Introduction
Osteosarcoma is a type of malignant tumor, which
often occurs in children (1).
Surgical methods in the treatment of osteosarcoma frequently
involve wide resection or amputation, which is likely to result in
a disability or deformity, with the consequence of poor quality of
life (2). For this reason, early
warning, diagnosis and treatment are exceptionally important.
Previous studies revealed that microRNA (miRNA/miR) has a major
role in the occurrence and development of numerous diseases such as
ischemic cardiomyopathy, gastric cancer and hepatocellular
carcinoma (3,4). As a non-coding single-chain
micro-molecule RNA, miRNA is made up of 21–25 nucleotides (5,6). Through
incomplete complementary sequence combination with the 3′
untranslated region (3′UTR) sequence of target mRNA, miRNA is able
to inhibit mRNA translation or cause direct degradation to the
extent that the miRNA level shows the effect of gene silencing
following transcription (7). Studies
have revealed that multiple miRNAs are able to serve as oncogenes
or anti-oncogenes in various tumors, including osteosarcoma, while
specific miRNA is involved in each stage of the occurrence and
development of tumors (8,9).
Previous studies indicated that miR-144 is
downregulated expression in colon cancer miR-144 is in close
association with the development of colorectal cancer and can be
used as a molecular diagnostic marker of colon cancer (10,11). A
previous study demonstrated that downregulation of miR-144 is able
to result in the proliferation of bladder cancer cells via EZH2 and
Wnt signaling pathways (12). Another
study investigating the function of miR-144 revealed that GATA4 may
activate miR-144 precursors and induce their expression (13), and studies have also suggested that
the overexpression of miR-144 may cause apoptosis of myocardial
cells and inhibit their proliferation (14,15).
Furthermore, miR-144 is considered to be a potential target for the
treatment of ischemic heart disease (13,15).
However, whether miR-144 can affect the biological function of
osteosarcoma cells is yet to be reported, and its mechanism remains
unknown. Therefore, the current study focused on investigating the
effect of miR-144 on the proliferation and apoptosis of
osteosarcoma cell lines (MG-63 and U-2) and investigating its
mechanism of action.
Materials and methods
Cell lines and cell culture
The human osteosarcoma cell lines, MG-63 and U-2 OS,
were obtained from the American Type Culture Collection (Manassas,
VA, USA). MG-63 was maintained in minimum essential medium
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% fetal bovine serum (FBS; Invitrogen; Thermo Fisher
Scientific, Inc.), and U-2 OS was cultured in RPMI-1640
(Invitrogen; Thermo Fisher Scientific, Inc.) containing 10% horse
serum (Invitrogen; Thermo Fisher Scientific, Inc.) and 5% FBS. The
two mediums were supplemented with 100 U/ml penicillin and 100 U/ml
streptomycin sulfate (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany), and the cells were cultured in a humidified 5%
CO2 incubator at 37°C.
Evaluation of miR-144 expression in
osteosarcoma cells
For RNA analysis, each cell line was seeded at
1×105 cells/well in a volume of 2 ml on 6-well
flat-bottomed plates. Total RNA from these cell lines was isolated
using the mirVana miRNA Isolation kit (Ambion; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
protocol following 48 h incubation at 37°C. Then, cDNA was
synthesized from 10 ng of total RNA using TaqMan MicroRNA
hsa-miR-144 specific primers (Applied Biosystems; Thermo Fisher
Scientific, Inc.) and a TaqMan MicroRNA Reverse Transcription kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.). Reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) was
performed to measure miR-144 expression, and the primers were as
follows: miR-144 forward, 5′GCT GGG ATA TCA TCA TAT ACT G3′ and
reverse, 5′CGG ACT AGT ACA TCA TCT ATA CTG3′; and U6 forward, 5′CTC
GCT TCG GCA GCA CA3′ and reverse, 5′AAC GCT TCA CGA ATT TGC GT3′.
