Introduction
Osteosarcoma is the most common type of primary bone
tumor in adolescents and young adults (1,2), and is
characterized by an abundance of genomic aberrations (3). According to previous studies, the 5-year
survival rate of osteosarcoma is 60–70%, and the prognosis has not
significantly improved over the last 30 years (4). Despite improvements in osteosarcoma
treatment, the molecular mechanism underlying osteosarcoma
development remains unclear. Therefore, it is important to explore
the molecular mechanism of osteosarcoma development to additionally
improve osteosarcoma treatment. However, the mechanisms of
osteosarcoma development are complex, and numerous factors,
including genes associated with osteosarcoma (3), copy number variations (CNVs) across the
whole genome (5) and miRNAs (6), may contribute to the development of
osteosarcoma. In particular, CNV may contribute to the development
of various types of cancer, including osteosarcoma and B-cell
lymphoma (7,8). CNV has been considered as a marker for
cancer predisposition (9). CNV may
contribute to the pathogenesis of osteosarcoma (10) and the disease risk for diffuse large
B-cell lymphoma (11).
Furthermore, a number of previous studies revealed
that microRNAs (miRNAs/miRs) are associated with the development of
osteosarcoma (12,13), and may be used as the molecular
targets for osteosarcoma, including miR-143, miR-382 and miR-214.
Downregulation of miR-143 is associated with the lung metastasis of
osteosarcoma via the upregulated expression of matrix
metalloproteinase 13 (14).
Overexpression of miR-382 may suppress the metastasis of
osteosarcoma (15). Upregulated
expression of miR-214 may contribute to the pathogenesis of
osteosarcoma and may be associated with adverse prognosis (16). In addition, miRNAs, including miR-143
(17), miR-27a (18), miR-223 (19), miR-191 (20), miR-133a (21) and miR-26a (22), may also be associated with the
development of osteosarcoma. These results indicated that miRNAs
may serve an important role in the pathogenesis of
osteosarcoma.
Previously, miRNAs expression profiles (23,24) and
SNP microarray (1) were used to
identify the genes, miRNAs, and single nucleotide polymorphisms
(SNPs) associated with the development of osteosarcoma. In certain
studies, specific genes associated with osteosarcoma development
were identified by the integrative analysis of copy number and gene
expression data (25). Based on the
aforementioned, comprehensive analysis of microRNA (GSE65071) and
CNV (GSE33153) microarray data were performed in the present study
to identify the miRNAs, and genes involved in the pathogenesis of
osteosarcoma. In addition, a miRNA-gene regulatory network of the
overlapping genes of the genes in CNV regions and target genes of
differentially expressed miRNAs was constructed. These miRNAs and
genes associated with osteosarcoma development were identified in
order to additionally explore the mechanism of osteosarcoma, and
may be used as candidate target miRNAs and genes for gene
therapy.
Materials and methods
Microarray data
Microarray data (GSE65071 and GSE33153) were
downloaded from Gene Expression Omnibus (GEO, http://www.ncbi.nlm.nih.gov/geo/). For GSE65071,
there were 20 osteosarcoma samples and 10 healthy control samples,
and the platform was GPL19631Exiqon human V3 microRNA PCR panel
I+II. A total of 32 osteosarcoma samples were available on
GSE33153, of which the platform was GPL6801 [GenomeWideSNP_6]
Affymetrix Genome-Wide Human SNP 6.0 Array.
Data preprocessing and analysis of
copy number variation
GSE65071 was preprocessed using preprocessCore
package v1.38.1 (http://www.bioconductor.org/packages/release/bioc/html/preprocessCore.html)
in R language. Then, the differentially expressed miRNAs between
the osteosarcoma and control groups were calculated using the Limma
package v3.32.2 (http://www.bioconductor.org/packages/release/bioc/html/limma.html),
and the adjusted P-value <0.05 and |log (fold change) |>2
were chosen as the cut-off criterion. Then, two-way clustering
analysis was performed using the gplots package v3.0.1 (https://cran.r-project.org/web/packages/gplots/index.html).
For GSE33153, the log R Ratio (LRR) and B allele
frequency (BAF) were extracted using the affy2sv package v1.0.12
(https://bitbucket.org/brge/affy2sv),
and then CNV calling was performed using PennCNV v2014 (07)
(http://penncnv.openbioinformatics.org/). CNV
identified in ≥4 samples was considered as CNV regions, which may
be involved in the development of osteosarcoma.
Identification of target genes of
miRNAs
The regulatory associations between long non-coding
RNA, miRNA, competing endogenous RNA, mRNA and RNA binding proteins
may be identified using the starBase database (http://starbase.sysu.edu.cn/), which included the
clinical data and expression profiles of 14 types of cancer. The
target genes of differentially expressed miRNAs were identified
using the starBase database. The cut-off criterion of miRNA-gene
pairs, which were used to screen the target genes, was listed as
follows: i) miRNA-gene pairs were confirmed by ≥2 experiments; and
ii) the change in the trend of miRNA and target gene expression
values was the opposite.
