Introduction
Alzheimer's disease (AD) is a neurodegenerative
disorder with progressive cognitive impairment. Molecular genetic
studies have demonstrated the presence of complex genetic
heterogeneity in AD (1–3). Previous investigations reported that
certain gene mutations are associated with familial early-onset AD,
such as amyloid-β precursor protein, presenilin I and presenilin II
(3–5). In addition, it was demonstrated that
there are likely numerous gene polymorphisms that are risk factors
for late-onset AD (LOAD), but to date, only the apolipoprotein E
(APOE) ε4 allele has been widely confirmed as a susceptibility gene
for LOAD (1). However, the APOE gene
polymorphism may only contribute to 50% of genetic susceptibility
(1). This suggests there may be
other genes that could influence susceptibility to the onset of
LOAD. The investigation of other susceptibility genes would
facilitate the research on LOAD pathogenesis.
In 1996, the clusterin (CLU) gene was
suggested to be a potential AD susceptibility gene (2). The CLU gene is located at
p21-p12 of chromosome 8 (5). The
protein expressed by CLU is known as apolipoprotein J which
is secreted by neurons and astrocytes, and being the second-largest
glycoprotein in the brain following APOE, it has similar biological
functions to APOE (5). CLU is
expressed in cerebrospinal fluid and amyloid plaques in brain
tissues, is able to combine with β-amyloid peptide to participate
in the processes of metabolism, transportation and clearance of
amyloid-β (Aβ), and has an important role in the cell cycle
regulation and lipid metabolism of AD (3–5). A
previous study reported that the CLU gene may be associated
with AD (2). Unfortunately, no gene
locus identified in the earliest study regarding the correlation
between CLU gene variants and AD (2).
With the advance of molecular genetic technology, it
became possible to include significantly greater numbers of
individuals in the disease-association studies, thereby allowing
the identification of more potential disease susceptibility genes.
In October 2009, ‘Nature Genetics’ simultaneously published two
AD-associated large-scale genome-wide association studies (GWAS)
completed by Lambert et al (6) and Harold et al (7), which both confirmed that the CLU
gene single nucleotide polymorphism (SNP) rs11136000 was associated
with LOAD. Another meta-analysis of genome-wide detection also
suggested that the p22-p21.1 of chromosome 8, namely the CLU
chromosomal locus, was a LOAD-associated susceptibility region
(8). Since then, the association
between AD and CLU variants has once again become the focus
of numerous studies, in which various experiments were carried out
in an attempt to demonstrated the correlation (9–26).
However, the results of these studies were not concordant.
The present investigation reviewed and conducted a
meta-analysis of previously published case-control studies on the
correlation between the CLU gene polymorphism rs11136000 and
LOAD both in China and abroad. The increased sample size could
improve the significance of the results, and subgroup analyses of
various ethnicities could eliminate possible ethnic differences.
The aim of the present study was to provide objective
evidence-based results for the association between CLU gene
polymorphisms and AD susceptibility.
Materials and methods
Subjects
The previously published case-control studies
regarding the association between CLU gene variants and LOAD
in various ethnic groups were reviewed. The patients with LOAD in
these studies were selected as study cases, and the CLU
rs11136000 polymorphism as the locus of interest, which in the
previous studies was demonstrated to be significant although
controversial. The control groups in these studies were healthy
participants of the same ethnicity as the cases studied. The
genotype frequencies of TT, TC and CC, as well as the allele
frequencies of T and C, in the case and control groups of these
studies were obtained for meta-analysis. The control groups in this
study were healthy participants with the same ethnicity as the
cases studied.
Literature search
A systematic search was conducted using PubMed
(www.ncbi.nlm.nih.gov/pubmed), EMbase
(store.elsevier.com/embase), CBMdisc
(www.cnki.com.cn), CMCC, Wanfang (www.wanfangdata.com.cn), and other databases in
English or Chinese (the last search was updated on August 31,
2014), with ‘Clusterin AND Alzheimer disease’ as MeSH terms, and
‘Clusterin AND Alzheimer’ and ‘rs11136000’ as free terms. Through
literature retrospection, the references of the retrieved studies
were further reviewed to ensure that a comprehensive selection of
the relevant studies was carried out. According to the abstracts
provided by the databases, the original sources of the studies were
located in order to obtain full texts and complete data.
