Introduction
Lung cancer accounts for the majority of all
cancer-related deaths worldwide (1).
Non-small cell cancer (NSCLC) accounts for about 80-85 % of all
types of lung cancer, and the survival rate is very low (2). Since almost all patients have relapses,
it leads to the metastasis of disease and the death of patients.
Therefore it is urgent to search for better ideas to treat human
NSCLC.
MicroRNAs (miRNAs), an endogenous non-coding RNA,
were found in eukaryotic organisms, with a length of about 20~25
nucleotides. The abnormal expression of miRNAs was closely
correlated with the tumor development, including regulating cell
proliferation, migration, invasion and apoptosis (3–5). In recent
years, microarray analysis or other advanced technologies have
detected many miRNAs (6,7), and increasing research has shown that
miRNAs forge closer relationships with various cancers, including
gastric cancer, osteosarcoma and colorectal cancer (8–10).
MiR-1256 is located on chromosome 12 (11), a previous report has shown that
miR-1256 expression was downregulated and it inhibited
proliferation of prostate cancer and nasopharyngeal carcinoma
(12). However, the study of miR-1256
in NSCLC cells has not been reported.
It has been shown that miRNAs can inhibit gene
expression by degrading target messenger RNAs or blocking its
translation by combining its 3′-untranslated region (3-UTR),
thereby participating in the development of cancers (13,14). For
example, miR-1256 mimic inhibited prostate cancer cell growth and
invasion by regulating the PGK-1 (12). It has been proven that tectonic family
member 1 (TCTN1) played an important role in cellular
differentiation, apoptosis, and disease processes (15). Moreover, research has shown that TCTN1
regulated malignant cells growth of gastric cancer, prostate
cancer, colorectal cancer and glioblastoma (16–19). It
provides a novel therapeutic strategy for the treatment of cancers.
However, the function of TCTN1 and how miR-1256 regulated tumor
development in human NSCLC via targeting of TCTN1 has not been
reported.
In the present study, we investigated miR-1256 and
TCTN1 expression level in human NSCLC. The results showed that
miR-1256 was downregulated, whereas, TCTN1 was upregulated in
NSCLC, TCTN1 is characterised as a direct target of miR-1256 in
NSCLC cells. Our results provided a new potential direction for
NSCLC diagnosis and therapeutic interventions.
Materials and methods
Tissue collection
Normal tissues and normal lung cancer tissues were
acquired from 30 paired patients after they signed informed
consent. They had undergone surgery at People's Hospital of Rizhao
(Rizhao, China) between May, 2011 and April, 2016. Then, we stored
the samples at −80°C. All the experiments were approved by the
Ethics Committee of People's Hospital of Rizhao (Rizhao,
China).
Cell culture and transfection
The NSCLC cell lines (A549 and SK-MES-1) were
obtained from the Cell Resource Center, Institute of Biochemistry
and Cell Biology at the Chinese Academy of Sciences (Shanghai,
China). The two cell lines were cultured in RPMI-1640 (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), supplement with
10% fetal bovine serum (FBS), penicillin (100 U/ml) and
streptomycin (100 U/ml), which was incubated at 37°C under 5%
CO2 atmosphere. The cells were collected in logarithmic
phase for transfection. They were transfected using DharmaFect
Transfection Reagent (GE Healthcare Dharmacon, Inc., Lafayette, CO,
USA) according to the manufacturer's instructions.
Western blot analysis
After transfection for 48 h, RIPA lysis containing
proteinase inhibitors (Beyotime Institute of Biotechnology, Haimen,
China) and phenylmethanesulfonyl fluoride were used to extract
total protein from the A549 and SK-MES-1 cells. The protein
concentrations were tested with the BCA protein assay kit (Beyotime
Institute of Biotechnology). A 5X loading buffer was diluted to 1X
by adding into the supernatants containing total protein,
subsequently, the samples were heated at 95°C for 5 min, then
cooled to room temperature and stored in a refrigerator until use.
The total protein (50 µg) was added into the hole of SDS-PAGE and
performed electrophoresis at 60V voltage until the bromophenol blue
runs out of the bottom. The proteins were then transferred to
nitrocellulose filter membranes, then, skim milk (5-10%) was used
to block the proteins on the membranes at room temperature for 2 h.
Firstly, the membranes were incubated with the primary antibody
rabbit polyclonal to TCTN1 (cat. no. ab105381; 1:1,000; Abcam,
Cambridge, MA, USA) at 4°C overnight, after washed with 1×TBST (pH
7.4) three times later; the secondary antibodies goat anti-rabbit
IgG-HRP (cat. no. sc-2004; 1:3,000; Santa Cruz Biotechnology, Inc.