Comprehensive analysis of miRNAs and
CNV regions
Genes in the CNV regions were screened using
scan_region.pl of PennCNV. The overlap between genes contained
within the CNV regions and targets of the differentially expressed
miRNAs were obtained. miRNA-gene pairs that overlapped were
obtained based on the starBase database, and a miRNA-gene
regulatory network was constructed.
Results
Analysis of differentially expressed
miRNA and CNV regions
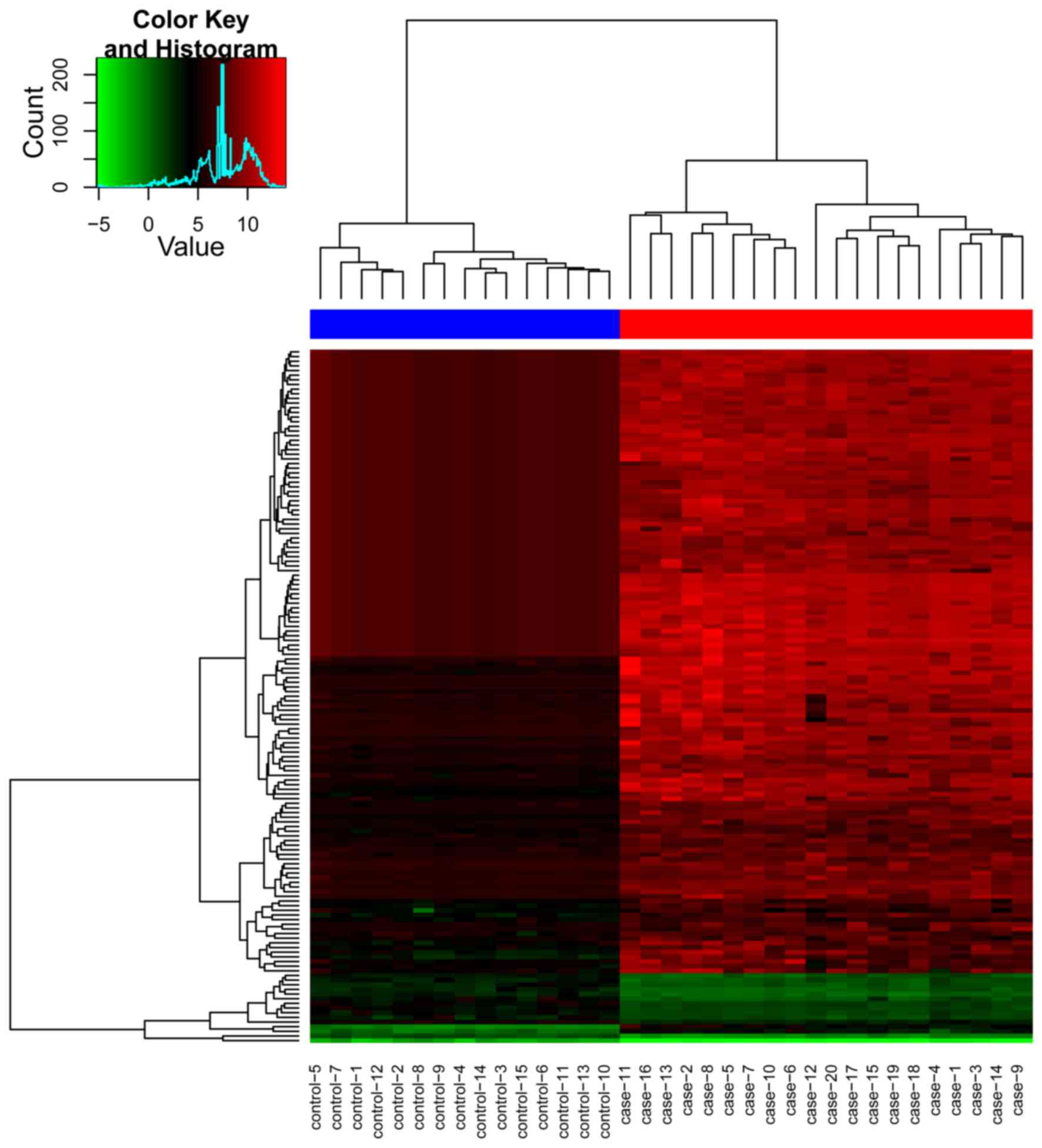
A total of 149 differentially expressed miRNAs
between the osteosarcoma group and the control group were
identified, including 13 downregulated, and 136 upregulated
differentially expressed miRNAs (Table
I). The clustering plot indicated that the differentially
expressed miRNAs evidently separated the osteosarcoma samples from
the control samples (Fig. 1).
 | Table I.Top ten differentially expressed
miRNAs. |
Table I.