The following temperature profile was used: Initial denaturation at
95°C for 10 min, followed by 45 cycles of denaturation at 95°C for
10 sec, annealing at 60°C for 10 sec and extension at 65°C for 10
sec. PCR was performed in a LightCyclerTM 480 system (Roche Applied
Science, Penzburg, Germany) using the LightCyclerTM 480 Probes
Master kit (Roche Applied Science). Expression levels of miR-144
were normalized to that of the small nuclear RNA U6 (Applied
Biosystems; Thermo Fisher Scientific, Inc.) transcript. Relative
quantification of miRNA expression was calculated using the
2−ΔΔCq method (16).
Transfection of miR-144 mimics and
transfection efficiency evaluation
Designed and provided by RiboBio (Guangzhou RiboBio
Co., Ltd., Guangzhou, China), human miR-144 duplex mimics and
scramble control (with the final concentration of 10 nM) were
transfected into cells using Lipofectamine™ 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. At 24 h post-transfection at 37°C, cells
transfected with miRNA mimics or scramble were used for subsequent
experiments, including proliferation and apoptosis assays.
Cell proliferation and viability
assays
MG-63 or U-2 OS cells in the logarithmic phase were
transfected with miR-144 mimics or scramble control and seeded at
4.0×103 cells/well in 96-well flat-bottomed microtiter
plates in a final volume of 100 µl culture medium per well. The
cells were incubated in a humidified atmosphere at 37°C and 5%
CO2 for 24, 48, 72 and 96 h. MTT assays were used to
measure cell viability during each time period. Dimethyl sulfoxide
was added into each well to dissolve the purple formazan.
Subsequent to the formazan crystals being dissolved, the absorbance
was determined spectrophotometrically at 570 nm using a reference
wavelength of 630 nm on an ELX800 UV universal microplate reader
(Bio-Tek Instruments Inc., Winooski, VT, USA).
Proliferation and apoptosis
assays
Proliferation status of the cells was determined by
measuring the incorporation of 5-bromo-2′-deoxyuridine (BrdU). The
cells were incubated with 100 µM BrdU (Sigma-Aldrich; Merck KGaA)
for 1–2 h at 37°C, and BrdU labeling was detected by confocal laser
scanning microscopy or flow cytometry following incubation for 30
min at 37°C with a fluorescein isothiocyanate- or
allophycocyanin-conjugated anti-BrdU antibody (cat. no. 558599;
dilution, 1:200; BD Biosciences, Franklin Lakes, NJ, USA). The
images were captured on a BD Pathway™ 435 confocal
imager with an objective magnification, ×20, and merged using BD
AttoVision™ software v1.6 (BD Biosciences). Staining of
samples without the addition of BrdU was used as a negative
control. The cells were subsequently analyzed and quantified by
flow cytometry. In brief, the cells were stained for 30 min at 37°C
with the following antibody: Anti-BrdU (cat. no. 558599; dilution,
1:200; BD Biosciences), then the labeled cells were thoroughly
washed with 2X PBS and analyzed on BD FACSCantoII flow cytometer
(BD Biosciences). Data were analyzed by BD FACSDiva Software v6.0
(BD Biosciences).
Dexamethasone was used to induce cell
apoptosis
Duration the culturing of cells, dexamethasone with
a concentration of 0.1 µM was added into culture medium. In
situ Cell Death Detection kit (Roche Diagnostics GmbH,
Mannheim, Germany) was used for TUNEL staining to determine the
apoptotic status of the cells, according to the manufacturer's
protocol. Zeiss Axio Imager 2 fluorescence microscope (Carl Zeiss
AG, Oberkochen, Germany) was used to observe and capture images at
a magnification, ×100. A total of 3 fields of view in each group
were obtained.
Protein expression analysis
Western blot analysis was used to confirm mammalian
target of rapamycin (mTOR) or proteins associated with
proliferation or apoptosis expression in miR-144-mimic-transfected
cells and control cells. In brief, cell proteins were collected
with radioimmunoprecipitation assay lysis buffer (Auragene
Bioscience, Changsha, China) to obtain cellular protein and the
protein concentrations were measured using bicinchoninic acid
protein assay kit (Pierce; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. A total of 30 µg of
cellular proteins were separated on 10 or 12% SDS-PAGE gels, and
were subsequently transferred onto polyvinylidene fluoride (PVDF)
membranes (EMD Millipore, Billerica, MA, USA). Then, the membranes
were blocked in 5% milk for 1 h at room temperature and incubated
with primary antibodies overnight at 4°C, and subsequently
incubated with horseradish peroxidase-conjugated anti-rabbit IgG
(cat. no. 7074; dilution, 1:2,000; Cell Signaling Technology, Inc.,
Danvers, MA, USA) at room temperature for 1 h. The following
primary antibodies and dilutions were used: mTOR rabbit monoclonal
antibody (cat. no. 2983; dilution, 1:1,000; Cell Signaling
Technology, Inc.) and GAPDH rabbit monoclonal antibody (cat. no.