Santa Cruz, CA, USA) were added and incubated at room temperature
for 2 h. GAPDH primary antibody (cat. no. 70699; 1:5,000; Abcam,
Cambridge, MA, USA) was chosen as the internal reference. Protein
bands were detected using chemiluminescence method (ECL; EMD
Millipore, Billerica, MA, USA). GADPH served as a loading
control.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
We used TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) to extract total RNA from the
cell lines or tissue samples. For the detection of miR-1256, U6 was
used as an internal control, and cDNA was synthesized using a
TaqMan microRNA reverse transcription kit (Applied Biosystems;
Thermo Fisher Scientific, Inc.), and qPCR was performed using the
TaqMan microRNA PCR kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.). For the analysis of TCTN1, GAPDH was used as an
internal control, reverse transcription was conducted using the
moloney murine leukemia virus reverse transcription system (Promega
Corporation, Madison, WI, USA), followed by qPCR using the SYBR
Premix Ex Taq kit (Takara Biotechnology Co., Ltd., Dalian, China).
The sequences of the primers were as follows: miR-1256
GGCGCGATTTTAGTTTATC (forward) and TTTAATTACCAACCGAATACG (reverse);
for TCTN1: CCTTT GCGTGAATGTTGTTC (forward) and AGAGGGACTGGC
TGGGTATT (reverse); for U6 CTCGCTTCGGCAGCACA (forward) and
AACGCTTCACGAATTTGCGT (reverse); for GADPH: ACAACTTTGGTATCGTGGAAGG
(forward) and: GCCATCACGCCACAGTTTC (reverse). GADPH and U6 were
used as internal reference. The relative expression level was
calculated using the 2−ΔΔCq method (20).
Methyl thiazolyl tetrazolium (MTT)
Cell viability was detected by MTT
assay
After transfection, A549 and SK-MES-1 cells were
placed in 96-well plates and cultured for 1-4 days. Then, MTT
solution (20 µl) was added into each well to incubate for 4 h at
37°C, then carefully removing the medium and adding 100 µl dimethyl
sulfoxide (Sigma; Merck KGaA, Darmstadt, Germany) per well to
dissolve the formazan crystals, the plates were read at a
wavelength of 490 nm to measure the absorbance of each well at 0,
24, 48, 72 and 96 h.
Transwell assay
Cell migration assays were performed using transwell
chambers, 8 mm pore size culture inserts were placed into the
24-well plates to separate the top and the lower chambers, and
1×105 cells were added into the top chambers and
RPMI-1640 medium with 20% FBS and seeded to the lower chambers as
an attractant, after cell incubation at 37°C, 5% CO2
atmosphere for 24 h, the media was removed. Cotton swabs was used
to remove the cells on the upper surface of the membrane, and 100%
methanol was used to fix the cells migrated to the lower membrane
and the 0.1% crystal violet was used to stain the cells. The
numbers of cells were quantified by a microscope (Olympus
Corporation, Tokyo, Japan).
Dual luciferase reporter assay
The recombinan pMIR-reportor luciferase vector was
used for TCTN1 3′UTR luciferase assays. The wild-type or mutated
miR-1256 binding site targeted TCTN1 3′UTR were cloned into the
pMIR-reportor luciferase vector. Cells transfected with control
mimic and miR-1256 mimic were collected using
Lipofectamine®2000. The Dual Luciferase Reporter Assay
System (Promega Corporation, Madison, WI, USA) was used to measure
the luciferase activity after transfection for 48 h.
Statistical analysis
All experiments were repeated in triplicate,
GraphPad Prism 5.02 software (GraphPad Software, Inc., La Jolla,
CA, USA) and SPSS 16.0 software (SPSS, Inc., Chicago, IL, USA) were
used to perform statistical analyses, Student's t-test or post hoc
test after one-way analysis of variance in SPSS were used to
analyze the differences between the groups. Tukey's post hoc test
was the post hoc test used following one-way analysis of variance.
A P<0.05 was considered to indicate a statistically significant
difference.