Inclusion criteria and quality
control
All the participanting authors evaluated the quality
of the selected studies and reached a consensus regarding the
appropriate inclusion criteria and quality control. Studies meeting
the following criteria were included in the meta-analysis: 1)
Case-control studies involving the polymorphism rs11136000; 2) LOAD
cases diagnosed according to the internationally accepted
standards, DSM-IV and NINCDS-ADRDA (27), with the controls being local matched
healthy individuals; 3) sample size was explicit, genotype and
allele frequency data could be obtained, original data was provided
directly or could be used to calculate the odds ratio (OR) and 95%
confidence intervals (CI), and information was sufficient to
compare the statistical differences between the groups; 4)
experimental methods for detecting gene polymorphisms were
scientific and rigorous; and 5) allele frequency met the
Hardy-Weinberg equilibrium (HWE). The articles published with
incomplete data were excluded from the meta-analysis. Among
repeatedly reported studies, those with the most complete
information were used.
Statistical analysis
RevMan5.2 version 5.2 (The Nordic Cochrane Centre,
The Cochrane Collaboration, Oxford, UK) and Stata 11.0 software
version 11.0 (StataCorp LP, College Station, TX, USA) were used for
statistical analysis.
HWE test
A χ2 test was used to investigate the HWE
of rs11136000 allele distributions among the selected studies, in
order to verify the representativeness of the study population.
Studies in which the allele frequency of the control group did not
satisfy HWE law were removed.
Heterogeneity test
A Q test was performed to examine the
heterogeneity among the selected studies. RevMan software provided
the P-value of the Q tests, with P<0.10 considered to
indicate a statistically significant difference. Different effect
models for meta-analysis were selected based on the results of the
Q tests.
Selection of statistical models
If there was no statistically significant
heterogeneity among the results of the studies, a Peto
Mantel-Haenszel fixed-effect model was used to determine the
overall OR. If there was statistically significant heterogeneity, a
Dersimonian-Laird random-effect model was used. The RevMan software
provided the overall OR and its 95% CI.
Test for overall effect
Forest plots were provided by the RevMan software,
on which the Z and P-values were shown on the ‘test for
overall effect’, with P<0.05 considered to indicate a
statistically significant difference.
Bias analysis
Funnel plots were generated by the RevMan software
for the preliminary analysis of publication bias. Using the Stata
software, Egger's linear regression and Begg's rank correlation
tests were performed to objectively evaluate publication bias and
to verify the symmetry of the funnel plots, with P<0.10
considered to indicate a statistically significant difference.
Lastly, the fail-safe number (Nfs) was calculated to
further measure publication bias using the following formula:
Nfs0.05 = (ΣZ/1.64)2 - k. The larger the
Nfs value, the smaller the bias.