5014; dilution, 1:1,000; Cell Signaling Technology, Inc.), p53
rabbit monoclonal antibody (cat. no. 2527; dilution, 1:1,000; Cell
Signaling Technology, Inc.), PCNA rabbit monoclonal antibody (cat.
no. 13110; dilution, 1:1,000; Cell Signaling Technology, Inc.),
Bcl-2 rabbit monoclonal antibody (cat. no. 3498; dilution, 1:1,000;
Cell Signaling Technology, Inc.), Bcl-xL rabbit monoclonal antibody
(cat. no. 2764; dilution, 1:1,000; Cell Signaling Technology,
Inc.), Bax rabbit monoclonal antibody (cat. no. 5023; dilution,
1:1,000; Cell Signaling Technology, Inc.), caspase 3 rabbit
monoclonal antibody (cat. no. 14220; dilution, 1:1,000; Cell
Signaling Technology, Inc.). Signals for each protein expression
were detected with electrochemiluminescence reagents (GE Healthcare
Life Sciences, Little Chalfont, UK) according to the manufacturer's
protocol.
Target genes of miR-144
TargetScanHuman 7.0 (http://www.targetscan.org/) and miRBase (http://www.mirbase.org/) microRNA databases were used
to identify the target genes of miR-144.
Construction of reporter plasmids and
the luciferase reporter assay
To construct a luciferase reporter plasmid, a
full-length fragment of the mTOR 3′UTR was subcloned into pmirGLO
dual-luciferase miRNA target expression vector (Promega
Corporation, Madison, WI, USA) located 5′ to the firefly
luciferase. The nucleotide sequences of the constructed plasmids
were confirmed by DNA sequencing. For luciferase reporter assays,
MG-63 cells (5×104/well) were seeded into a 96-well
plate and then co-transfected via Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) with the primer
GLO-mTOR 3′UTR or Rictor 3′UTR (Promega Corporation) construct and
miR-144-mimics or scramble control (Ambion; Thermo Fisher
Scientific, Inc.). Assays were performed 48 h after transfection
using the Dual-Luciferase Reporter Assay system (Promega
Corporation) according to the manufacturer's protocol. The firefly
luciferase signals were normalized to the Renilla luciferase
signals. Transfections were repeated three times in independent
experiments.
Statistical analysis
Quantitative data are expressed as the mean ±
standard error. Statistical analysis was performed by using one-way
analysis of variance followed by the least significant difference
test. Statistical analysis was performed with GraphPad Prism 7
software (GraphPad Software, Inc., La Jolla, CA, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
miR-144 expression is decreased in
osteosarcoma cells
To investigate whether miR-144 regulates
osteosarcoma progression, the expression level of miR-144 was
examined in osteosarcoma cell lines (MG-63 and U-2 OS). Compared
with the normal osteoblast cells, miR-144 expression levels were
significantly reduced in osteosarcoma cell lines (Fig. 1A). To investigate the biological role
of miR-144 in regulating osteosarcoma cells, miR-144 mimics or
scramble control were transfected into MG-63 or U-2 OS cells. As
shown in Fig. 1B and C, the
expression level of miR-144 was significantly increased in
miR-144-mimic-transfected cells compared with control cells or
scramble-transfected cells.