Results
MiR-1256 expression is reduced in
NSCLC cells
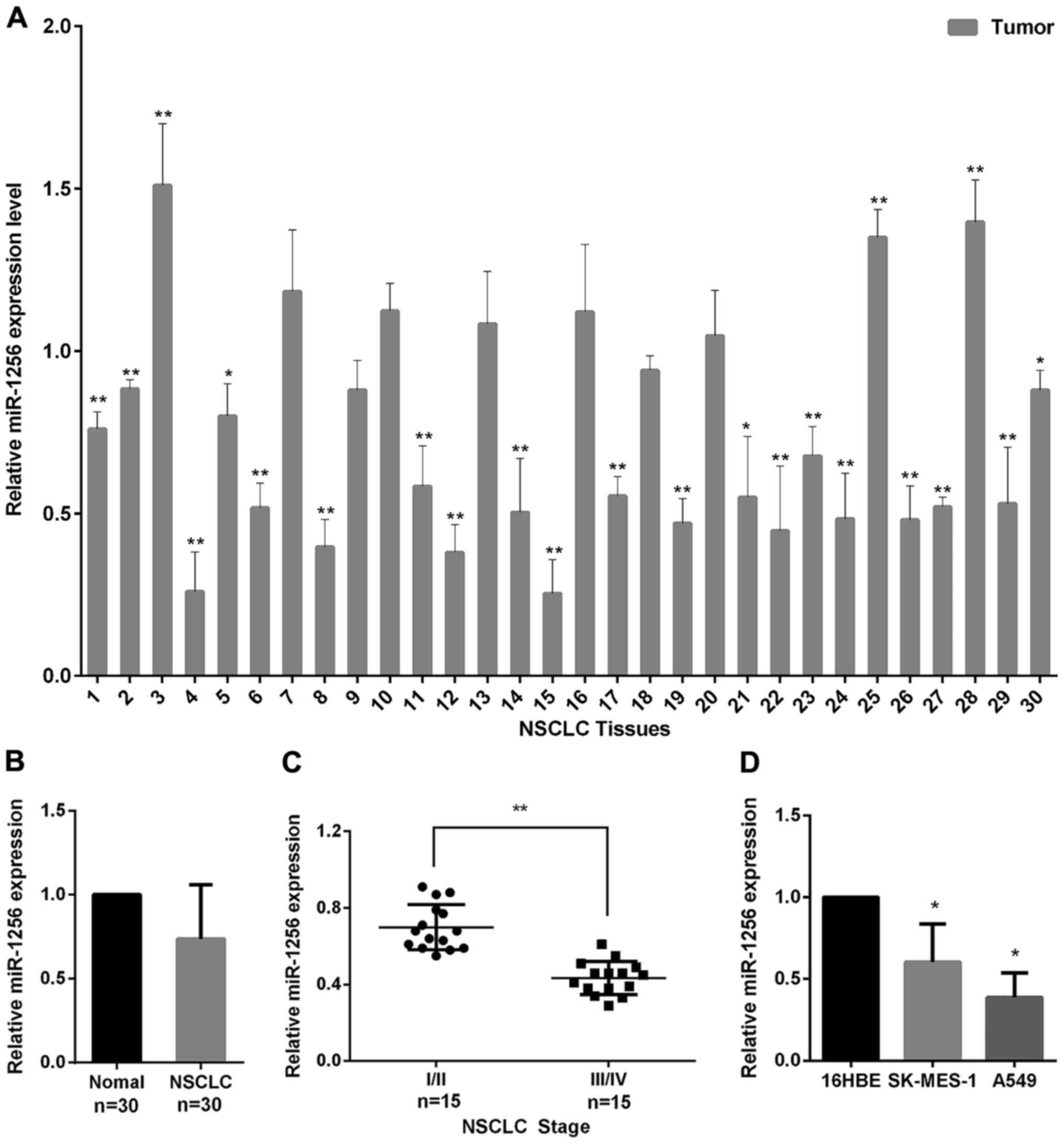
We used qRT-PCR to examine the miR-1256 levels in 30
paired of NSCLC and normal tissues to explore the role of miR-1256
in NSCLC. As seen in Fig. 1A, we
found that miR-1256 levels were frequently downregulated in 22/30
(73%) of the NSCLC tissues, and the miR-1256 average expression in
tumor tissues was decreased compared with the normal tissues
(Fig. 1B). We also detected miR-1256
expression in NSCLC tissues of varying stages, the result shows
that, the expression of miR-1256 in stages III–IV (higher stage
lesions) was significantly lower than in stages I–II (lower stage
lesions) (Fig. 1C). In addition, the
miR-1256 expression in A549 and SK-MES-1 cell lines was also
markedly downregulated, compared with normal 16HEB cells (Fig. 1D). Taken together, these data suggest
that miR-1256 may play a tumor suppressive role in the progression
of NSCLC.