Results
Characteristics of eligible
studies
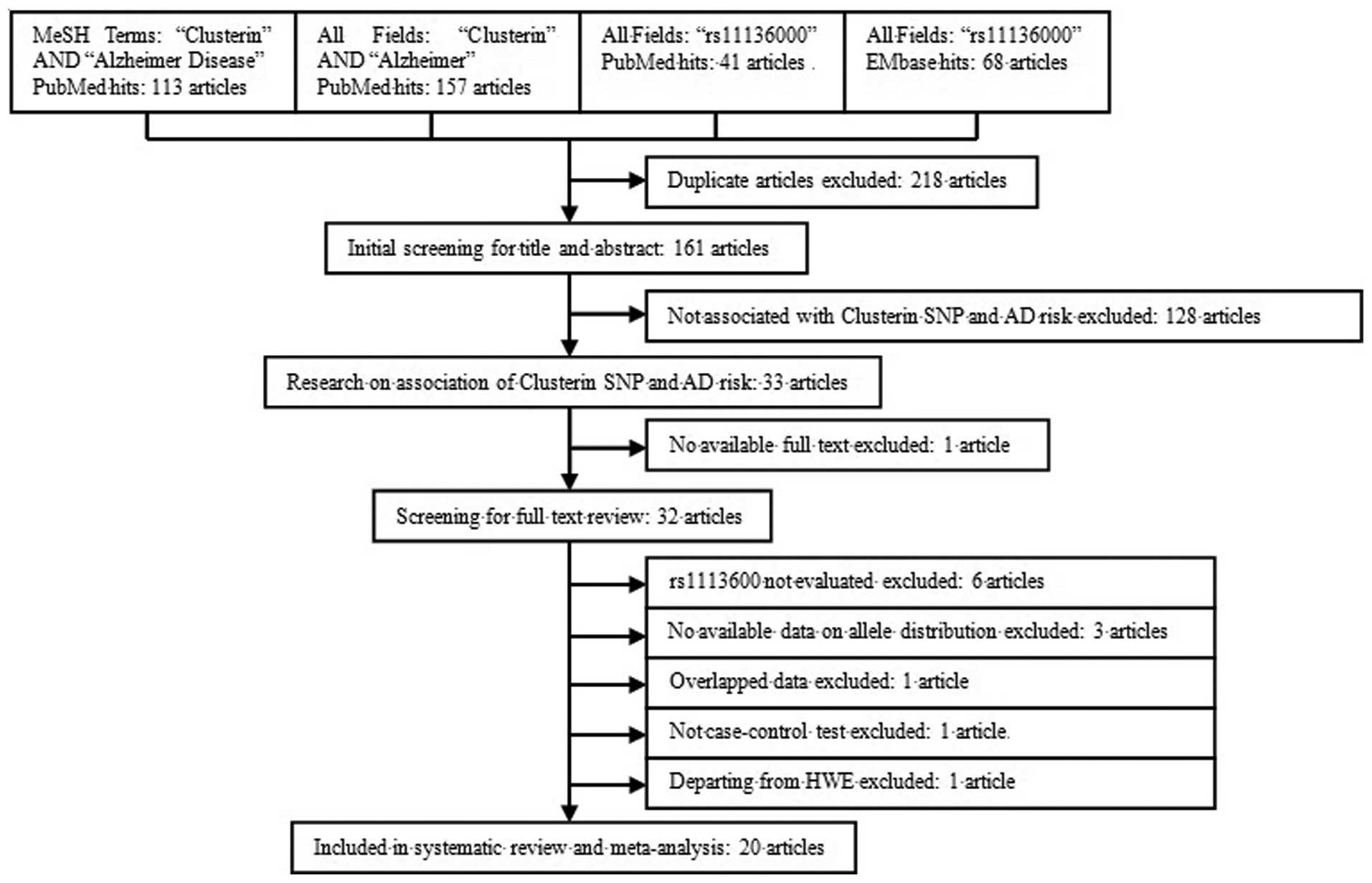
Following a literature review and further screening
according to the inclusion criteria, 20 studies were included in
the meta-analysis (Fig. 1) (6,7,9–26), all
of which were published after 2009 in foreign biomedical journals,
but not reported in Chinese journals. The study population of these
investigations included Caucasian, Hispanic, and Asian populations,
populations of African descent, as well as other ethnicities, of
which 12 were from Europe or America, and 8 were from Asia. A total
of 32,017 individuals with LOAD were included as cases, whose
diagnoses were predominantly based on the clinical criteria, with
only a minority diagnosed based on pathological evidence. A total
of 43,956 individuals were included as controls, in which the
allele distributions were in accordance with the HWE. The
characteristics of the eligible studies are shown in Table I. The genotype and allele frequencies
of the CLU gene polymorphism rs11136000 are shown in
Table II.
 | Table I.Characteristics of the studies
included in the meta-analysis. |
Table I.