Top ten differentially expressed
miRNAs.
| miRNAs | logFC | P-value | Adjusted
P-value |
|---|
| hsa-miR-624-5p | 4.998135150 |
2.08×10−30 |
7.41×10−28 |
| hsa-miR-505-5p | 3.631886325 |
2.96×10−28 |
5.27×10−26 |
|
hsa-let-7f-2-3p | 3.613251767 |
4.96×10−28 |
5.89×10−26 |
| hsa-miR-877-3p | 4.267538662 |
4.38×10−27 |
3.89×10−25 |
| hsa-miR-183-5p | 4.469024403 |
1.94×10−26 |
1.26×10−24 |
| hsa-miR-342-5p | 5.006618179 |
2.12×10−26 |
1.26×10−24 |
|
hsa-let-7f-1-3p | 3.182939079 |
8.79×10−26 |
4.47×10−24 |
| hsa-miR-671-5p | 5.476974410 |
1.58×10−25 |
7.02×10−24 |
| hsa-miR-95 | 5.163185811 |
2.96×10−25 |
1.13×10−23 |
|
hsa-miR-499a-5p | 5.429803038 |
3.18×10−25 |
1.13×10−23 |
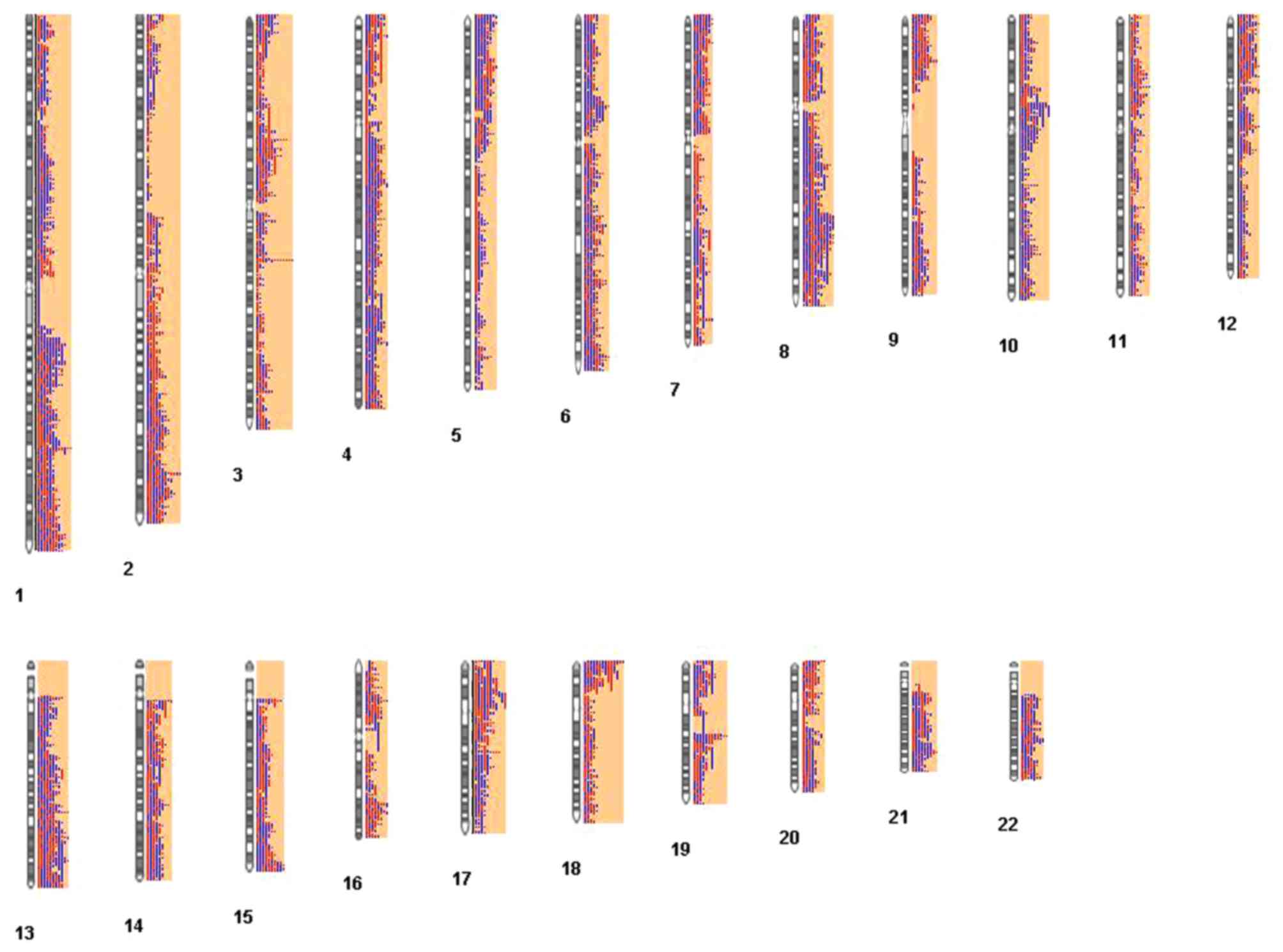
For GSE33153, 987 CNV regions were identified. The
distribution of the 987 CNV regions among chromosomes is presented
in Fig. 2. In Fig. 2, the red and blue column represents
the deletions, and overlap of copy number, respectively. The longer
the column, the higher the number of samples that exhibited
deletions or overlapping in these CNV regions.