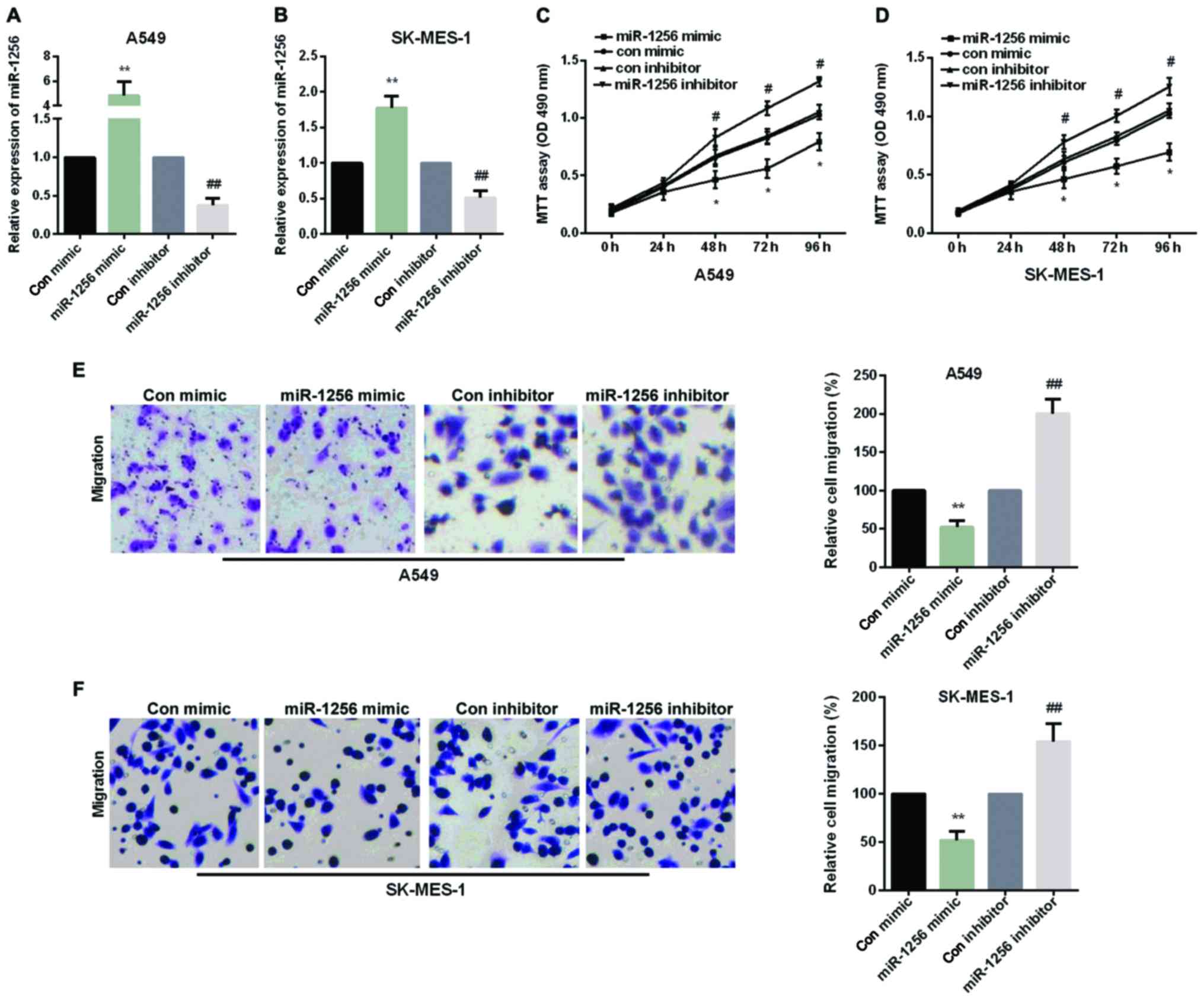
MiR-1256 inhibited cell proliferation
and migration of NSCLC
We performed an experiment to explore whether
re-expression of miR-1256 could contribute to the NSCLC cell
proliferation and migration. The A549 cells and SK-MES-1 cells were
transfected with miR-1256 mimic or miR-1256 inhibitor, as shown in
Fig. 2A and B. MTT assays was used to
examine the effect of miR-1256 on cell proliferation in A549 and
SK-MES-1 cells and transwell assays were used to detect the effect
of the miR-1256 on cell migration in A549 and SK-MES-1 cells. As
shown in Fig. 2C and D, cellular
proliferation data showed that transfection with miR-1256 mimic
exhibited a significant decrease to cell growth after 48 h, while
miR-1256 inhibitor increased cell proliferation remarkably in both
NSCLC cell lines. The results of transwell showed that, after
transfection with miR-1256 mimic, the migration ability of cells
was reduced distinctly in two NSCLC cell lines and transfection
with miR-1256 inhibitor, the migration ability of cells was
significantly augmented (Fig. 2E and
F).
