five pairs of gastric cancer tissues and adjacent
noncancerous tissues were obtained at the First Hospital of Jilin
University (Changchun, China). This study was approved by the First
Hospital of Jilin University review board and each patient provided
written informed consent. All tissues were snap-frozen and stored
in liquid nitrogen within 20 min of resection. TNM cancer staging
and histological classification were performed by a pathologist,
according to the World Health Organization (WHO) criteria (16).
RNA isolation and microarray
hybridization and scanning
Briefly, a total of 15 mg RNA was extracted from
each tissue sample, using TRIzol™ (Invitrogen Life Technologies,
Carlsbad, CA, USA), followed by purification using the RNeasy Mini
kit (Qiagen, Düsseldorf, Germany), according to the manufacturer's
instructions. The A260/A280 ratio was determined by a UV2800
ultraviolet spectrophotometer (Unico, Dayton, NY, USA). RNA samples
with ratios of 1.8–2.0 were considered highly purified.
RNA samples were analyzed using the GeneChip Human
Exon 1.0 ST Array (Affymetrix, Santa Clara, CA, USA), according to
the protocol detailed in the GeneChip Expression Analysis Technical
Manual (P/N 900223). Briefly, 1 mg of RNA template was reverse
transcribed into cDNA, followed by endonuclease digestion and
labeling with the DNA labeling reagent provided by Affymetrix. The
labeled samples were mixed with hybridization cocktail and
hybridized to the microarray at 45°C with centrifugation at 1 × g
for 17 h. The array was washed and stained on the GeneChip Fluidics
Station 450, using the appropriate fluidics solutions, prior to
insertion into the Affymetrix autoloader carousel. Arrays were
scanned using the GeneChip Scanner 3000 with GeneChip Operating
Software. All instruments, chips and reagents were obtained from
Affymetrix.
Analysis of differentially expressed
genes
In order to analyze the arrays and extract raw
signal data, GeneChip Operating Software was used. After importing
raw signal data, the Limma algorithm, linear models and empirical
Bayes methods were utilized to analyze the data and identify
differentially expressed genes. Stringent criteria were used in
order to prevent very small fold changes from being judged as
differentially expressed as a result of small residual standard
deviations. The resulting P-values were adjusted using the
Benjamini and Hochberg false discovery rate (BH-FDR) algorithm
(17). Gene expression was considered