Analysis of target genes of
differentially expressed miRNAs
In total, 7,313 miRNA-gene pairs were screened for
149 differentially expressed miRNAs, and 5 differentially expressed
miRNAs (hsa-miR-200b-3p, hsa-miR-200a-3p, hsa-miR-429,
hsa-miR-34a-5p, and hsa-miR-9-5p) with corresponding target genes
were identified, as summarized in Table
II.
| Table II.miRNA-target gene pairs. |
Table II.
miRNA-target gene pairs.
| miRNAs | Counts | Target
genesa |
|---|
|
hsa-miR-200b-3p | 282 | ICK, NPTX1,
DPY19L1, RECK, TRIO, DDX3Y, GALNT2, R3HDM2, PPM1B, TIMP2 |
|
hsa-miR-200a-3p | 159 | GIGYF1, GIGYF1,
MTF2, CASC4, ULK2, MBNL3, KPNA4, ZEB2, ZEB2, TET3 |
| hsa-miR-429 | 235 | SEC24A, TRIM52,
DPY19L1, PAPOLA, GALNT2, KIAA1432, MEX3C, PLCG1, LRIG1, DCBLD2 |
| hsa-miR-34a-5p | 92 | TNRC18, SIDT2,
ARPP19, NAA50, RCAN1, DNAJB1, XPO5, HCN3, SIRT1, MBD6 |
| hsa-miR-9-5p | 144 | ULK2, SMARCE1,
ZC3H10, VAT1, BCAT2, DICER1, ZC3H12A, GTPBP3, SH2B3, ITM2B |
Comprehensive analysis of
differentially expressed miRNA and CNV regions
A total of 761/3,635 genes in 987 CNV regions and
the genes of 7,313 miRNA-gene pairs were identified overlap. The
miRNAs with highest degrees of overlap (hsa-miR-27a-3p degree, 112;
hsa-miR-124-3p degree, 102; hsa-miR-9-5p degree, 90; hsa-miR-182-5p
degree, 79; hsa-miR-26a-5p degree, 79) and genes (FRS2
degree, 14; CORO1C degree, 12; FOXP1 degree, 11;
CPEB4 degree, 11; GLCCI1 degree, 10) are presented in
Table III.
| Table III.miRNAs and genes in the miRNA-gene
regulatory network. |
Table III.
miRNAs and genes in the miRNA-gene
regulatory network.
| A, miRNAs in the
miRNA-gene regulatory network |
|---|
|
|---|
| Marker | Degrees | Counts | Target
genesa |
|---|
| hsa-miR-27a-3p | 112 | 51 | TNRC18, PPARA,
ACLY, DNAJB9, NR2F2, HMGCS1, SGMS1, CKAP4, RPN2, CALM |
| hsa-miR-124-3p | 102 | 42 | SGMS1, DRAM1,
SUCLG2, FRS2, NUDCD2, FAR1, PIP4K2C, SERTAD3, SH3PXD2A, KLHL24 |
| hsa-miR-9-5p | 90 | 39 | FREM2, SMARCE1,
KLHDC10, DYRK1B, SLC39A14, FURIN, BAHD1, MAPKAPK2, AP2M1, BCL6 |
| hsa-miR-182-5p | 79 | 38 | RTN4, STARD13,
FRS2, KIAA1217, CNOT6, EXOC4, SNAP23, KCMF1, QSER1, SYPL1,
BDNF |
| hsa-miR-26a-5p | 79 | 29 | SRP19, MARK1,
SEMA6D, ACSL3, EIF4G2, LSM12, MAPK6, CCNJL, MFHAS1, COX5A |
| hsa-miR-429 | 79 | 32 | SUZ12, CRKL, TOB1,
FXR1, LIN7B, EVI5, GLCCI1, FRS2, ARID4B, SSH2 |
| hsa-miR-141-3p | 71 | 30 | RHEB, LENG8,
ATP2A2, BAHD1, GLCCI1, ATP1B1, PTPRG, MBTD1, GRIN2D, STAT5B |
| hsa-miR-96-5p | 70 | 33 | ARPP19, CELSR1,
KIAA1217, DOCK1, CAPNS1, CCNG1, PROK2, APPL1, PGAP1, SHC1 |
| hsa-miR-34a-5p | 63 | 30 | HCN3, HECW2, STC1,
MAP2K1, APH1A, NUMBL, NFE2L1, GREM2, ARID4B, LGR4 |
|
hsa-miR-200a-3p | 63 | 27 | RHEB, NCKAP5,
B3GNT5, LENG8, CALU, ATP2A2, BAHD1, NRP1, SPAG9, GLCCI1 |
|
| B, Genes in the
miRNA-gene regulatory network |
|
| Marker | Degrees | Counts | Target genes |
|
| FRS2 | 14 | – | – |
| CORO1C | 12 | – | – |
| FOXP1 | 11 | – | – |
| CPEB4 | 11 | – | – |
| GLCCI1 | 10 | – | – |
| CELF1 | 10 | – | – |
| MET | 10 | – | – |
| ZFHX4 |
9 | – | – |
| DOCK4 |
8 | – | – |
| MYH10 |
8 | – | – |
Discussion
Osteosarcoma is characterized by an abundance of
genomic aberrations (3). In the
present study, a comprehensive analysis of microRNA data and CNV
microarray data was performed to identify miRNAs, and genes
associated with the pathogenesis of osteosarcoma. A total of 149
differentially expressed miRNAs, including 13 down- and 136
upregulated, in the GEO GSE65071 dataset were identified. For the
GEO GSE33153 dataset, 987 CNV regions were identified. In addition,
a miRNA-gene regulatory network of 761 overlapping genes out of
3,635 genes in 987 CNV regions and the genes in 7,313 miRNA-gene
pairs was constructed. Concurrently, miRNAs (hsa-miR-27a-3p,