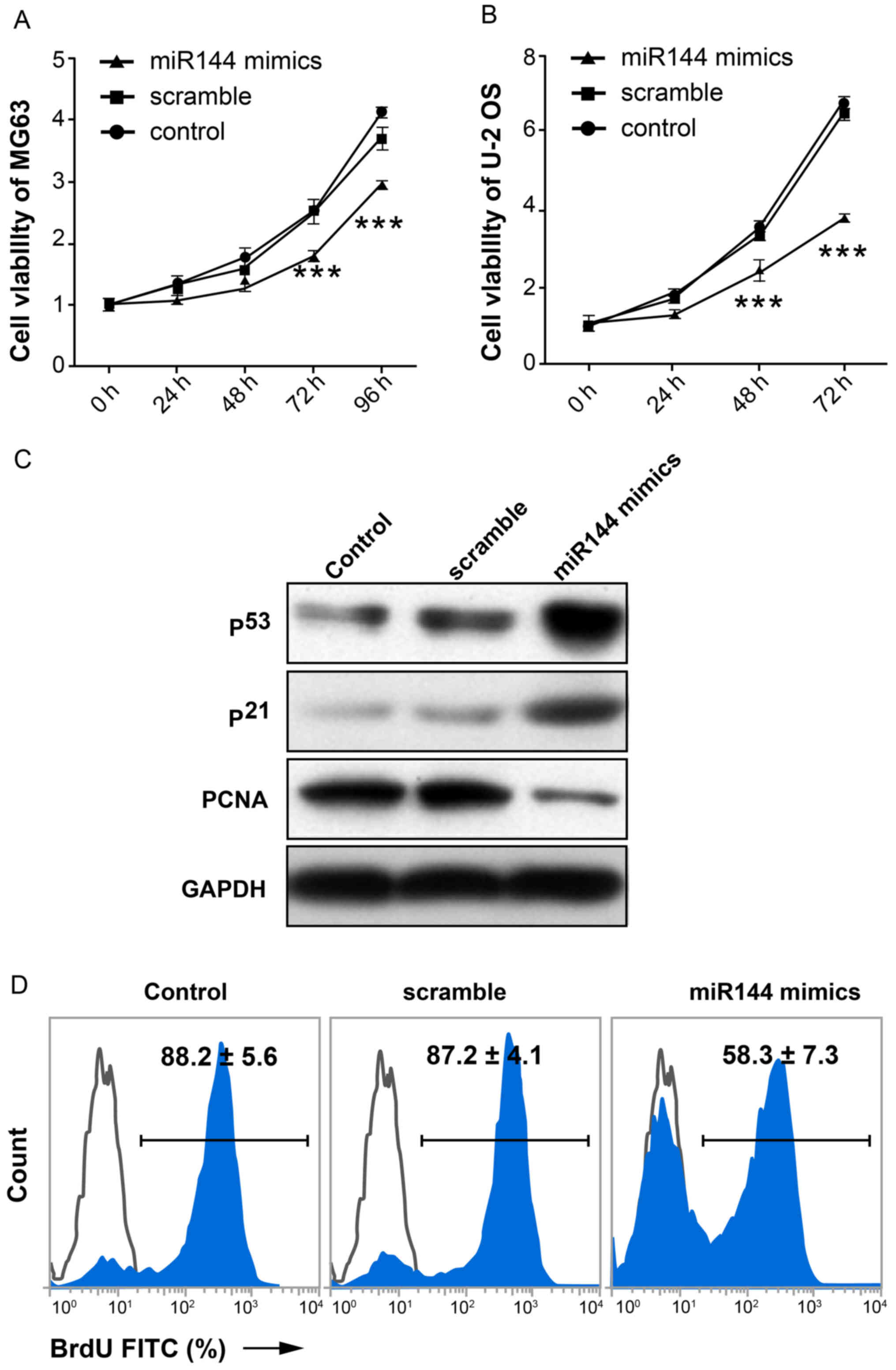
miR-144 inhibits proliferation of
osteosarcoma cells
To investigate the possible effect of miR-144 on
proliferation of osteosarcoma cells, MTT assay was performed. The
results demonstrated that the proliferation capacity of cells in
the miR-144-mimic-transfected group exhibited a significant decline
compared with the scramble and control groups (P<0.01; n=3;
Fig. 2A and B). The expression level
of proteins associated with proliferation (p53, p21 and PCNA) in
MG-63 cells is shown in Fig. 2C. p53
and p21 act as cell cycle arrest genes (17). PCNA is a biomarker of cell
proliferation (18–20). The expression level of p53 and p21 in
the miR-144-mimic-transfected group was markedly increased compared
with the scramble and control groups. By contrast, the expression
of PCNA was markedly decreased in the miR-144-mimic-transfected
group compared with the other two groups. BrdU-incorporated flow
cytometry showed similar data (58.3±7.3 vs. 87.2±4.1,
miR-144-mimic-transfected group and scramble control, respectively;
Fig. 2D). These data demonstrated
that miR-144 inhibited the proliferation of osteosarcoma cells.