to be significantly different if both FDR values were <0.05,
limiting the FDR to ≤5%, and gene expression exhibited a ≥2-fold
change between cancer and the corresponding normal tissues; that
is, log2FC >1 or log2FC <-1, and P-value <0.05.
Construction of a transcription factor
gene network using gene expression profiling, TRED and the BioGRID
database
A transcription factor (TF)-gene network was
constructed, based on gene expression profiling, TRED and the
BioGRID database, using cytoscape software, according to the
regulatory interactions and the differential expression values of
each TF and gene. Attributing associations among all genes and TFs
created the adjacent matrix. The resulting analysis is presented in
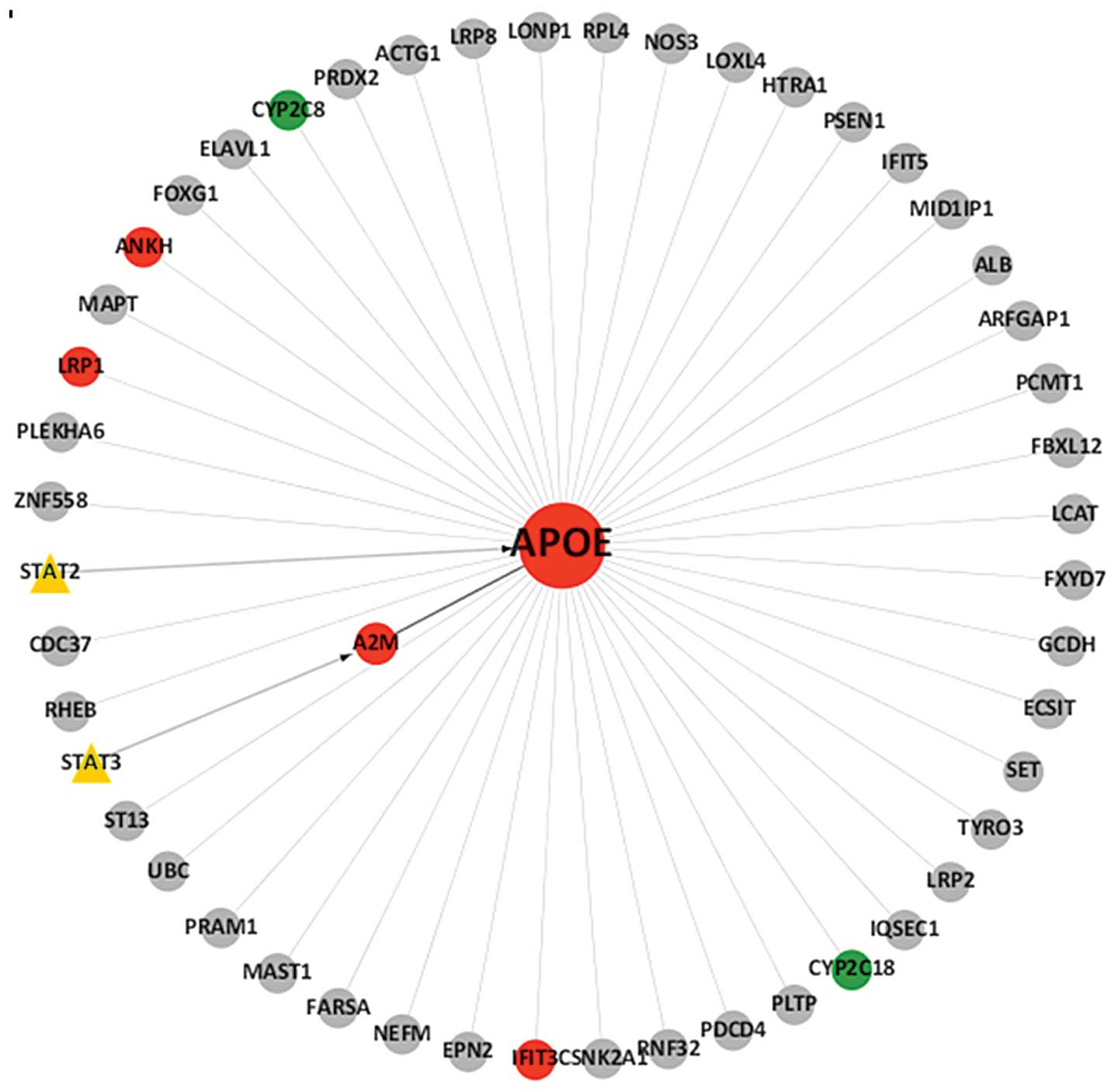
Fig. 1.
Functional enrichment analysis of
genes
The database for annotation, visualization and
integrated discovery (DAVID) functional annotation software was
applied to analyze the functional enrichment of aberrant genes. The
‘GENETIC_ASSOCIATION_DB_DISEASE_CLASS’ and ‘GENE ONTOLOGY’ options
provided information about disease association enrichment and
functional enrichment of gene clusters. The
‘GENETIC_ASSOCIATION_DB_DISEASE_CLASS’ was selected in order to
identify disease class enrichment, and ‘GOTERM_MF_FAT’ was used to
identify functional enrichment, with the Benjamini method for
determining a significant enrichment score ≥1.3.
Results and Discussion
In order to identify differentially expressed genes
in gastric cancer, Affymatrix Exon Arrays containing 17,800 human
genes were utilized, and five pairs of gastric tumor tissues and
adjacent normal tissues were analyzed. A total of 1,224 genes
demonstrated a ≥2-fold change in expression in tumor tissues
relative to that of adjacent normal tissues (data not shown). Among
these differentially expressed genes, 730 were upregulated while
495 were downregulated. Specifically, the expression of ApoE was
greater (log2FC=1.345) in gastric cancer tissues compared with that
adjacent normal tissues (P<0.01). A previous study demonstrated
that ApoE was upregulated in gastric cancer tissues, and that its
overexpression was associated with a poor prognosis in patients
with gastric cancer (18). Therefore,
ApoE-associated genes and its regulatory network in gastric cancer
tissues, were further investigated using in silico
analyses.
In order to identify genes and TFs associated with
ApoE-overexpression, the BioGRID database was utilized.