hsa-miR-124-3p, hsa-miR-9-5p, hsa-miR-182-5p, hsa-miR-26a-5p) and
genes [fibroblast growth factor receptor substrate 2 (FRS2),
coronin 1C (CORO1C), forkhead Box P1 (FOXP1),
cytoplasmic polyadenylation element binding protein 4
(CPEB4) and glucocorticoid induced 1 (GLCCI1)] with
the highest degree of overlap in the miRNA-gene regulatory network
were identified. Higher degrees represent an increased level of
association to the development of osteosarcoma. Therefore,
hsa-miR-27a-3p, hsa-miR-124-3p, hsa-miR-9-5p, hsa-miR-182-5p,
hsa-miR-26a-5p, FRS2, CORO1C, FOXP1,
CPEB4 and GLCCI1 may contribute to the pathogenesis
of osteosarcoma.
The expression of hsa-miR-27a-3p serves an important
role in the development of a number of types of cancer, including
breast and pancreatic cancer. The expression of hsa-miR-27a-3p was
positively correlated with a hypoxia gene signature in breast
cancer (26). The inhibition of
hsa-miR-27a-3p expression may exhibit potentially antiproliferative
effects in pancreatic cancer (27).
In addition, hsa-miR-27a-3p was considered as a candidate biomarker
for Alzheimer's disease (28).
However, the correlation between hsa-miR-27a-3p and osteosarcoma
has not been widely investigated. In the present study,
hsa-miR-27a-3p was identified as a hub gene with a high degree of
association in miRNA-gene regulatory network. In combination with
previous studies, the present study hypothesized that
hsa-miR-27a-3p may serve a role in the development of
osteosarcoma.
Hsa-miR-9-5p, hsa-miR-155-5p and hsa-miR-203 are
potent prognostic factors for acute myeloid leukemia (29). In addition, a 3-miRNA scoring system
(hsa-miR-9-5p, hsa-miR-155-5p and hsa-miR-203) was used for the
prognostication of the patients with de novo acute myeloid
leukemia (30). Hsa-miR-182-5p has
been considered as a marker for distinguishing between human
ovarian cancer tissues and normal tissues (31). The knockdown of miR-182-5p
significantly decreased the growth of prostate tumor, and
FOXF2, reversion inducing cysteine rich protein with kazal
motifs (RECK) and metastasis suppressor 1 were identified as
potential target genes of miR-182-5p (32). Previous studies have also revealed
that mothers against decapentaplegic homolog 4 (Smad4) and
RECK were the potential target genes of miR-182-5p, and that
miR-182-5p served a key role in bladder cancer by knocking down the
expression of Smad4 and RECK (33). In the present study, hsa-miR-9-5p and
hsa-miR-182-5p were identified as the hub genes with high degrees
of association in the miRNA-gene regulatory network, implying that
hsa-miR-9-5p and hsa-miR-182-5p may serve a key role in the
pathogenesis of osteosarcoma.
In the miRNA-gene regulatory network, FRS2,
CORO1C, FOXP1, CPEB4 and GLCCI1 with
high degrees of association were identified, implying that these
genes may be associated with the regulatory mechanism of
osteosarcoma. FRS2 is considered to be a gene that is
associated with numerous types of cancer, including ovarian cancer,
liposarcoma and prostate cancer (34–36).
FRS2 serves as an amplified oncogene that induces the
downstream activation of the Ras-mitogen-activated protein kinase
pathway in high-grade serous ovarian cancer (36). In addition, FRS2 serves an
essential role in fibroblast growth factor receptor (FGFR)
signaling, and activated FGFR/FRS2 signaling may lead to the
development of high-grade liposarcoma (34). Concomitantly, a previous study also
identified that the functional overlap of FRS2 and
FRS3 may mediate mitogenic FGF signaling in prostate cancer
(37). From these data, the present
study hypothesized that FRS2 was involved in the regulatory
mechanism of osteosarcoma.
