Introduction
Pancreatic cancer has a poor prognosis with a 5-year
survival of <5% and the fourth most deadly cancer (1). The poor prognosis of pancreatic cancer
is associated with its chemotherapy resistance and early metastasis
(2,3). Hence, the high mortality of pancreatic
cancer indicates a requirement for early detection and novel
therapies.
The PI3K/Akt pathway, a survival signaling pathway,
is involved in the control of cell apoptosis and proliferation and
>50% human cancers are related to the deregulation of this
pathway, including pancreatic cancer (4–7). Akt
contributes to cell plasticity in pancreas as a regulator and its
overexpression has been proved to be a common phenomenon in
pancreatic cancer (8–11). Therefore, the PI3K/Akt pathway plays
a crucial role in the development of pancreatic cancer.
Polo-like kinase 1 (Plk1), a serine/threonine
kinase, plays an important role in cell cycle and apoptosis
(12). Plk1 is overexpressed in
various tumors and its overexpression is associated with poor
prognosis (12,13). Plk1 overexpression is a common and
early event and contributes to gemcitabine resistance in pancreatic
cancer (14,15).
Apoptosis occurs in multicellular organisms as a
programmed cell death and it can be activated through intrinsic
pathway and extrinsic pathway (16). The intrinsic pathway involves the
mitochondria (17). Activation of
caspases is an important process for the intrinsic pathway and is
controlled by anti- or pro-apoptotic proteins, such as XIAP, Bcl-2
and BAX (18,19).
In cell cycle, PI3K/Akt-dependent phosphorylation of
Plk1-Ser99 is required for metaphase-anaphase transition and
Plk1-dependent phosphorylation of IRS2-S556 inhibits mitotic exit
through reducing Akt activity (20,21).
However, the mechanism of cell apoptosis induced by PI3K/Akt
pathway and Plk1 still remains unclear.
In this study, we investigated the function of
PI3K/Akt pathway on cell proliferation and apoptosis in pancreatic
carcinoma in vitro and in vivo and the effect of Plk1
in this process. Our results could help to understand
apoptosis-related pathway, such as caspase-related or Bcl-2
family-mediated pathway, and benefit the therapy of pancreatic
cancer.
Materials and methods
Cell culture and antibodies
Human pancreatic cancer cell lines (AsPC-1, BxPC-3,
PANC-1) were acquired from ATCC (Manassas, VA, USA) and maintained
in RPMI-1640 medium (Gibco, Gaithersburg, MD, USA), supplemented
with 100 U/ml penicillin, 100 U/ml streptomycin, and 10% fetal
bovine serum (FBS) (Gibco) at 37°C in 5% CO2. Antibodies
against p-Akt, Akt, Plk1, XIAP, caspase-3, cleaved caspase-3 were
purchased from Cell Signaling Technology (Beverly, MA, USA) for
western blot analysis. Antibodies against Bcl-2, BAX were purchased
from Abcam (Abcam, UK) for western blot analysis. Akt antibody for
immunohistochemistry (IHC) was purchased from Abcam (Abcam). GAPDH
and actin antibodies for western blot analysis were acquired from
Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Immunohistochemistry (IHC) and tissue
microarray
The pancreatic cancer tissue microarrays were
purchased from Xian Ailina Biotechnology Co., Ltd. (Xian Ailina
Biotechnology Co., Ltd., China). The expression of Akt was measured
by IHC as described (22). Akt
staining in human tissues were scored independently by two
pathologists, by evaluating a semi-quantitative immunoreactivity
score (IRS) as described (23).
Then, tissues with IRS 0–5 and IRS 6–9 were defined as low and high
expression of Akt, respectively.
Short hairpin RNA
The short hairpin RNA for Plk1 (shPlk1) and empty
vector (EV) was synthesized with the vector pYr-1.1 (hU6/EGFP/Neo)
(Changsha Yingrun Biotechnology Co., Ltd., China). Cell HEK 293
(ATCC, USA) was used to pack recombination adenovirus and the empty
vector (EV) was constructed as an experimental control.