Overexpression of miR-144 promotes
apoptosis of osteosarcoma cells induced by dexamethasone
To determine the effect of miR-144 on the apoptosis
of osteosarcoma cells, dexamethasone was added to the culture
medium to induce apoptosis. TUNEL staining images showed a greater
number of TUNEL+ cells in the miR-144-mimic-transfected
group compared with the control or scramble-transfected groups
(Fig. 3A). The expression level of
proteins associated with apoptosis signaling pathways was examined
by western blot analysis as shown in Fig.
3B. The data showed that two anti-apoptotic proteins, Bcl-2 and
Bcl-xL were markedly downregulated in the miR-144-mimic group
compared with the control and scramble groups. However, two
pro-apoptotic proteins, Bax and caspase 3, were markedly
upregulated in the miR-144-mimic-transfected group. These data
showed that the overexpression of miR-144 may promote apoptosis of
osteosarcoma cells induced by dexamethasone.
miR-144 directly targets mTOR and
inhibits its expression
In order to identify the target genes of miR-144,
candidate genes were identified using TargetScanHuman 7.0
(http://www.targetscan.org/) and miRBase
(http://www.mirbase.org/) micoRNA databases.
miR-144 targets the mTOR gene, an important molecule involved in
tumorigenesis and cancer progression (21,22)
(Fig. 4A). To confirm this
prediction, a luciferase reporter vector containing the 3′UTR of
mTOR [pGL3-mTOR-3′UTR-WT (wild-type)] was constructed. The reporter
assay showed that miR-144 was able to significantly suppress
luciferase expression (Fig. 4B).
Additionally, miR-144 reduced the levels of mTOR protein in MG-63
cells (Fig. 4C). As expected, mTOR
levels were markedly upregulated in MG-63 or U-2 OS cells compared
with normal osteoblast cells (Fig.
4D). These data demonstrated that miR-144 inhibits endogenous
mTOR expression in osteosarcoma cells.
Discussion
MiRNAs primarily function at the post-transcription
level, and they may induce downregulation of gene expression
through sequence complementarity between the miRNAs and their
targeted mRNA (7). miRNA sequences
have properties of high conservation, tissue specificity and time
sequence in different species (23,24). As
described in the literature, miRNA performs an important role in
the occurrence, development and metastasis of tumors (25,26).
Furthermore, the abnormal expression of miRNA is associated with
the occurrence of certain tumors such as gastric and breast cancer
(27,28). Results of the present study revealed
that miR-144 can directly target mTOR and inhibit the signaling
pathway of phosphoinositide 3-kinase (PI3K)/Akt, so as to inhibit
the proliferation of osteosarcoma cells and promote their
apoptosis.
Significant changes in miR-144 expression have been
observed in a variety of diseases, including colorectal and bladder
cancer (10,12). However, the majority of studies have
focused on the blood system, while fewer studies have investigated
tumors. Through meta-analysis, Wang et al (29) found the presence of
differentially-expressed miRNAs in osteosarcoma tissues. In
particular, the differential expression of miR-144 was revealed in
colorectal cancer, pancreatic cancer, thyroid cancer, laryngeal
cancer and malignant mesothelioma tissues. Additionally, miR-144
was demonstrated to be downregulated in laryngeal cancer (30), cervical cancer (31), follicular thyroid carcinoma (32) and malignant mesothelioma (33). Therefore, the present study
hypothesized that transfecting cells with an externally constructed
vector of miRNA-144 may cause cell growth retardation and death,
primarily apoptosis.