CORO1C is a target gene of miR-206 that has
been demonstrated to inhibit cell migration in triple-negative
breast cancer (38). CORO1C is
also considered as a target gene of miR-1/133a following
genome-wide gene expression and luciferase reporter assay analyses
(39). Furthermore miR-1 and miR-133a
inhibited the proliferation, migration, and invasion of
lung-squamous cell carcinoma cells (39). In addition, miR-1 and miR-133b
mediated cell proliferation, and cell cycle progression by
regulating the expression of hepatocyte growth factor receptor
protein in human osteosarcoma (40).
miR-133b and miR-206 expression were identified to be significantly
decreased and may be used as potential prognostic markers for
patients with osteosarcoma (41).
Decreased miR-206 expression is associated with the development of
osteosarcoma, and the transfection of miR-206 mimics promoted cell
apoptosis, and inhibited cell invasion and migration (42). CORO1C may serve as the target
gene of miR-1, miR-133b and miR-206, which have all been associated
with osteosarcoma development. Furthermore, CORO1C was
identified as a node gene with a high degree of association in the
miRNA-gene regulatory network. However, CORO1C was not
identified as a target gene of the miRNAs in the present study.
Together, the results indicate that CORO1C may serve a role
in osteosarcoma pathogenesis and development by regulating
miRNAs.
FOXP1 expression is associated with the
development of osteosarcoma (43,44).
CPEB4 serves a role in metastatic cancer and cancer
progression (45,46); however, whether CPEB4 is
involved in the pathogenesis of osteosarcoma remains unclear. In
the present study, FOXP1 and CPEB4 were also
identified as the node genes with high degrees of association in
the miRNA-gene regulatory network. Based on the results, it was
hypothesized that FOXP1 and CPEB4 may contribute to
osteosarcoma progression, and development. However, these results
require additional confirmation.
In the present study, hsa-miR-27a-3p, hsa-miR-9-5p,
hsa-miR-182-5p, FRS2, CORO1C, FOXP1 and
CPEB4 were identified as node genes with high degrees of
association in the miRNA-gene regulatory network, and may serve a
role in the pathogenesis and development of osteosarcoma.
References
|
1
|
Savage SA, Mirabello L, Wang Z,
Gastier-Foster JM, Gorlick R, Khanna C, Flanagan AM, Tirabosco R,
Andrulis IL, Wunder JS, et al: Genome-wide association study
identifies two susceptibility loci for osteosarcoma. Nat Genet.
45:799–803. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kansara M and Thomas DM: Molecular
pathogenesis of osteosarcoma. DNA Cell Biol. 26:1–18. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Both J, Krijgsman O, Bras J, Schaap GR,
Baas F, Ylstra B and Hulsebos TJ: Focal chromosomal copy number
aberrations identify CMTM8 and GPR177 as new candidate driver genes
in osteosarcoma. PLoS One. 9:e1158352014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Broadhead ML, Clark JC, Myers DE, Dass CR
and Choong PF: The molecular pathogenesis of osteosarcoma: A
review. Sarcoma. 2011:9592482011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zong C, Lu S, Chapman AR and Xie XS:
Genome-wide detection of single-nucleotide and copy-number
variations of a single human cell. Science. 338:1622–1626. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jin J, Cai L, Liu ZM and Zhou XS:
miRNA-218 inhibits osteosarcoma cell migration and invasion by
down-regulating of TIAM1, MMP2 and MMP9. Asian Pac J Cancer Prev.
14:3681–3684. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang H, Wang Z, Bao L, et al: The
significance of copy number variation in multiple osteosarcoma's
malignance grade, drug resistance and classification. Mar Sci Bull.
24:41–46. 2015.
|
|
8
|
Cui W, Cai Y, Wang W, Liu Z, Wei P, Bi R,
Chen W, Sun M and Zhou X: Frequent copy number variations of
PI3K/AKT pathway and aberrant protein expressions of PI3K subunits
are associated with inferior survival in diffuse large B cell
lymphoma. J Transl Med. 12:102014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Silva AG, Krepischi AC, Pearson PL,
Hainaut P, Rosenberg C and Achatz MI: The profile and contribution
of rare germline copy number variants to cancer risk in Li-Fraumeni
patients negative for TP53 mutations. Orphanet J Rare Dis.
9:632014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gokgoz N, Wunder JS and Andrulis IL:
Abstract 5075: Genome-wide analysis of DNA copy number variations
in osteosarcoma. Can Res. 72:50752012. View Article : Google Scholar
|
|
11
|
Conde L, Riby J, Zhang J, Bracci PM and
Skibola CF: Copy number variation analysis on a non-Hodgkin
lymphoma case-control study identifies an 11q25 duplication
associated with diffuse large B-cell lymphoma. PLoS One.