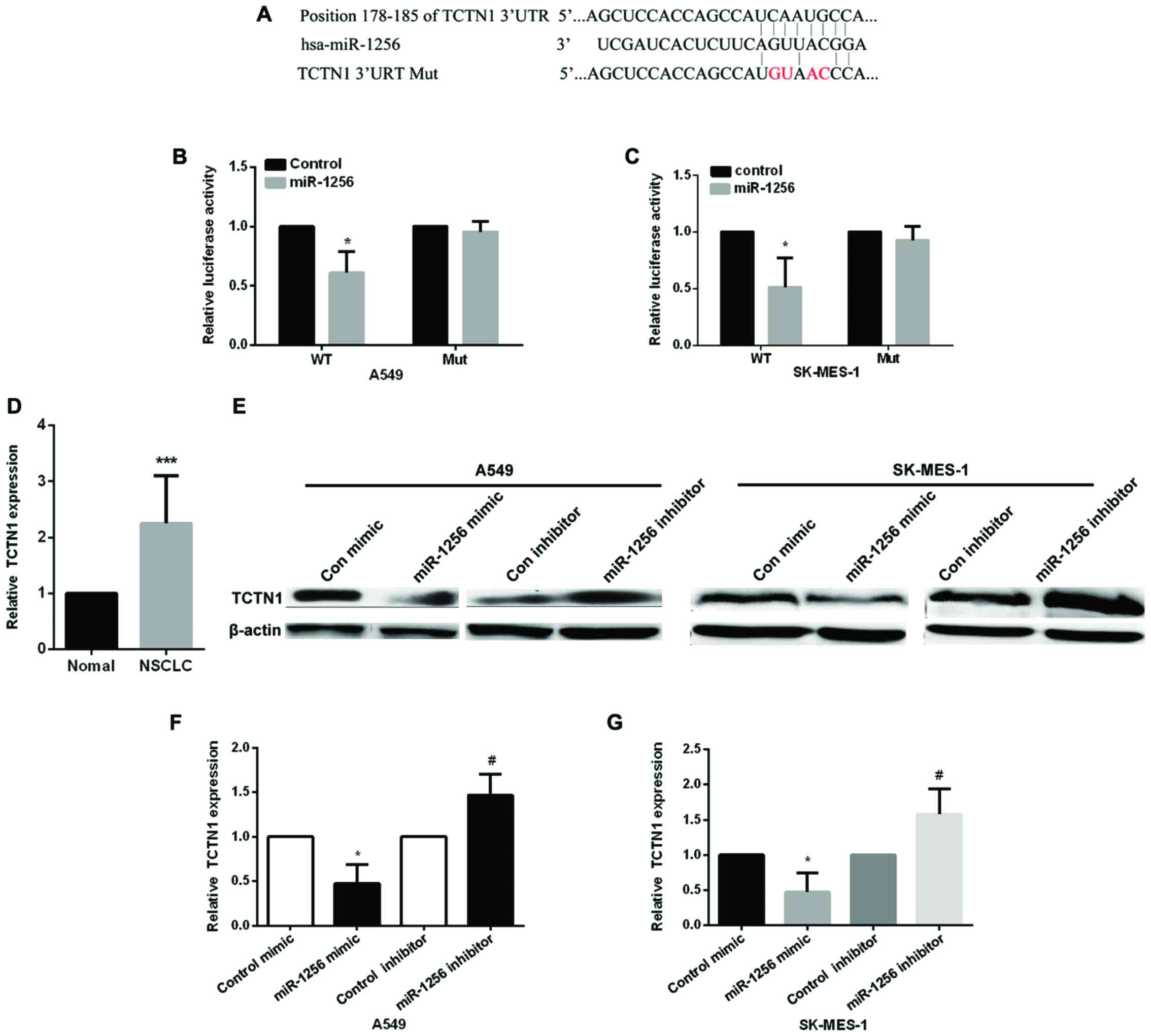
MiR-1256 directly targets TCTN1
Next, we detected the underlying mechanism of
miR-1256 in lung cancer cells, we used the TargetScan prediction
programs (www.targetscan.org) to predict the
potential targets of miR-1256, the results showed that miR-1256 may
target the 3′-UTR of TCTN1 (Fig. 3A),
the wild-type and mutant type of TCTN1 3′-UTR was constructed to
psiCHECK-2 vector to validate whether miR-1256 regulated NSCLC
cells by regulating TCTN1, then, we used luciferase reporter
vectors transfected into A549 and SK-MES-1 cells to detect the
luciferase activity (Fig. 3B and C),
compared with the control group, miR-1256 mimic markedly decreased
the relative luciferase activity in cells transfected with
wild-type TCTN1 3′-UTR, but it has no significantly changed on mut
type. These data proved that TCTN1 is regulated by miR-1256 in
NSCLC cells. We then detected TCTN1 expression in NSCLC tissues and
found that the expression of TCTN1 was increased (Fig. 3D). Next, we transfected two NSCLC cell
lines with miR-1256 mimic or inhibitor to determine the effect of
miR-1256 on the expression of TCTN1 to further confirm the above
results (Fig. 3E and G), the protein
level and mRNA of TCTN1 was markedly downregulated in both two
NSCLC cell lines of the ectopic expression of miR-1256, while
inhibiting miR-1256 significantly upregulated the protein and mRNA
levels of TCTN1.