Results of the present study are consistent with the
results of previous studies on liver cancer and non-small cell lung
cancer (34,35). It is generally considered that miR-144
is able to function as a suppressor gene of tumor proliferation and
affect the prognosis of patients with osteosarcoma (36,37).
Studies have reported that subsequent to transfecting liver cancer
HepG2 cells, the plasmids of overexpressed miR-144 are able to
promote apoptosis and inhibit the occurrence of liver cancer
metastasis by affecting the expression of E2F3 (34,38).
Results of the present study demonstrated that overexpression of
miR-144 in osteosarcoma cells is able to markedly inhibit
proliferation and strengthen the apoptosis-promoting effect of
dexamethasone on osteosarcoma cells. Therefore, this indicates that
miR-144 may also serve as a tumor suppressor gene in osteosarcoma
cells.
mTOR is an evolutionarily conserved protein kinase
(39). As aberrant signaling of the
mTOR pathway is associated with various malignancies, mTOR protein
is considered to be a key regulatory factor for cell growth, and
has gradually become a novel target for tumor therapy (40). mTOR may exert its function through
participation in the PI3K/Akt/mTOR pathway, and the regulatory
effect of the deregulation of the mTOR signaling pathway on tumors
is primarily via inhibiting apoptosis, promoting cell survival,
accelerating cell cycle transmission, enhancing angiogenesis and
promoting the invasion and metastasis of tumor cells (41). An in vitro study on non-small
lung cancer cell lines indicated that human papilloma virus 16 may
activate the PI3K/Akt/mTOR signaling pathways to induce the
expression of angiogenic cytokines, including hypoxia-inducible
factor-1α, vascular endothelial growth factor and interleukin-8
(42). Zhang et al (43) used mTOR-targeted inhibition combined
with antiandrogen for the treatment of prostate cancer and
demonstrated that the antitumor effect was greater than the single
drug, which indicated that inhibiting mTOR was able to increase the
effectiveness of anticancer drugs. Iwaya et al (10) reported that downregulating miR-144 in
colon cancer cells may activate mTOR signaling pathways and
further, promote the proliferation of colon cancer cells and
increase the sensitivity of colon cancer cells to rapamycin.
Results of the present study identified that upregulating the
miR-144 in osteosarcoma cells was able to downregulate the activity
of mTOR signaling pathways and inhibit the proliferation of
osteosarcoma cells, thereby promoting their sensitivity to
dexamethasone-induced apoptosis. These results are consistent with
the findings of Iwaya et al (10).
In conclusion, miR-144 may effectively inhibit the
proliferation and apoptosis of osteosarcoma cells by directly
targeting the mRNA 3′UTR region of mTOR genes and inhibiting the
synthesis of mTOR proteins, thereby inhibiting PI3K/Akt signaling
pathways. As studies on miR-144 continue, miR-144 may become a
novel target for the treatment of osteosarcoma.
Acknowledgements
The authors would like to acknowledge Technical
Support from the First Affiliated Hospital, China Medical
University (Shenyang, China).
References
|
1
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ruggieri P, Mavrogenis AF and Mercuri M:
Quality of life following limb-salvage surgery for bone sarcomas.
Expert Rev Pharmacoecon Outcomes Res. 11:59–73. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Latronico MV, Catalucci D and Condorelli
G: MicroRNA and cardiac pathologies. Physiol Genomics. 34:239–242.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li Y and Kowdley KV: MicroRNAs in common
human diseases. Genomics Proteomics Bioinformatics. 10:246–253.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim VN: MicroRNA biogenesis: Coordinated
cropping and dicing. Nat Rev Mol Cell Biol. 6:376–385. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Seitz H, Youngson N, Lin SP, Dalbert S,
Paulsen M, Bachellerie JP, Ferguson-Smith AC and Cavaillé J:
Imprinted microRNA genes transcribed antisense to a reciprocally
imprinted retrotransposon-like gene. Nat Genet. 34:261–262. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ying SY, Chang DC and Lin SL: The microRNA
(miRNA): Overview of the RNA genes that modulate gene function. Mol
Biotechnol. 38:257–268. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:pp. 2257–2261.