9:e1053822014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shimada M: MicroRNA-mediated regulation of
apoptosis in osteosarcoma. J Carcinog Mutagen. 2013.doi:
10.4172/2157-2518.S6-001. View Article : Google Scholar
|
|
13
|
Jones KB, Salah Z, Del Mare S, Galasso M,
Gaudio E, Nuovo GJ, Lovat F, LeBlanc K, Palatini J, Randall RL, et
al: miRNA signatures associate with pathogenesis and progression of
osteosarcoma. Cancer Res. 72:1865–1877. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Osaki M, Takeshita F, Sugimoto Y, Kosaka
N, Yamamoto Y, Yoshioka Y, Kobayashi E, Yamada T, Kawai A, Inoue T,
et al: MicroRNA-143 regulates human osteosarcoma metastasis by
regulating matrix metalloprotease-13 expression. Mol Ther.
19:1123–1130. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu M, Jin H, Xu CX, Sun B, Song ZG, Bi WZ
and Wang Y: miR-382 inhibits osteosarcoma metastasis and relapse by
targeting Y box-binding protein 1. Mol Ther. 23:89–98. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Z and Cai H, Lin L, Tang M and Cai H:
Upregulated expression of microRNA-214 is linked to tumor
progression and adverse prognosis in pediatric osteosarcoma.
Pediatr Blood Cancer. 61:206–210. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shimbo K, Miyaki S, Ishitobi H, Kato Y,
Kubo T, Shimose S and Ochi M: Exosome-formed synthetic microRNA-143
is transferred to osteosarcoma cells and inhibits their migration.
Biochem Biophys Res Commun. 445:381–387. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Salah Z, Arafeh R, Maximov V, Galasso M,
Khawaled S, Abou-Sharieha S, Volinia S, Jones KB, Croce CM and
Aqeilan RI: miR-27a and miR-27a* contribute to metastatic
properties of osteosarcoma cells. Oncotarget. 6:4920–4935. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang H, Yin Z, Ning K, Wang L, Guo R and
Ji Z: Prognostic value of microRNA-223/epithelial cell transforming
sequence 2 signaling in patients with osteosarcoma. Hum Pathol.
45:1430–1436. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang YZ, Zhang J, Shao HY, Chen JP and
Zhao HY: MicroRNA-191 promotes osteosarcoma cells proliferation by
targeting checkpoint kinase 2. Tumour Biol. 36:6095–6101. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fujiwara T, Katsuda T, Hagiwara K, Kosaka
N, Yoshioka Y, Takahashi RU, Takeshita F, Kubota D, Kondo T,
Ichikawa H, et al: Clinical relevance and therapeutic significance
of microRNA-133a expression profiles and functions in malignant
osteosarcoma-initiating cells. Stem Cells. 32:959–973. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Song QC, Shi ZB, Zhang YT, Ji L, Wang KZ,
Duan DP and Dang XQ: Downregulation of microRNA-26a is associated
with metastatic potential and the poor prognosis of osteosarcoma
patients. Oncol Rep. 31:1263–1270. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kobayashi E, Satow R, Ono M, Masuda M,
Honda K, Sakuma T, Kawai A, Morioka H, Toyama Y and Yamada T:
Microrna expression and functional profiles of osteosarcoma.
Oncology. 86:94–103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maire G, Martin JW, Yoshimoto M,
Chilton-MacNeill S, Zielenska M and Squire JA: Analysis of
miRNA-gene expression-genomic profiles reveals complex mechanisms
of microRNA deregulation in osteosarcoma. Cancer Genet.
204:138–146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kuijjer ML, Rydbeck H, Kresse SH, Buddingh
EP, Lid AB, Roelofs H, Bürger H, Myklebost O, Hogendoorn PC,
Meza-Zepeda LA and Cleton-Jansen AM: Identification of osteosarcoma
driver genes by integrative analysis of copy number and gene
expression data. Genes Chromosomes Cancer. 51:696–706. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Camps C, Saini HK, Mole DR, Choudhry H,
Reczko M, Guerra-Assunção JA, Tian YM, Buffa FM, Harris AL,
Hatzigeorgiou AG, et al: Integrated analysis of microRNA and mRNA
expression and association with HIF binding reveals the complexity
of microRNA expression regulation under hypoxia. Mol Cancer.
13:282014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu X, Dai C, Hou A, Cui J, Cheng D and Xu
D: Dysregulated microRNA Profile in HeLa Cell Lines Induced by
LupeolBioinformatics Research and Applications. Springer; pp.
71–80. 2014
|
|
28
|
Frigerio CS, Lau P, Salta E, Tournoy J,
Bossers K, Vandenberghe R, Wallin A, Bjerke M, Zetterberg H,
Blennow K and De Strooper B: Reduced expression of hsa-miR-27a-3p
in CSF of patients with Alzheimer disease. Neurology. 81:2103–2106.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chuang MK, Chiu YC, Chou WC, Hou HA,
Chuang EY and Tien HF: A simple, powerful, and widely applicable
Micro-RNA scoring system in prognostication of de novo myeloid
leukemia patients. Blood. 124:712014.
|
|
30
|
Chuang MK, Chiu YC, Chou WC, Hou HA,
Chuang EY and Tien HF: A 3-microRNA scoring system for
prognostication in de novo acute myeloid leukemia patients.