Cell proliferation assay
The cell lines of AsPC-1, BxPC-3, and PANC-1 were
seeded in 96-well dishes at 1×104 cells per well. Cell
viability was determined by MTT assays after cells were incubated
with different concentrations of LY294002 (Cell Signaling
Technology, USA) for 24 h.
Quantitative reverse transcription-PCR
(qRT-PCR) assay
The mRNA level was quantified by qRT-PCR. Primers
used for the Akt gene: forward, 5′-TCACCATCACACCACCT GAC-3′; and
reverse, 5′-CTCAAATGCACCCGAGAAAT-3′. Primers used for the Plk1
gene: forward, 5′-ACC AGC ACG TCG TAG GAT TC-3′; and reverse 5′-ATA
ACT CGG TTT CGG TGC AG-3′. Primers used for the GAPDH gene were
5′-AAC GGA TTT GGT CGT ATT GG-3′ (forward) and 5′-GGA TCT CGC TCC
TGG AAG AT-3′ (reverse) (Invitrogen, USA).
Western blot analysis
Cells were washed in cold PBS twice, then
solubilized in RIPA lysis buffer (Vazyme, Nanjing, China). Samples
with the same amount of protein were analyzed by western blotting
as described (24).
Flow cytometry
Apoptosis was detected by flow cytometry. The cells
were incubated with LY294002 for 48 h or recombinant adenovirus for
24, 48 and 72 h; then washed twice with ice-cold PBS and
resuspended in 1x binding buffer (BD Pharmingen, USA) at a
concentration of 1x105 cells/ml. Added 5 μl of
APC Annexin V (BD Pharmingen) and 5 μl of 7-AAD (BD
Pharmingen). Then, samples were analyzed by flow cytometer (BD
FACSCalibur equipped with CellQuest Pro).
Tumor xenograft model
Human pancreatic cancer cells (dissolved in PBS;
1×107/120 μl; BxPC-3) were injected
subcutaneously in the right subaxillary of female nude mice
(Chinese Academy of Sciences, Shanghai, China). When tumor volume
was 80–120 mm3, 12 nude mice were divided into two
groups (control group, LY294002 group) randomly. The LY294002 group
was injected with LY294002 (25 mg/kg/d; dissolved in DMSO) via the
abdominal route; and the control group was treated with equal 1%
DMSO via the abdominon. Tumor volume was observed every 2 days and
was calculated by the formula: (length × width2)/2. Mice
were sacrificed at day 12 and the weight and volume of the tumors
were measured. Then the tumor along the maximum transverse incision
was cut in half: one half was made into paraffin block after
fixation in 4% neutral formaldehyde and the other half was
cryopreserved at −80°C. All animals received human care and all
experiments were carried out according to the guidelines outlined
in the Guide for the Care and Use of Laboratory Animals.
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) assay
Paraffin blocks were cut into slices with 4
μm thickness. The TUNEL Brighted Apoptosis Detection kit
(Vazyme) was used to detect cellular apoptosis in tumor tissues
according to the instructions.
Statistical analysis
All statistical analyses were performed using
GraphPad Prism 5.0 software (GraphPad Software, Inc., La Jolla, CA,
USA). Each experiment was performed three times. All data were
expressed as mean ± SD unless otherwise specified. Data from each
group were statistically analyzed using a two-tailed Student's
t-test, except immunohistochemical score which was analyzed by
Chi-square test. Differences were considered statistically
significant at P<0.05. P-values of the statistical significances
for differences were set as *P<0.05,
**P<0.01 and ***P<0.001, as shown in
the figures.