Additionally, TRED was used to analyze cis- and trans-regulatory
elements that have been identified in mammals. Thus, integration of
the analyses from the gene expression profiling, and the BioGRID
and TRED databases was used to analyze ApoE-associated genes. The
results demonstrated that eight genes, namely signal transducer and
activator of transcription 2 (STAT2), STAT3, low-density
lipoprotein receptor-related protein 1 (LRP1), α2-macroglobulin
(A2M), interferon-induced protein with tetratricopeptide repeats 3
(IFIT3), ankylosis progressive homolog (ANKH), and the cytochrome
P450 genes, CYP2C8 and CYP2C18, were associated with ApoE and were
differentially expressed, with six of these found to be upregulated
and two downregulated (Table I).
Ultimately, TF-gene regulatory networks centered on ApoE, were
established for the gastric cancer tissues examined in the present
study (Fig. 1).
 | Table I.Summary of eight differentially
expressed genes related to apolipoprotein E in gastric cancer
tissues. |
Table I.
Summary of eight differentially
expressed genes related to apolipoprotein E in gastric cancer
tissues.
| Gene symbol | log2FC | Diseases | Entrez gene name | Location | Family |
|---|
| STAT2 | 1.205043 | Antigen Presentation,
Cancer, Cardiovascular Disease, Connective Tissue Disorders,
Dermatological Diseases and Conditions, Endocrine System Disorders,
Gastrointestinal Disease, Genetic Disorder, Hematological Disease,
Hepatic System Disease, Immunological Disease, Infection Mechanism,
Infectious Disease, Inflammatory Disease, Inflammatory Response,
Neurological Disease, Ophthalmic Disease, Organismal Injury and
Abnormalities, Reproductive System Disease, Respiratory Disease,
Skeletal and Muscular Disorders | Signal transducer and
activator of transcription 2 1,91 kDa | Nucleus | Transcription
regulator |
| STAT3 | 1.179164 | Antigen Presentation,
Cancer, Cardiovascular Disease, Connective Tissue Disorders,
Dermatological Diseases and Conditions, Endocrine System Disorders,
Gastrointestinal Disease, Genetic Disorder, Hematological Disease,
Hepatic System Disease, Immunological Disease, Infection Mechanism,
Infectious Disease, Inflammatory Disease, Inflammatory Response,
Neurological Disease, Ophthalmic Disease, Organismal Injury and
Abnormalities, Reproductive System Disease, Respiratory Disease,
Skeletal and Muscular Disorders | Signal transducer and
activator of transcription 3 1,91 kDa | Nucleus | Transcription
regulator |
| A2M | 1.450473 | Antigen Presentation,
Endocrine System Disorders, Genetic Disorder, Infectious Disease,
Inflammatory Disease, Inflammatory Response, Metabolic Disease,
Neurological Disease | α2-macroglobulin | Extracellular
space | Transporter |
| IFIT3 | 1.575557 | Cancer, Organismal
Injury and Abnormalities, Reproductive System Disease | Interferon-induced
protein with tetratricopeptide repeats 3 | Cytoplasm | Other |
| ANKH | 1.040222 | Connective Tissue
Disorders, Developmental Disorder, Genetic Disorder, Inflammatory
Disease, Organismal Injury and Abnormalities, Skeletal and Muscular
Disorders | Ankylosis,
progressive homolog (mouse) | Plasma membrane | Transporter |
| LRP1 | 1.320753 | Antigen Presentation,
Cancer, Cardiovascular Disease, Endocrine System Disorders, Genetic
Disorder, Hematological Disease, Inflammatory Response, Metabolic
Disease, Neurological Disease, Nutritional Disease, Organismal
Injury and Abnormalities, Psychological Disorders, Skeletal and
Muscular Disorders | Low density
lipoprotein-related protein 1 (α2-macroglobulin receptor) | Plasma
membrane | Transmembrane
receptor |
| CYP2C8 | −1.774129 | (Unknown or not
applicable) | Cytochrome P450,
family 2, subfamily C, polypeptide 8 | Cytoplasm | Enzyme |
| CYP2C18 | −2.802344 | (Unknown or not
applicable) | Cytochrome P450,
family 2, subfamily C, polypeptide 18 | Cytoplasm | Enzyme |
In addition, DAVID was utilized to provide a
functional enrichment analysis of these eight differentially
expressed genes. Using DAVID, the JAK-STAT cascade, acute-phase
response, acute inflammatory response and steroid hormone response
genes were identified as significantly enriched (Table II). These eight aberrantly expressed
genes were categorized into immune, pharmacogenomic, metabolic and
cardiovascular disease classes (Table
III).
| Table II.Functional enrichment analysis of
genes in the regulatory network. |
Table II.