Characteristics of the studies
included in the meta-analysis.
|
|
|
| LOAD | Control |
|
|
|---|
|
|
|
|
|
|
|
|
|---|
| Included
studies | Population | Results | Cases (%
female) | Diagnostic
criteria | Average age of
onset | Average age | Case (%
female) | Average age | Genotyping | References |
|---|
| Caucasian |
|
|
|
|
|
|
|
|
|
|
| Lambert
et al, 2009 | Stage 1
(France) | Pos | 2,025 (66%) | C | 68.3±9.0 | 73.7±8.9 | 5,328 (61%) | 73.8±5.4 | Illumina infinium
system | (6) |
|
| Stage 2
(Italy) | Pos | 1,520 (68%) | C | 73.8±8.8 | 76.6±8.7 | 1,291 (55%) | 72.3±8.9 | Illumina infinium
system |
|
|
| Stage 2
(Spain) | Neg | 755 (57%) | C | 72.5±9.4 | 75.3±9.3 | 833 (62%) | 76.9±10.9 | Illumina infinium
system |
|
|
| Stage 2
(Belgium) | Pos | 1,084 (66%) | C | 74.4±8.6 | 78.6±8.1 | 509 (58%) | 67.0±12.9 | Illumina infinium
system |
|
|
| Stage 2
(Finland) | Pos | 619 (67%) | C | 71.4±7.5 | 71.4±7.5 | 664 (60%) | 69.2±6.0 | Illumina infinium
system |
|
| Harold
et al, 2009 | Stage 1 (USA) | Pos | 1,159 (58%) | M | 73.5 | 80.7 | 1,783 (56%) | 68.1 | Illumina infinium
system | (7) |
|
| Stage 1 (UK,
Ireland) | Pos | 2,227 (65%) | M | 72.9 | 79.7 | 5,241 (53%) | 51.2 | Illumina infinium
system |
|
|
| Stage 1
(Germany) | Pos | 555 (64%) | C | 70.5 | 72.9 | 824
(51%) | 56.5 | Illumina infinium
system |
|
|
Giedraitis et al,
2009 | Sweden (ULSAM) | Neg | 86 (0%) | C | 80.2 | – | 404 (0%) | 81.8 | Illumina GoldenGate
assay | (9) |
|
Golenkina et al,
2010 | Russian | Neg | 534 (−) | C | – | – | 702 (−) | – | – | (10) |
| Jun
et al, 2010 | USA (ADGC-C) | Pos | 5,935 (−) | M | – | – | 7,034 (−) | – | Illumina or
affymetrix arrays | (17) |
|
Seshadri et al,
2010 | Spain (ACE) | Pos | 1,140 (70%) | C | – | 78.8±7.9 | 1,209 (53%) | 49.9±9.2 | Illumina or
affymetrix arrays | (24) |
|
Carrasquillo et al,
2010 | USA | Pos | 1,829 (−) | M | – | – | 2,576 (−) | – | TaqMan SNP
genotyping assays | (13) |
|
Schjeide et al,
2011 | Germany | Neg | 214 | C | – | – | 211 (−) | – | OpenArray
genotyping system | (23) |
|
| USA | Pos | 2,654 | M | – | – | 1,175 (−) | – | OpenArray
genotyping system |
|
| Bettens
et al, 2012 | Stage 1
(Belgium) | Pos | 1,057 (66%) | C | 74.9±8.9 | – | 873 (57.4%) | 65.1±14.9 | PCR | (12) |
|
| Stage 2