Results
Akt is overexpressed in pancreatic
cancer
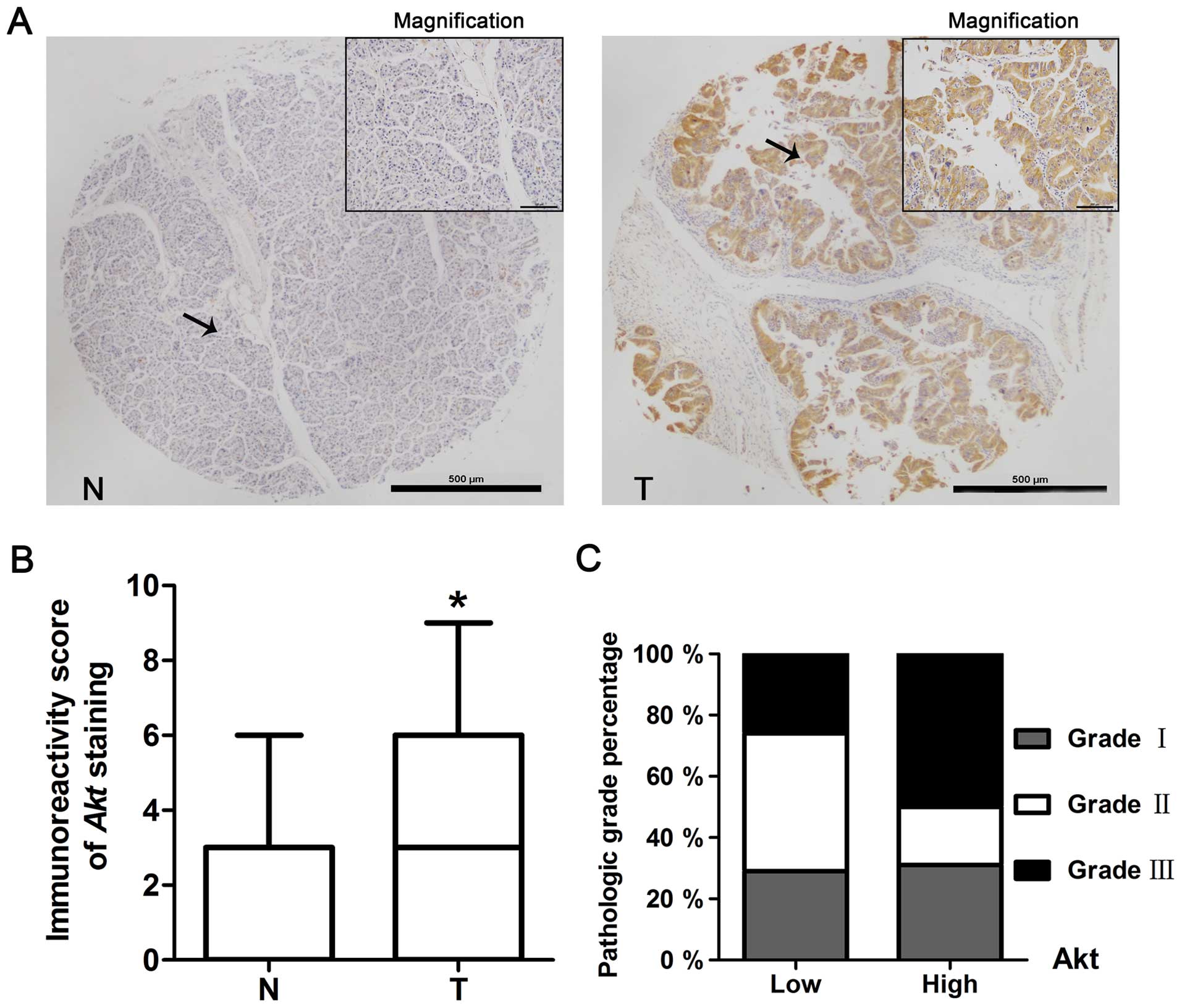
Immunohistochemistry staining was used to detect the
expression of Akt in pancreatic cancer. The staining of pancreatic
tissue micro-array (including 51 cases of normal pancreas and 91
cases of pancreatic cancer) showed that 27% of normal tissues had
Akt positive staining (88% with low expression, IRS 0–5; 12% with
high expression, IRS 6–12), whereas 59% of tumor tissues had Akt
positive staining (71% with low expression, IRS 0–5; 29% with high
expression, IRS 6–12) (Fig. 1A and
B, Table I). The expression of
Akt in tumor tissues was significantly higher than it in normal
tissues (P<0.05).