Functional enrichment analysis of
genes in the regulatory network.
| Category | Term | P-value | Genes | Fold
enrichment | Benjamini |
|---|
| GO:0007259 | JAK-STAT
cascade |
1.4×10−2 | STAT3, STAT2 | 115.62 |
8.8×10−1 |
| GO:0006953 | Acute-phase
response |
1.5×10−2 | A2M, STAT3 | 112.73 |
6.6×10−1 |
| GO:0002526 | Acute inflammatory
response |
3.6×10−2 | A2M, STAT3 |
46.014 |
8.3×10−1 |
| GO:0048545 | Steroid hormone
response |
6.9×10−2 | A2M, STAT3 | 23.4 |
9.3×10−1 |
| Table III.Disease class enrichment analysis of
genes in the TF-gene regulatory network. |
Table III.
Disease class enrichment analysis of
genes in the TF-gene regulatory network.
| Term | P-value | Genes | Fold
enrichment | Benjamini |
|---|
| Immune |
5.8×10−2 | A2M, LRP1, ANKH,
STAT3 | 3.053 |
3.0×10−1 |
|
Pharmacogenomic |
6.0×10−2 | LRP1, CYP2C8,
STAT3 |
5.5436 |
2.2×10−1 |
| Metabolic |
6.6×10−2 | A2M, LRP1, CYP2C8,
ANKH |
2.9038 |
1.8×10−1 |
| Cardiovascular |
2.5×10−3 | A2M, LRP1, CYP2C8,
ANKH, STAT3 |
0.02926 |
2.9×10−2 |
LRP1, also termed ApoE-specific lipoprotein
receptor, or ApoE receptor, is a cell-surface protein that is
involved in the metabolism of cholesterol, by mediating the
endocytosis of ApoE-containing lipoproteins from plasma into cells.
ApoE secreted from cancer cells, suppresses invasion and metastatic
endothelial recruitment by engaging LRP1 and LRP8 receptors,
respectively. The function of LRP1 in the regulation of tumor
growth has been well documented. Studies have shown that increased
LRP1 expression correlates with high levels of invasion (19) and, conversely, silencing of LRP1
prevents the spread of malignant cells (20). In gliomas, LRP1 expression in tumors
greatly exceeded that in normal brain tissues (21) and its expression was found to be
correlated with tumor aggressiveness (22). The abundant expression of LRP1 mRNA
suggests that it may be involved in the uptake of ApoE phospholipid
discoidal particles or ApoE-enriched high-density lipoprotein in
gastric cancer.
LRP1 is an A2M receptor. Recent studies have
demonstrated that A2M may also regulate cell signal transduction
via LRP1 (23–25). A2M is responsible for the binding and
inactivation of plasma proteases, as well as the transport of
various cytokines, growth factors and hormones (26). ApoE is non-covalently bound to A2M in
human plasma, and therefore, the present in silico analysis
of ApoE binding may provide insights into the pathogenic and
intracellular role of ApoE in cancer cells. In addition, A2M has
previously been reported as a candidate biomarker for the early
diagnosis in numerous types of cancers, including gastric cancer
(27–29).
Additional genes, including IFIT3, ANKH, CYP2C18 and
CYP2C8, were demonstrated to be correlated with ApoE expression in
the present study, and have also previously been linked to ApoE
expression in Alzheimer's disease (30). IFIT3 inhibits cell migration and shows
marked antiproliferative effects (31). Overexpression of IFIT3 has been shown
to induce tumor proliferation, angiogenesis and chemoresistance in
pancreatic carcinoma cells (32).