2006; View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Iwaya T, Yokobori T, Nishida N, Kogo R,
Sudo T, Tanaka F, Shibata K, Sawada G, Takahashi Y, Ishibashi M, et
al: Downregulation of miR-144 is associated with colorectal cancer
progression via activation of mTOR signaling pathway.
Carcinogenesis. 33:2391–2397. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kalimutho M, Del Vecchio Blanco G, Di
Cecilia S, Sileri P, Cretella M, Pallone F, Federici G and
Bernardini S: Differential expression of miR-144* as a novel
fecal-based diagnostic marker for colorectal cancer. J
Gastroenterol. 46:1391–1402. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo Y, Ying L, Tian Y, Yang P, Zhu Y, Wang
Z, Qiu F and Lin J: miR-144 downregulation increases bladder cancer
cell proliferation by targeting EZH2 and regulating Wnt signaling.
FEBS J. 280:4531–4538. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang X, Wang X, Zhu H, Zhu C, Wang Y, Pu
WT, Jegga AG and Fan GC: Synergistic effects of the GATA-4-mediated
miR-144/451 cluster in protection against simulated
ischemia/reperfusion-induced cardiomyocyte death. J Mol Cell
Cardiol. 49:841–850. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang F, Huang XY, Yan DS, Zhou X and Yang
DY: MicroRNA-144 over-expression induced myocytes apoptosis.
Zhonghua Xin Xue Guan Bing Za Zhi. 39:353–357. 2011.(In Chinese).
PubMed/NCBI
|
|
15
|
Yu M, Liu Y, Zhang B, Shi Y, Cui L and
Zhao X: Inhibiting microRNA-144 abates oxidative stress and reduces
apoptosis in hearts of streptozotocin-induced diabetic mice.
Cardiovasc Pathol. 24:375–381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pietenpol JA and Stewart ZA: Cell cycle
checkpoint signaling: Cell cycle arrest versus apoptosis.
Toxicology. 181–182. 1–481. 2002.PubMed/NCBI
|
|
18
|
Bologna-Molina R, Mosqueda-Taylor A,
Molina-Frechero N, Mori-Estevez AD and Sánchez-Acuña G: Comparison
of the value of PCNA and Ki-67 as markers of cell proliferation in
ameloblastic tumors. Med Oral Patol Oral Cir Bucal. 18:e174–e179.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Juríková M, Danihel Ľ, Polák Š and Varga
I: Ki67, PCNA, and MCM proteins: Markers of proliferation in the
diagnosis of breast cancer. Acta Histochem. 118:544–552. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Castilla C, McDonough P, Tumer G, Lambert
PC and Lambert WC: Sometimes it takes darkness to see the light:
Pitfalls in the interpretation of cell proliferation markers (Ki-67
and PCNA). Skinmed. 10:90–92. 2012.PubMed/NCBI
|
|
21
|
Geissler EK, Schlitt HJ and Thomas G:
mTOR, cancer and transplantation. Am J Transplant. 8:2212–2218.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sabatini DM: mTOR and cancer: Insights
into a complex relationship. Nat Rev Cancer. 6:729–734. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Altuvia Y, Landgraf P, Lithwick G, Elefant
N, Pfeffer S, Aravin A, Brownstein MJ, Tuschl T and Margalit H:
Clustering and conservation patterns of human microRNAs. Nucleic
Acids Res. 33:2697–2706. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen K and Rajewsky N: Deep conservation
of microRNA-target relationships and 3′UTR motifs in vertebrates,
flies, and nematodes. Cold Spring Harb Symp Quant Biol. 71:149–156.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Di Leva G, Garofalo M and Croce CM:
MicroRNAs in cancer. Annu Rev Pathol. 9:287–314. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Acunzo M, Romano G, Wernicke D and Croce
CM: MicroRNA and cancer-a brief overview. Adv Biol Regul. 57:1–9.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tong F, Cao P, Yin Y, Xia S, Lai R and Liu
S: MicroRNAs in gastric cancer: From benchtop to bedside. Dig Dis
Sci. 59:24–30. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Serpico D, Molino L and Di Cosimo S:
microRNAs in breast cancer development and treatment. Cancer Treat
Rev. 40:595–604. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang W, Peng B, Wang D, Ma X, Jiang D,
Zhao J and Yu L: Human tumor microRNA signatures derived from
large-scale oligonucleotide microarray datasets. Int J Cancer.