 | Table IExpression of Akt in human pancreas
and pancreatic cancer. |
Table I
Expression of Akt in human pancreas
and pancreatic cancer.
| Group | Expression of Akt
in pancreatic cancer
|
|---|
| n | Low expression
(%) | High expression
(%) | P-value |
|---|
| Normal tissue | 51 | 45 (88) | 6 (12) |
0.021a |
| Pancreatic
cancer | 91 | 65 (71) | 26 (29) | |
Correlation of Akt levels with clinical
characteristics
According to the study of the relationship between
the expression level of Akt in pancreatic cancer tissues and the
clinical characteristics of these patients, there was no
significantly difference among gender, age or clinical stage.
However, Akt expression level was correlated with pathologic grade
and there were more cancer tissues with Akt high expression in
grade 3 than those in grade 1–2 (grade 1–2 with 79% low expression
and 21% high expression of Akt; grade 3 with 57% low expression and
43% high expression of Akt; P<0.05) (Fig. 1C and Table II).
| Table IIThe relationship between the
expression level of Akt in pancreatic cancer and the clinical
characteristics of the patients. |
Table II
The relationship between the
expression level of Akt in pancreatic cancer and the clinical
characteristics of the patients.
|
Characteristics | Expression of Akt
in pancreatic cancer
|
|---|
| n | Low expression
(%) | High expression
(%) | P-value |
|---|
| Gender | | | | 0.842 |
| M | 47 | 34 (72) | 13 (28) | |
| F | 44 | 31 (70) | 13 (30) | |
| Age | | | | 0.273 |
| <60 | 57 | 43 (75) | 14 (25) | |
| ≥60 | 34 | 22 (65) | 12 (35) | |
| Pathologic
grade | | | | 0.029 |
| 1–2 | 61 | 48 (79) | 13 (21) | |
| 3 | 30 | 17 (57) | 13 (43) | |
| Clinical stage | | | | 0.602 |
| I–II | 78 | 57 (73) | 21 (27) | |
| II–IV | 13 | 8 (62) | 5 (38) | |
Inhibition of the PI3K/Akt pathway could
reduce cell proliferation and induce apoptosis in pancreatic cancer
cells in vitro
Cell proliferation influenced by different
concentrations of LY294002 was detected by MTT assay (Fig. 2A). The optimal concentrations of
LY294002 for different pancreatic cancer cell lines were
IC50AsPC-1=40 μM, IC50BxPC-3=15
μM and IC50PANC-1=35 μM, respectively.
Then qRT-PCR showed LY294002 had no effect on mRNA level of Akt
(Fig. 2B). While protein level of
p-Akt was downregulated and it suggested that LY294002 influenced
PI3K/Akt pathway through decreasing phosphorylation of Akt
(Fig. 2C). As described previously,
the PI3K/Akt pathway is an important survival signaling pathway in
pancreatic cancer and it is supposed to be related to cell
apoptosis in pancreatic cancer. Compared to the control group,
using LY294002 to block PI3K/Akt pathway it could induce
significant apoptosis (P<0.001) (Fig. 2D and E). Then we detected the
expression level of apoptosis-related proteins such as XIAP, Bcl-2,
BAX and caspase-3 by western blotting and found that both
caspase-dependent signaling pathway (downregulating XIAP and
upregulating cleaved caspase-3) and Bcl-2 family mediated signaling
pathway (downregulating Bcl-2 and upregulating BAX) contributed to
the apoptosis induced by LY294002 (Fig.