ANKH is a transmembrane protein that transports intracellular
pyrophosphate to the extracellular milieu (33) and has been demonstrated to be
overexpressed in bladder cancer (34)
and small cell lung cancer cell lines (35). CYP450 has been shown to be
downregulated in hepatocytes in response to inflammation and
infection (36). Local chronic
inflammation is hypothesized to contribute to tumorigenesis,
particularly in gastric cancer that is associated with H.
pylori infection (37). The
present study also indicated that CYP2C18 and CYP2C8 were
downregulated in gastric cancer tissues. A separate study
demonstrated that CYP2C18 was associated with the development of
gastric cancer (38).
In order to identify ApoE regulatory TFs, TRED was
used. The results demonstrated that ApoE may be regulated directly
by STAT2, or indirectly by STAT3. In order to gain an improved
understanding of the regulatory network, a brief framework of the
network was configured (Fig. 1).
STAT2 and STAT3 are members of the signal transducer and activator
of transcription family. STAT3 is involved in selectively inducing
and maintaining a procarcinogenic inflammatory microenvironment
that promotes tumor cell transformation (39). The JAK-STAT signaling pathway is known
to regulate genes that are involved in the regulation of cell
proliferation, differentiation and apoptosis, by transducing
signals from the cell membrane to the nucleus (40). Targeting the JAK-STAT3 signaling
pathway, and specifically STAT3, has been hypothesized to be a
potential therapeutic strategy for cancer (41). STAT2 has been identified as a novel
contributor to carcinogensis, and may increase the gene expression
and secretion of proinflammatory mediators, thereby activating the
oncogenic STAT3 signaling pathway (42). The current data demonstrated
significantly increased levels of STAT3 and STAT2 in gastric cancer
tissues. However, further investigation of the importance of
JAK-STAT activation in this disease is required.
The present study demonstrated that a combination of
interaction discovery experiments and computational analyses from
diverse biological data, may help to identify causative genes in
gastric cancer. In particular, ApoE as was identified as a
potential biomarker of this disease. Although further studies are
required, these findings indicate a role for ApoE in the
development of gastric cancer.
Acknowledgements
The authors would like to thank all those who
participated in this study. Particular thanks is extended to Dr
Quan Lin (The First Hospital of Jilin University), for assistance
with sample collection and to the five anonymous patients involved
in this study. The authors would also like to thank Medjaden
Bioscience ltd. for editing and proofreading this manuscript.
References
|
1
|
Cui J, Chen Y, Chou WC, Sun L, Chen L, Suo
J, Ni Z, Zhang M, Kong X, Hoffman LL, et al: An integrated
transcriptomic and computational analysis for biomarker
identification in gastric cancer. Nucleic Acids Res. 39:1197–1207.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee HJ, Song IC, Yun HJ, Jo DY and Kim S:
CXC chemokines and chemokine receptors in gastric cancer: From
basic findings towards therapeutic targeting. World J Gastroentero.
20:1681–1693. 2014. View Article : Google Scholar
|
|
3
|
DeBerardinis RJ, Lum JJ, Hatzivassiliou G
and Thompson CB: The biology of cancer: Metabolic reprogramming
fuels cell growth and proliferation. Cell Metab. 7:11–20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hsu PP and Sabatini DM: Cancer Cell
Metabolism: Warburg and beyond. Cell. 134:703–707. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ríos-Marco P, Martín-Fernández M,
Soria-Bretones I, Ríos A, Carrasco MP and Marco C:
Alkylphospholipids deregulate cholesterol metabolism and induce
cell-cycle arrest and autophagy in U-87 MG glioblastoma cells.
Biochim Biophys Acta. 1831:1322–1334. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pussinen PJ, Karten B, Wintersperger A,
Reicher H, McLean M, Malle E and Sattler W: The human breast
carcinoma cell line HBL-100 acquires exogenous cholesterol from
high-density lipoprotein via CLA-1 (CD-36 and LIMPII analogous
1)-mediated selective cholesteryl ester uptake. Biochem J.
349:559–566. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
de Weille J, Fabre C and Bakalara N:
Oxysterols in cancer cell proliferation and death. Biochem
Pharmacol. 86:154–160. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tosi MR and Tugnoli V: Cholesteryl esters
in malignancy. Clin Chim Acta. 359:27–45. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Goldstein JL and Brown MS: The low-density
lipoprotein pathway and its relation to atherosclerosis. Annu Rev
Biochem. 46:897–930. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Coleman PS, Chen LC and Sepp-Lorenzino L:
Cholesterol Metabolism and Tumor Cell Proliferation(Cholesterol:
Its Functions and Metabolism in Biology and Medicine). Bittman R:
Subcellular Biochemistry. 28. Plenum Press; New York: pp. 363–435.