Knockdown of TCTN1 expression inhibits
NSCLC cell proliferation and migration
We used TCTN1-specific siRNA to knock down TCTN1 in
A549 cells to examine the potential carcinogenicity of TCTN1 in
lung cancer, the transfection efficiency of TCTN1 siRNA was shown
in Fig. 4A and B. MTT assay and
transwell assay results showed that knockdown of TCTN1 decreased
the proliferative and migratory ability of A549 cells (Fig. 4C and D), TCTN1 siRNA deprived the
promoting effect of miR-1256 inhibitor on cell proliferation and
migration, suggesting that TCTN1 is regulated by miR-1256 in
regulating of lung cancer cells (Fig. 4E
and F).
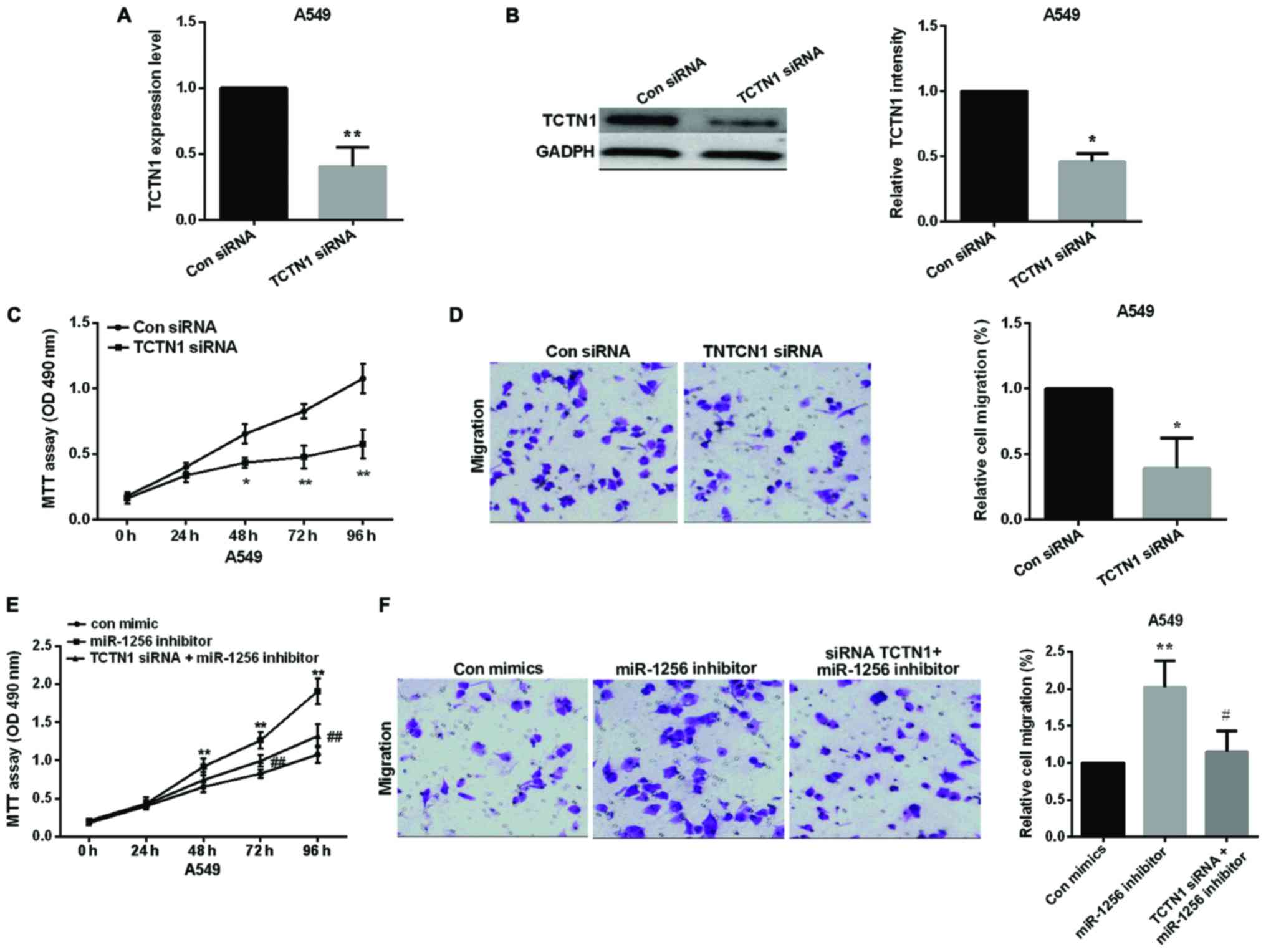 | Figure 4.Knockdown of TCTN1 inhibits NSCLC cell
proliferation and migration and TCTN1 reversed partial function of
miR-1256. (A and B) The TCTN1 expression level in NSCLC cells after
transfected with TCTN1 siRNA was detected by qRT-PCR and western
blot analysis, *P<0.05, **P<0.01. (C and D) MTT and transwell
assay, A549 cell proliferation and migration after knockdown of
TCTN1, *P<0.05, **P<0.01. (E and F) Treatment with both
miR-1256 inhibitor and siRNA TCTN1 reversed the promotion of A549
cell proliferative and migratory capacities compared with treatment
with miR-1256 inhibitor alone, **P<0.01; #P<0.05;
##P<0.01. TCTN1, tectonic family member 1; NSCLC,
non-small cell lung cancer; qRT-PCR, quantitative real-time PCR;
MTT, methyl thiazolyl tetrazolium assay. |
Discussion
Substantial evidence has stated that various miRNAs
play critical roles in the regulating of NSCLC development by
inducing mRNA degradation or inhibiting translation of a target
gene (21). MiRNAs functioned as
either oncogenes or suppressor genes in NSCLC (22). For example, Yang et al found
that miR-126 expression level in NSCLC was downregulated and
associated with poor survival as a tumor suppressor (23). However, miR-17-92 expression was
increased in lung cancers, and overexpression of miR-17-92 enhanced
cell proliferation (24). MiR-21 was
proven to downregulate PTEN expression and to stimulate NSCLC
growth and invasion (25). A previous
study showed that miR-31 targeted a tumor suppressor in regulating
lung cancer as an oncogene (26). A
study stated that miR-1256 expression was reduced in prostate
cancer (12). However, the major role
of miR-1256 in NSCLC is unclear. We first examined the effect of
miR-1256 in NSCLC cells to understand the role of miR-1256 in
NSCLC. We found that miR-1256 was markedly downregulated, and
playing a critical role in the tumorigenesis and progression of
NSCLC. We also confirmed the inhibitory effect of miR-1256 in NSCLC
cell proliferation and migration, overexpression of miR-1256 could
reduce the proliferative and migratory cell numbers, whereas,
miR-1256 inhibitor facilitated cell migration and proliferation.
Furthermore, we first identified that miR-1256 directly targeted