2C). That means PI3K/Akt pathway can control cell survival
through several routes and LY294002 might become an ideal drug to
induce cancer cell apoptosis.
Plk1 plays a crucial role in the
pancreatic cancer cell apoptosis induced by PI3K/Akt pathway
inhibition
Plk1 is essential for cell cycle regulation and our
previous study showed Plk1 overexpression is correlated to cell
proliferation and chemotherapy resistance in pancreatic cancer
(25). As PI3K/Akt pathway is
reported to be linked to anti-apoptotic signal transduction and
chemoresistance of pancreatic cancer (26,27),
the relationship between Plk1 and PI3K/Akt pathway is worthy of
study and concern. Then we constructed recombinant adenovirus
rAd-Plk1-shRNAs (shPlk1) to further confirm the effect of Plk1 in
the process of apoptosis induced by PI3K/Akt pathway inhibition.
LY294002 can downregulate Plk1 mRNA level as well as Plk1 protein
level (P<0.01) (Fig. 3A and B)
while Plk1 suppression has no influence on the expression of Akt
(Fig. 3C and D). Flow cytometry
showed Plk1 knockdown could induce significant apoptosis in
pancreatic cancer cells as cancer cells infected with shPlk1 for 48
h had obviously higher apoptosis rate than the EV group (P<0.01)
(Fig. 3E and F). We also found that
Plk1 knockdown induced cancer cell apoptosis through downregulating
XIAP, Bcl-2 and upregulating cleaved caspase-3 and BAX (Fig. 3D). Thus as a result, we believe that
LY294002 could activate apoptosis-related pathway, such as
caspase-related or Bcl-2 family-mediated pathway through
downregulating Plk1.
PI3K/Akt pathway inhibition suppressed
pancreatic cancer growth and induced apoptosis in vivo
Finally, we investigated the effect of PI3K/Akt
pathway inhibition in vivo through a tumor xenograft model.
In the BxPC-3 xenograft model, both the tumor volume (1,122±226 vs
260±94 mm3; P<0.01) and the weight (1,025±161 vs
449±65 mg; P<0.01) of LY294002 group were significantly less
than the control group (Fig. 4A–C).
Hematoxylin-eosin staining showed more necrosis and higher
expression of cleaved caspase-3 in LY294002 group (Fig. 4D). TUNEL assay revealed LY294002
induced pancreatic cancer cell apoptosis in the tumor xenograft
model (Fig. 4E). All the data above
indicated that inhibition of PI3K/Akt pathway could suppress
pancreatic cancer growth and induced tumor necrosis and cell
apoptosis in vivo.
Discussion
Pancreatic cancer is a malignant tumor with 5-year
survival <5%, despite years of efforts, ~80% of patients have
locally advanced or metastatic disease when they are diagnosed
(28).
PI3K/Akt pathway is important in the development and
progression of pancreatic cancer and its activation is a common
event during this process (29–31).
Schlieman et al (10) found
activation of Akt existed in half of pancreatic cancer cases, which
is consistent with the result of our study. Our study also revealed
that the expression level of Akt was closely related to the
pathologic grade of pancreatic cancer (Table II). Thus, inhibition of PI3K/Akt
pathway could be a potential therapeutic target for the treatment
of pancreatic cancer. Kim et al (32) showed Akt inhibition could enhance
chemosensitivity of gemcitabine in pancreatic cancer, Bondar et
al (33) and our study showed
inhibition of PI3K/Akt pathway induces cell apoptosis and reduces
cell proliferation in pancreatic cancer, but the mechanism of this
process is unclear.
Apoptosis plays an important role in the growth and
development of mammals and its deregulation results in many
diseases, especially tumorigenesis (34–36).