1997, PubMed/NCBI
|
|
11
|
Weisgraber KH, Rall SC Jr and Mahley RW:
Human E apoprotein heterogeneity. Cysteine-arginine interchanges in
the amino acid sequence of the apo-E isoforms. J Biol Chem.
256:9077–9083. 1981.PubMed/NCBI
|
|
12
|
Rall SC Jr, Weisgraber KH, Innerarity TL
and Mahley RW: Structural basis for receptor binding heterogeneity
of apolipoprotein E from type III hyperlipoproteinemic subjects.
Proc Natl Acad Sci USA. 79:4696–4700. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
El Roz A, Bard JM, Valin S, Huvelin JM and
Nazih H: Macrophage apolipoprotein E and proliferation of MCF-7
breast cancer cells: Role of LXR. Anticancer Res. 33:3783–3789.
2013.PubMed/NCBI
|
|
14
|
Sakashita K, Tanaka F, Zhang X, Mimori K,
Kamohara Y, Inoue H, Sawada T, Hirakawa K and Mori M: Clinical
significance of ApoE expression in human gastric cancer. Oncol Rep.
20:1313–1319. 2008.PubMed/NCBI
|
|
15
|
Jiang C, Xuan Z, Zhao F and Zhang MQ:
TRED: A transcriptional regulatory element database, new entries
and other development. Nucleic Acids Res. 35 (Database
Issue):D137–D140. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zurleni T, Gjoni E, Ballabio A, Casieri R,
Ceriani P, Marzoli L and Zurleni F: Sixth and seventh
tumor-node-metastasis staging system compared in gastric cancer
patients. World J Gastrointest Surg. 5:287–293. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tan YD and Xu H: A general method for
accurate estimation of false discovery rates in identification of
differentially expressed genes. Bioinformatics. 30:2018–2025. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
De Feo E, Simone B, Persiani R, Cananzi F,
Biondi A, Arzani D, Amore R, D'Ugo D, Ricciardi G and Boccia S: A
case-control study on the effect of Apolipoprotein E genotypes on
gastric cancer risk and progression. BMC Cancer. 12:4942012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Song H, Li Y, Lee J, Schwartz AL and Bu G:
Low-density lipoprotein receptor-related protein 1 promotes cancer
cell migration and invasion by inducing the expression of matrix
metalloproteinases 2 and 9. Cancer Res. 69:879–886. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dedieu S, Langlois B, Devy J, Sid B,
Henriet P, Sartelet H, Bellon G, Emonard H and Martiny L: LRP-1
silencing prevents malignant cell invasion despite increased
pericellular proteolytic activities. Mol Cell Biol. 28:2980–2995.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lopes MB, Bogaev CA, Gonias SL and
VandenBerg SR: Expression of alpha 2-macroglobulin receptor/low
density lipoprotein receptor-related protein is increased in
reactive and neoplastic glial cells. FEBS Lett. 338:301–305. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yamamoto M, Ikeda K, Ohshima K, Tsugu H,
Kimura H and Tomonaga M: Increased expression of low density
lipoprotein receptor-related protein/alpha2-macroglobulin receptor
in human malignant astrocytomas. Cancer Res. 57:2799–2805.
1997.PubMed/NCBI
|
|
23
|
Barcelona PF, Luna JD, Chiabrando GA,
Juarez CP, Bhutto IA, Baba T, McLeod DS, Sánchez MC and Lutty GA:
Immunohistochemical localization of low density lipoprotein
receptor-related protein 1 and alpha (2)-Macroglobulin in retinal
and choroidal tissue of proliferative retinopathies. Exp Eye Res.
91:264–272. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bonacci GR, Cáceres LC, Sánchez MC and
Chiabrando GA: Activated alpha(2)-macroglobulin induces cell
proliferation and mitogen-activated protein kinase activation by
LRP-1 in the J774 macrophage-derived cell line. Arch Biochem
Biophys. 460:100–106. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qiu Z, Strickland DK, Hyman BT and Rebeck
GW: alpha 2-Macroglobulin exposure reduces calcium responses to
N-methyl-D-aspartate via low density lipoprotein receptor-related
protein in cultured hippocampal neurons. J Biol Chem.