129:1624–1634. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang P, Fu T, Wang X and Zhu W: Primary,
study of miRNA expression patterns in laryngeal carcinoma by
microarray. Lin Chung Er Bi Yan Hou Tou Jing Wai Ke Za Zhi.
24:535–538. 2010.(In Chinese). PubMed/NCBI
|
|
31
|
Tian Y, Tian Y, Luo X, Zhou T, Huang Z,
Liu Y, Qiu Y, Hou B, Sun D, Deng H, et al: Identification and
characterization of microRNAs related to salt stress in broccoli,
using high-throughput sequencing and bioinformatics analysis. BMC
Plant Biol. 14:2262014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rossing M, Borup R, Henao R, Winther O,
Vikesaa J, Niazi O, Godballe C, Krogdahl A, Glud M, Hjort-Sørensen
C, et al: Down-regulation of microRNAs controlling tumourigenic
factors in follicular thyroid carcinoma. J Mol Endocrinol.
48:11–23. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Guled M, Lahti L, Lindholm PM, Salmenkivi
K, Bagwan I, Nicholson AG and Knuutila S: CDKN2A, NF2, and JUN are
dysregulated among other genes by miRNAs in malignant
mesothelioma-A miRNA microarray analysis. Genes Chromosomes Cancer.
48:615–623. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cao T, Li H, Hu Y, Ma D and Cai X: miR-144
suppresses the proliferation and metastasis of hepatocellular
carcinoma by targeting E2F3. Tumour Biol. 35:10759–10764. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zha W, Cao L, Shen Y and Huang M: Roles of
Mir-144-ZFX pathway in growth regulation of non-small-cell lung
cancer. PLoS One. 8:e741752013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao M, Huang J, Gui K, Xiong M, Cai G, Xu
J, Wang K, Liu D, Zhang X and Yin W: The downregulation of miR-144
is associated with the growth and invasion of osteosarcoma cells
through the regulation of TAGLN expression. Int J Mol Med.
34:1565–1572. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang W, Zhou X and Wei M: MicroRNA-144
suppresses osteosarcoma growth and metastasis by targeting ROCK1
and ROCK2. Oncotarget. 6:10297–10308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xue J, Niu YF, Huang J, Peng G, Wang LX,
Yang YH and Li YQ: miR-141 suppresses the growth and metastasis of
HCC cells by targeting E2F3. Tumour Biol. 35:12103–12107. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang H, Rudge DG, Koos JD, Vaidialingam B,
Yang HJ and Pavletich NP: mTOR kinase structure, mechanism and
regulation. Nature. 497:217–223. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pene F, Claessens YE, Muller O, Viguié F,
Mayeux P, Dreyfus F, Lacombe C and Bouscary D: Role of the
phosphatidylinositol 3-kinase/Akt and mTOR/P70S6-kinase pathways in
the proliferation and apoptosis in multiple myeloma. Oncogene.
21:6587–6597. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Foster KG and Fingar DC: Mammalian target
of rapamycin (mTOR): Conducting the cellular signaling symphony. J
Biol Chem. 285:14071–14077. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang E, Feng X, Liu F, Zhang P, Liang J
and Tang X: Roles of PI3K/Akt and c-Jun signaling pathways in human
papillomavirus type 16 oncoprotein-induced HIF-1α, VEGF, and IL-8
expression and in vitro angiogenesis in non-small cell lung cancer
cells. PLoS One. 9:e1034402014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang W, Zhu J, Efferson CL, Ware C,
Tammam J, Angagaw M, Laskey J, Bettano KA, Kasibhatla S, Reilly JF,
et al: Inhibition of tumor growth progression by antiandrogens and
mTOR inhibitor in a Pten-deficient mouse model of prostate cancer.
Cancer Res. 69:7466–7472. 2009. View Article : Google Scholar : PubMed/NCBI
|