There are two apoptosis pathways: intrinsic pathway and extrinsic
pathway. Most apoptosis are induced by the intrinsic pathway which
is activated by intracellular signals generated such as caspase
signaling pathway when cells are stressed (37). It is reported that XIAP is
phosphorylated by Akt at residue serine 87 in vitro and
in vivo and this process results in resistance to
cisplatin-induced XIAP degradation, caspase-3 activation, and
apoptosis (38). Our study showed
similar results that XIAP was downregulated and cleaved caspase-3
was upregulated after PI3K/Akt pathway was inhibited by LY294002 in
pancreatic cancer. Moreover, we explored whether Bcl-2 family
mediated apoptosis is related to PI3K/Akt pathway inhibition and
found Bcl-2/BAX ratio was decreased after pancreatic cancer cells
were treated with LY294002, indicating that PI3K/Akt pathway
inhibition can induce cancer cell apoptosis by activating Bcl-2
family-mediated apoptosis pathway.
As described previously, both PI3K/Akt pathway and
Plk1 play an important role in the development, progression and
chemoresistance of pancreatic cancer and PI3K/Akt-dependent
phosphorylation of Plk1-Ser99 is required for metaphase-anaphase
transition (20,25–27).
Our previous study revealed Plk1 knockdown could lead to cell cycle
arrest and enhance chemosensitivity to gemcitabine in pancreatic
cancer. Our present study indicated that Plk1 knockdown could
induce apoptosis in pancreatic cancer through downregulating XIAP,
Bcl-2 and upregulating cleaved caspase-3 and BAX. At the same time,
PI3K inhibitor LY294002 downregulated Plk1 and induce apoptosis. It
means that PI3K/Akt pathway inhibition can induce cancer cell
apoptosis through activating the caspase pathway and decreasing
Bcl-2/BAX ratio via suppression of Plk1 expression.
In conclusion, this study implied that PI3K/Akt
pathway inhibition could suppress cell proliferation and lead to
cell apoptosis in pancreatic cancer. Apoptosis induced by PI3K/Akt
pathway inhibition is correlated with the expression of Plk1 and
downregulating Plk1 can activate apoptosis-related pathway, such as
caspase-related and Bcl-2 family-mediated pathway. These results
indicate that combination therapy, especially targeting to PI3K/Akt
pathway might be an achievable access for the treatment of
pancreatic cancer.
Acknowledgments
This study was supported in part by grants from the
National Natural Science Foundation of China (nos. 30972910 and
81172269) and Natural Science Foundation of Jiangsu Province, P.R.
China (no. 050104313).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bilimoria KY, Bentrem DJ, Ko CY, Stewart
AK, Winchester DP and Talamonti MS: National failure to operate on
early stage pancreatic cancer. Ann Surg. 246:173–180. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Duong HQ, Kim HJ, Kang HJ, Seong YS and
Bae I: ZSTK474, a PI3K inhibitor, suppresses proliferation and
sensitizes human pancreatic adenocarcinoma cells to gemcitabine.
Oncol Rep. 27:182–188. 2012.
|
|
4
|
Yuan TL and Cantley LC: PI3K pathway
alterations in cancer: Variations on a theme. Oncogene.
27:5497–5510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bellacosa A, Kumar CC, Di Cristofano A and
Testa JR: Activation of AKT kinases in cancer: Implications for
therapeutic targeting. Adv Cancer Res. 94:29–86. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Michl P and Downward J: Mechanisms of
disease: PI3K/AKT signaling in gastrointestinal cancers. Z
Gastroenterol. 43:1133–1139. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Falasca M, Selvaggi F, Buus R, Sulpizio S
and Edling CE: Targeting phosphoinositide 3-kinase pathways in
pancreatic cancer - from molecular signalling to clinical trials.