(France) | Neg | 1,465 (66%) | C | 69.5±8.2 | – | 717 (62.3%) | 74.0±8.0 | PCR |
|
|
| Stage 2
(Canada) | Neg | 323 (55%) | C | 75.3±9.7 | – | 250 (60.0%) | 73.0±10.2 | PCR |
|
| Kamboh
et al, 2012 | USA | Neg | 1,348 (66%) | M | 72.6±6.4 | – | 1,359 (61%) | 74.7±6.5 | TaqMan SNP
genotyping assays | (18) |
|
Carrasquillo et al,
2014 | USA | Pos | 54 (76%) | N | – | – | 2,523 (56.7%) | – | TaqMan SNP
genotyping assays | (14) |
| Asian |
|
|
|
|
|
|
|
|
|
|
| Yu
et al, 2010 | China | Neg | 324 (56%) | C | – | 76.8±5.5 | 388 (54%) | 75.9±4.6 | MALDI-TOF mass
spectrometry | (25) |
| Gu
et al, 2011 | Indiana | Neg | 106 (56%) | C | – | 76.7±7.0 | 98 (55.1%) | 76.1±7.1 | PCR | (11) |
| Ohara
et al, 2012 | Japan | Pos | 824 (77%) | C | – | 83.2±6.5 | 2,933 (56.0%) | 60.2±11.5 | Invader assay | (22) |
| Lin
et al, 2012 | Taiwan | Pos | 268 (−) | C | – | – | 389 (−) | – | – | (19) |
| Chen
et al, 2012 | Hong Kong | Neg | 462 (−) | C | – | – | 350 (−) | – | Sequenom
platform | (15) |
| Ma
et al, 2012 | China | Neg | 127 (58%) | C | – | 73.1±8.6 | 143 (55.2%) | 73.8±6.3 | PCR-RFLP | (20) |
|
Miyashita et al,
2013 | Stage 1
(Japan) | Pos | 1,008 (72%) | C | 73.0±4.3 | – | 1,016 (57%) | 77.0±5.9 | Affymetrix GeneChip
6.0 microarrays | (21) |
|
| Stage 3
(Korean) | Pos | 339 (72%) | C | – | 73.7±9.5 | 1,129 (49%) | 71.0±4.9 | TaqMan assays |
|
| Lu
et al, 2014 | Stage 2
(China) | Neg | 499 (55%) | C | – | 70.0±10.0 | 592 (59.3%) | 68.9±9.4 | PCR-RFLP | (26) |
| African
descent |
|
|
|
|
|
|
|
|
|
|
| Jun
et al, 2010 | USA (ADGC-AA) | Neg | 462 (−) | M | – | – | 449 (−) | – | Illumina
660Quad | (17) |
| Hispanics |
|
|
|
|
|
|
|
|
|
|
| Jun
et al, 2010 | USA (ADGC-H) | Neg | 549 (−) | M | – | – | 544 (−) | – | Illumina HumanHap
650Y chip | (17) |
| Other/Mixed |
|
|
|
|
|
|
|
|
|
|
| Jun
et al, 2010 | USA (ADGC-Wadi
Ara) | Neg | 124 (−) | M | – | – | 142 (−) | – | Illumina
660Quad | (17) |
| Ferrari
et al, 2012 | UK | Pos | 342 (59%) | C | 76.8±8.6 | – | 277 (64.6%) | 70.2±8.6 | TaqMan SNP
genotyping assays | (16) |
| Table II.Distribution of CLU rs11136000
genotypes and alleles among LOAD cases and controls in the included
studies. |
Table II.
Distribution of CLU rs11136000
genotypes and alleles among LOAD cases and controls in the included
studies.