277:14458–14466. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Andersson M, Jönsson U and Olsson A: A
slow form of alpha-2-macroglobulin in diseased and healthy dogs. J
Comp Pathol. 127:37–44. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hudler P, Kocevar N and Komel R: Proteomic
approaches in biomarker discovery: New perspectives in cancer
diagnostics. ScientificWorldJournal. 2014:2603482014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ahn HS, Shin YS, Park PJ, Kang KN, Kim Y,
Lee HJ, Yang HK and Kim CW: Serum biomarker panels for the
diagnosis of gastric adenocarcinoma. Br J Cancer. 106:733–739.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hanas JS, Hocker JR, Cheung JY, Larabee
JL, Lerner MR, Lightfoot SA, Morgan DL, Denson KD, Prejeant KC,
Gusev Y, et al: Biomarker identification in human pancreatic cancer
sera. Pancreas. 36:61–69. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Soler-López M, Zanzoni A, Lluís R, Stelzl
U and Aloy P: Interactome mapping suggests new mechanistic details
underlying Alzheimer's disease. Genome Res. 21:364–376. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fensterl V and Sen GC: The ISG56/IFIT1
gene family. J Interf Cytok Res. 31:71–78. 2011. View Article : Google Scholar
|
|
32
|
Camaj P, Ischenko I, Seeliger H, Arnold G,
Jauch KW and Bruns CJ: Overexpression of the gene IFIT3 enhances
tumor growth, angiogenesis, metastasing and chemoresistance of the
pancreas carcinoma cells(Chirurgisches Forum und DGAV Forum 2009).
Schumpelick V, Bruch HP and Schackert HK: Deutsche Gesellschaft für
Chirurgie. 38. Springer; Berlin Heidelberg: pp. 17–18. 2009
|
|
33
|
Kirsch T, Kim HJ and Winkles JA:
Progressive ankylosis gene (ank) regulates osteoblast
differentiation. Cells Tissues Organs. 189:158–162. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zheng M, Simon R, Mirlacher M, Maurer R,
Gasser T, Forster T, Diener PA, Mihatsch MJ, Sauter G and Schraml
P: TRIO amplification and abundant mRNA expression is associated
with invasive tumor growth and rapid tumor cell proliferation in
urinary bladder cancer. Am J Pathol. 165:63–69. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang A, Zheng C, Hou M, Lindvall C,
Wallin KL, Angström T, Yang X, Hellström AC, Blennow E, Björkholm
M, et al: Amplification of the telomerase reverse transcriptase
(hTERT) gene in cervical carcinomas. Genes Chromosomes Cancer.
34:269–275. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Aitken AE and Morgan ET: Gene-specific
effects of inflammatory cytokines on cytochrome P450 2C, 2B6 and
3A4 mRNA levels in human hepatocytes. Drug Metab Dispos.
35:1687–1693. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yamaoka Y and Graham DY: Helicobacter
pylori virulence and cancer pathogenesis. Future Oncol.
10:1487–1500. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hu K and Chen F: Identification of
significant pathways in gastric cancer based on protein-protein
interaction networks and cluster analysis. Genet Mol Biol.
35:701–708. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Saturnino C, Palladino C, Napoli M,
Sinicropi MS, Botta A, Sala M, Carcereri de Prati A, Novellino E
and Suzuki H: Synthesis and biological evaluation of new
N-alkylcarbazole derivatives as STAT3 inhibitors: Preliminary
study. Eur J Med Chem. 60:112–119. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Silver-Morse L and Li WX: JAK-STAT in
heterochromatin and genome stability. JAKSTAT.
2:e260902013.PubMed/NCBI
|
|
41
|
Bournazou E and Bromberg J: Targeting the
tumor microenvironment: JAK-STAT3 signaling. JAKSTAT.
2:e238282013.PubMed/NCBI
|
|
42
|
Gamero AM, Young MR, Mentor-Marcel R, Bobe
G, Scarzello AJ, Wise J and Colburn NH: STAT2 contributes to
promotion of colorectal and skin carcinogenesis. Cancer Prev Res
(Phila). 3:495–504. 2010. View Article : Google Scholar : PubMed/NCBI
|