Anticancer Agents Med Chem. 11:455–463. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Edling CE, Selvaggi F, Buus R, Maffucci T,
Di Sebastiano P, Friess H, Innocenti P, Kocher HM and Falasca M:
Key role of phosphoinositide 3-kinase class IB in pancreatic
cancer. Clin Cancer Res. 16:4928–4937. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yamamoto S, Tomita Y, Hoshida Y, Morooka
T, Nagano H, Dono K, Umeshita K, Sakon M, Ishikawa O, Ohigashi H,
et al: Prognostic significance of activated Akt expression in
pancreatic ductal adenocarcinoma. Clin Cancer Res. 10:2846–2850.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schlieman MG, Fahy BN, Ramsamooj R,
Beckett L and Bold RJ: Incidence, mechanism and prognostic value of
activated AKT in pancreas cancer. Br J Cancer. 89:2110–2115. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Elghazi L, Weiss AJ, Barker DJ, Callaghan
J, Staloch L, Sandgren EP, Gannon M, Adsay VN and Bernal-Mizrachi
E: Regulation of pancreas plasticity and malignant transformation
by Akt signaling. Gastroenterology. 136:1091–1103. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Strebhardt K: Multifaceted polo-like
kinases: Drug targets and antitargets for cancer therapy. Nat Rev
Drug Discov. 9:643–660. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Holtrich U, Wolf G, Bräuninger A, Karn T,
Böhme B, Rübsamen-Waigmann H and Strebhardt K: Induction and
down-regulation of PLK, a human serine/threonine kinase expressed
in proliferating cells and tumors. Proc Natl Acad Sci USA.
91:1736–1740. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Weichert W, Schmidt M, Jacob J, Gekeler V,
Langrehr J, Neuhaus P, Bahra M, Denkert C, Dietel M and Kristiansen
G: Overexpression of Polo-like kinase 1 is a common and early event
in pancreatic cancer. Pancreatology. 5:259–265. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Song B, Liu XS, Rice SJ, Kuang S, Elzey
BD, Konieczny SF, Ratliff TL, Hazbun T, Chiorean EG and Liu X: Plk1
phosphorylation of orc2 and hbo1 contributes to gemcitabine
resistance in pancreatic cancer. Mol Cancer Ther. 12:58–68. 2013.
View Article : Google Scholar :
|
|
16
|
Lo ACY, Woo TTY, Wong RL and Wong D:
Apoptosis and other cell death mechanisms after retinal detachment:
Implications for photoreceptor rescue. Ophthalmologica. 226(Suppl
1): 10–17. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Christensen ME, Jansen ES, Sanchez W and
Waterhouse NJ: Flow cytometry based assays for the measurement of
apoptosis-associated mitochondrial membrane depolarisation and
cytochrome c release. Methods. 61:138–145. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fulda S and Debatin KM: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fulda S, Galluzzi L and Kroemer G:
Targeting mitochondria for cancer therapy. Nat Rev Drug Discov.
9:447–464. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kasahara K, Goto H, Izawa I, Kiyono T,
Watanabe N, Elowe S, Nigg EA and Inagaki M: PI 3-kinase-dependent
phosphorylation of Plk1-Ser99 promotes association with 14-3-3
gamma and is required for metaphase-anaphase transition. Nat
Commun. 4:18822013. View Article : Google Scholar
|
|
21
|
Chen L, Li Z, Ahmad N and Liu X: Plk1
phosphorylation of IRS2 prevents premature mitotic exit via AKT
inactivation. Biochemistry. 54:2473–2480. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang S, Wu X, Zhang J, Chen Y, Xu J, Xia
X, He S, Qiang F, Li A, Shu Y, et al: CHIP functions as a novel
suppressor of tumour angiogenesis with prognostic significance in
human gastric cancer. Gut. 62:496–508. 2013. View Article : Google Scholar
|
|
23
|
Weichert W, Röske A, Gekeler V, Beckers T,
Ebert MP, Pross M, Dietel M, Denkert C and Röcken C: Association of
patterns of class I histone deacetylase expression with patient
prognosis in gastric cancer: A retrospective analysis. Lancet
Oncol. 9:139–148. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jia L, Xing J, Ding Y, Shen Y, Shi X, Ren
W, Wan M, Guo J, Zheng S, Liu Y, et al: Hyperuricemia causes
pancreatic β-cell death and dysfunction through NF-κB signaling
pathway. PLoS One. 8:e782842013. View Article : Google Scholar
|
|
25
|
Yu C, Zhang X, Sun G, Guo X, Li H, You Y,
Jacobs JL, Gardner K, Yuan D, Xu Z, et al: RNA
interference-mediated silencing of the polo-like kinase 1 gene
enhances chemosensitivity to gemcitabine in pancreatic
adenocarcinoma cells. J Cell Mol Med. 12A:2334–2349. 2008.