Leukemia. 29:1051–1059. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang L, Zhu MJ, Ren AM, Wu HF, Han WM, Tan
RY and Tu RQ: A ten-microRNA signature identified from a
genome-wide microRNA expression profiling in human epithelial
ovarian cancer. PLoS One. 9:e964722014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hirata H, Ueno K, Shahryari V, Deng G,
Tanaka Y, Tabatabai ZL, Hinoda Y and Dahiya R: MicroRNA-182-5p
promotes cell invasion and proliferation by down regulating FOXF2,
RECK and MTSS1 genes in human prostate cancer. PLoS One.
8:e555022013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hirata H, Ueno K, Shahryari V, Tanaka Y,
Tabatabai ZL, Hinoda Y and Dahiya R: Oncogenic miRNA-182-5p targets
Smad4 and RECK in human bladder cancer. PLoS One. 7:e510562012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang K, Chu K, Wu X, Gao H, Wang J, Yuan
YC, Loera S, Ho K, Wang Y, Chow W, et al: Amplification of FRS2 and
activation of FGFR/FRS2 signaling pathway in high-grade
liposarcoma. Cancer Res. 73:1298–1307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tania V, Ajay J, Naveen K, Steve D, Susan
M and Gnanapragasam VJ: Role and expression of FRS2 and FRS3 in
prostate cancer. BMC Cancer. 11:4842011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Luo LY, Kim E, Cheung HW, Weir BA, Dunn
GP, Shen RR and Hahn WC: The tyrosine kinase adaptor protein FRS2
is oncogenic and amplified in high-grade serous ovarian cancer. Mol
Cancer Res. 13:502–509. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Valencia T, Joseph A, Kachroo N, Darby S,
Meakin S and Gnanapragasam VJ: Role and expression of FRS2 and FRS3
in prostate cancer. BMC Cancer. 11:4842011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang J, Tsouko E, Jonsson P, Bergh J,
Hartman J, Aydogdu E and Williams C: Abstract P4-07-12: miR-206
inhibits cell migration through direct targeting of the
actin-binding protein coronin 1C in triple-negative breast cancer.
Cancer Res. 73 (24 Suppl):(P4): 07–12. 2013.
|
|
39
|
Mataki H, Enokida H, Chiyomaru T, Mizuno
K, Matsushita R, Goto Y, Nishikawa R, Higashimoto I, Samukawa T,
Nakagawa M, et al: Downregulation of the microRNA-1/133a cluster
enhances cancer cell migration and invasion in lung-squamous cell
carcinoma via regulation of Coronin1C. J Hum Genet. 60:53–61. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Novello C, Pazzaglia L, Cingolani C, Conti
A, Quattrini I, Manara MC, Tognon M, Picci P and Benassi MS: miRNA
expression profile in human osteosarcoma: Role of miR-1 and
miR-133b in proliferation and cell cycle control. Int J Oncol.
42:667–675. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang C, Yao C, Li H, Wang G and He X:
Serum levels of microRNA-133b and microRNA-206 expression predict
prognosis in patients with osteosarcoma. Int J Clin Exp Pathol.
7:4194–4203. 2014.PubMed/NCBI
|
|
42
|
Bao YP, Yi Y, Peng LL, Fang J, Liu KB, Li
WZ and Luo HS: Roles of microRNA-206 in osteosarcoma pathogenesis
and progression. Asian Pac J Cancer Prev. 14:3751–3755. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
van Boxtel R, Gomez-Puerto C, Mokry M,
Eijkelenboom A, van der Vos KE, Nieuwenhuis EE, Burgering BM, Lam
EW and Coffer PJ: FOXP1 acts through a negative feedback loop to
suppress FOXO-induced apoptosis. Cell Death Differ. 20:1219–1229.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Santo EE, Ebus ME, Koster J, Schulte JH,
Lakeman A, van Sluis P, Vermeulen J, Gisselsson D, Øra I, Lindner
S, et al: Oncogenic activation of FOXR1 by 11q23 intrachromosomal
deletion-fusions in neuroblastoma. Oncogene. 31:1571–1581. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xu H and Liu B: CPEB4 is a candidate
biomarker for defining metastatic cancers and directing
personalized therapies. Med Hypotheses. 81:875–877. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ortiz-Zapater E, Pineda D, Martínez-Bosch
N, Fernández-Miranda G, Iglesias M, Alameda F, Moreno M, Eliscovich
C, Eyras E, Real FX, et al: Key contribution of CPEB4-mediated
translational control to cancer progression. Nat Med. 18:83–90.
2011. View Article : Google Scholar : PubMed/NCBI
|