TCTN1 in regulating of the development of NSCLC using luciferase
assay.
TCTN1 was proved to participate in various
developmental processes as a novel oncogene and the expression of
TCTN1 in multiple solid tumors was abnormal (27). TCTN1 was also involved in a variety of
biological processes. Many studies have reported that knockdown of
TCTN1 inhibited cell growth in different types of malignant tumor
cells. TCTN1 participated in the growth of gastric cancer cells,
prostate cancer cells, human glioblastoma, pancreatic cancer and
medulloblastoma (16,17,19,16). In
the present study, we found that TCTN1 expression in NSCLC was
increased compared with normal tissues, and inhibiting TCTN1 could
suppress the proliferation and migration of NSCLC cells. The
protein and mRNA expression showed that the expression of miR-1256
and TCTN1 was negatively correlated in NSCLC. Additionally, we
found that TCTN1 could reverse partial function of miR-1256 in
regulating NSCLC development.
Collectively, the above study proved that miR-1256
played an important role in regulating NSCLC cell proliferation and
migration. Moreover, miR-1256 mimic inhibited NSCLC cell
proliferation and migration via directly targeting TCTN1.
Therefore, miR-1256 could be used as a therapeutic cell target for
treating NSCLC patients.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ and XH contributed to the conception of the
study. LW and WL contributed significantly to the data analysis and
study preparation. ZM, WL and FL performed the data analyses and
wrote the study. XW helped perform the analysis with constructive
discussions. All authors have read and approved the final
study.
Ethics approval and consent to
participate
All this experiments were approved by the Ethics
Committee of People's Hospital of Rizhao (Rizhao, China). Patients
signed informed consent.
Consent for publication
Not applicable.
Competing interests
Authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee CT, Risom T and Strauss WM: MicroRNAs
in mammalian development. Birth Defects Res C Embryo Today.
78:129–139. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bueno MJ, Pérez de Castro I and Malumbres
M: Control of cell proliferation pathways by microRNAs. Cell Cycle.
7:3143–3148. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jovanovic M and Hengartner MO: miRNAs and
apoptosis: RNAs to die for. Oncogene. 25:6176–6187. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cui H and Yang L: Analysis of microRNA
expression detected by microarray of the cerebral cortex after
hypoxic-ischemic brain injury. J Craniofac Surg. 24:2147–2152.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shi C and Zhang Z: Screening of
potentially crucial genes and regulatory factors involved in
epithelial ovarian cancer using microarray analysis. Oncol Lett.