View Article : Google Scholar
|
|
26
|
Ng SS, Tsao MS, Nicklee T and Hedley DW:
Wortmannin inhibits pkb/akt phosphorylation and promotes
gemcitabine antitumor activity in orthotopic human pancreatic
cancer xenografts in immunodeficient mice. Clin Cancer Res.
7:3269–3275. 2001.PubMed/NCBI
|
|
27
|
Ng SSW, Tsao MS, Chow S and Hedley DW:
Inhibition of phosphatidylinositide 3-kinase enhances
gemcitabine-induced apoptosis in human pancreatic cancer cells.
Cancer Res. 60:5451–5455. 2000.PubMed/NCBI
|
|
28
|
Costello E, Greenhalf W and Neoptolemos
JP: New biomarkers and targets in pancreatic cancer and their
application to treatment. Nat Rev Gastroenterol Hepatol. 9:435–444.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
di Magliano MP and Logsdon CD: Roles for
KRAS in pancreatic tumor development and progression.
Gastroenterology. 144:1220–1229. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Stephen AG, Esposito D, Bagni RK and
McCormick F: Dragging ras back in the ring. Cancer Cell.
25:272–281. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hofmann I, Weiss A, Elain G, Schwaederle
M, Sterker D, Romanet V, Schmelzle T, Lai A, Brachmann SM,
Bentires-Alj M, et al: K-RAS mutant pancreatic tumors show higher
sensitivity to MEK than to PI3K inhibition in vivo. PloS One.
7:e441462012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim R, Yamauchi T, Husain K, Sebti S and
Malafa M: Triciribine phosphate monohydrate, an AKT inhibitor,
enhances gemcitabine activity in pancreatic cancer cells.
Anticancer Res. 35:4599–4604. 2015.PubMed/NCBI
|
|
33
|
Bondar VM, Sweeney-Gotsch B, Andreeff M,
Mills GB and McConkey DJ: Inhibition of the phosphatidylinositol
3′-kinase-AKT pathway induces apoptosis in pancreatic carcinoma
cells in vitro and in vivo. Mol Cancer Ther. 1:989–997.
2002.PubMed/NCBI
|
|
34
|
Lowe SW and Lin AW: Apoptosis in cancer.
Carcinogenesis. 21:485–495. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Arlt A, Müerköster SS and Schäfer H:
Targeting apoptosis pathways in pancreatic cancer. Cancer Lett.
332:346–358. 2013. View Article : Google Scholar
|
|
36
|
Goldar S, Khaniani MS, Derakhshan SM and
Baradaran B: Molecular mechanisms of apoptosis and roles in cancer
development and treatment. Asian Pac J Cancer Prev. 16:2129–2144.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ivey R, Desai M, Green K, Sinha-Hikim I,
Friedman TC and Sinha-Hikim AP: Additive effects of nicotine and
high-fat diet on hepatocellular apoptosis in mice: Involvement of
caspase 2 and inducible nitric oxide synthase-mediated intrinsic
pathway signaling. Horm Metab Res. 46:568–573. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dan HC, Sun M, Kaneko S, Feldman RI,
Nicosia SV, Wang HG, Tsang BK and Cheng JQ: Akt phosphorylation and
stabilization of X-linked inhibitor of apoptosis protein (XIAP). J
Biol Chem. 279:5405–5412. 2004. View Article : Google Scholar
|