14:725–732. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma DH, Li BS, Liu JJ, Xiao YF, Yong X,
Wang SM, Wu YY, Zhu HB, Wang DX and Yang SM: miR-93-5p/IFNAR1 axis
promotes gastric cancer metastasis through activating the STAT3
signaling pathway. Cancer Lett. 408:23–32. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shen S, Huang K, Wu Y, Ma Y, Wang J, Qin F
and Ma J: A miR-135b-TAZ positive feedback loop promotes
epithelial-mesenchymal transition (EMT) and tumorigenesis in
osteosarcoma. Cancer Lett. 407:32–44. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yuan Z, Baker K, Redman MW, Wang L, Adams
SV, Yu M, Dickinson B, Makar K, Ulrich N, Böhm J, et al: Dynamic
plasma microRNAs are biomarkers for prognosis and early detection
of recurrence in colorectal cancer. Br J Cancer. 117:1202–1210.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Purfield DC, Bradley DG, Kearney JF and
Berry DP: Genome-wide association study for calving traits in
Holstein-Friesian dairy cattle. Animal. 8:224–35. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Y, Kong D, Ahmad A, Bao B, Dyson G and
Sarkar FH: Epigenetic deregulation of miR-29a and miR-1256 by
isoflavone contributes to the inhibition of prostate cancer cell
growth and invasion. Epigenetics. 7:940–949. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Clark HF, Gurney AL, Abaya E, Baker K,
Baldwin D, Brush J, Chen J, Chow B, Chui C, Crowley C, et al: The
secreted protein discovery initiative (SPDI), a large-scale effort
to identify novel human secreted and transmembrane proteins: a
bioinformatics assessment. Genome Res. 13:2265–2270. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Z, Gao Y, Liu Y, Chen J, Wang J, Gan
S, Xu D and Cui X: Tectonic-1 contributes to the growth and
migration of prostate cancer cells in vitro. Int J Mol Med.
36:931–938. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang X, Yu Q, Zhang Y, Ling Z and Yu P:
Tectonic 1 accelerates gastric cancer cell proliferation and cell
cycle progression in vitro. Mol Med Rep. 12:5897–5902. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dai X, Dong M, Yu H, Xie Y, Yu Y, Cao Y,
Kong Z, Zhou B, Xu Y, Yang T, et al: Knockdown of TCTN1 strongly
decreases growth of human colon cancer cells. Med Sci Monit.
26:452–461. 2017. View Article : Google Scholar
|
|
19
|
Meng D, Chen Y, Zhao Y, Wang J, Yun D,
Yang S, Chen J, Chen H and Lu D: Expression and prognostic
significance of TCTN1 in human glioblastoma. J Transl Med.
12:2882014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Del Vescovo V, Grasso M, Barbareschi M and
Denti MA: MicroRNAs as lung cancer biomarkers. World J Clin Oncol.
5:604–620. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang J, Lan H, Huang X, Liu B and Tong Y:
MicroRNA-126 inhibits tumor cell growth and its expression level
correlates with poor survival in non-small cell lung cancer
patients. PloS one. 7:e429782012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hayashita Y, Osada H, Tatematsu Y, Yamada
H, Yanagisawa K, Tomida S, Yatabe Y, Kawahara K, Sekido Y and
Takahashi T: A polycistronic microRNA cluster, miR-17-92, is
overexpressed in human lung cancers and enhances cell
proliferation. Cancer Res. 65:9628–9632. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang JG, Wang JJ, Zhao F, Liu Q, Jiang K
and Yang GH: MicroRNA-21 (miR-21) represses tumor suppressor PTEN
and promotes growth and invasion in non-small cell lung cancer
(NSCLC). Clin Chim Acta. 411:846–852. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu X, Sempere LF, Ouyang H, Memoli VA,
Andrew AS, Luo Y, Demidenko E, Korc M, Shi W and Preis M:
MicroRNA-31 functions as an oncogenic microRNA in mouse and human
lung cancer cells by repressing specific tumor suppressors. J Clin
Invest. 120:1298–309. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Reiter JF and Skarnes WC: Tectonic, a
novel regulator of the Hedgehog pathway required for both
activation and inhibition. Genes Dev. 20:22–27. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao S, Chen X, Wan M, Jiang X, Li C, Cui
Y and Kang P: Tectonic 1 is a key regulator of cell proliferation
in pancreatic cancer. Cancer Biother Radiopharm. 31:7–13. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jing J, Wang C, Liang Q, Zhao Y, Zhao Q,
Wang S and Ma J: Lentivirus-mediated knockdown of tectonic family
member 1 inhibits medulloblastoma cell proliferation. Int J Clin
Exp Med. 8:13127–13135. 2015.PubMed/NCBI
|