|
| Genotype
distribution (%) | Allele distribution
(%) |
|---|
|
|
|
|
|---|
|
|
| LOAD | Control | LOAD | Control |
|
|
|
|---|
|
|
|
|
|
|
|
|
|
|
|---|
| Included
studies | Studied
population | TT | TC | CC | TT | TC | CC | T | C | T | C | OR | 95% CI | Z-value |
|---|
| Lambert et
al, 2009 (6) | Stage 1
(France) | 270 | 900 | 869 | 860 | 2,561 | 1,957 | 1,440 | 2,638 | 4,281 | 6,475 | 0.83 | 0.77, 0.89 | −5.0122 |
|
|
| (13.2) | (44.1) | (42.6) | (16.0) | (47.6) | (36.4) | (35.3) | (64.7) | (39.8) | (60.2) |
|
|
|
|
| Stage 2
(Italy) | 197 | 682 | 601 | 211 | 570 | 482 | 1,076 | 1,884 | 992 | 1,534 | 0.88 | 0.79, 0.99 | −2.2241 |
|
|
| (13.3) | (46.1) | (40.6) | (16.7) | (45.1) | (38.2) | (36.4) | (63.6) | (39.3) | (60.7) |
|
|
|
|
| Stage 2
(Spain) | 99 | 344 | 305 | 112 | 389 | 309 | 542 | 954 | 613 | 1,007 | 0.93 | 0.81, 1.08 | −0.9293 |
|
|
| (13.2) | (46.0) | (40.8) | (13.8) | (48.0) | (38.1) | (36.2) | (63.8) | (37.8) | (62.2) |
|
|
|
|
| Stage 2
(Belgium) | 155 | 472 | 408 | 79 | 241 | 171 | 782 | 1,288 | 399 | 583 | 0.89 | 0.76, 1.04 | −1.5117 |
|
|
| (15.0) | (45.6) | (39.4) | (16.1) | (49.1) | (34.8) | (37.8) | (62.2) | (40.6) | (59.4) |
|
|
|
|
| Stage 2
(Finland) | 86 | 286 | 224 | 109 | 323 | 218 | 458 | 734 | 541 | 759 | 0.88 | 0.75, 1.03 | −1.6241 |
|
|
| (14.4) | (48.0) | (37.6) | (16.8) | (49.7) | (33.5) | (38.4) | (61.6) | (41.6) | (58.4) |
|
|
|
| Harold et
al, 2009 (7) | Stage 1 (USA) | 163 | 509 | 481 | 328 | 1,085 | 774 | 835 | 1,471 | 1,741 | 2,633 | 0.86 | 0.77, 0.95 | −2.8678 |
|
|
| (14.1) | (44.1) | (41.7) | (15.0) | (49.6) | (35.4) | (36.2) | (63.8) | (39.8) | (60.2) |
|
|
|
|
| Stage 1 | 295 | 1038 | 887 | 787 | 2323 | 1,723 | 1,628 | 2,812 | 3,897 | 5769 | 0.86 | 0.80, 0.92 | −4.1228 |
|
| (UK, Ireland) | (13.3) | (46.8) | (40.0) | (16.3) | (48.1) | (35.7) | (36.7) | (63.3) | (40.3) | (59.7) |
|
|
|
|
| Stage 1
(Germany) | 66 | 240 | 233 | 144 | 368 | 294 | 372 | 706 | 656 | 956 | 0.76 | 0.65, 0.89 | −3.2328 |
|
|
| (12.2) | (44.5) | (43.2) | (17.5) | (46.8) | (35.7) | (34.5) | (65.5) | (40.7) | (59.3) |
|
|
|
| Giedraitis et
al, 2009 (9) | Sweden | 15 | 31 | 33 | 59 | 166 | 140 | 61 | 97 | 284 | 446 | 0.99 | 0.69, 1.41 | −0.0693 |
|
|
| (19.0) | (39.2) | (41.8) | (16.2) | (45.5) | (38.4) | (38.6) | (61.4) | (38.1) | (61.1) |
|
|
|
| Golenkina et
al, 2010 (10) | Russian | 58 | 262 | 214 | 99 | 341 | 262 | 378 | 690 | 539 | 865 | 0.88 | 0.75, 1.04 | −1.5277 |
|
|
| (10.9) | (49.0) | (40.1) | (14.1) | (48.6) | (37.3) | (35.4) | (64.6) | (38.4) | (61.6) |
|
|
|
| Jun et al,
2010 (17) | USA (ADGC-C) | – | – | – | – | – | – | – | – | – | – | 0.91 | 0.85, 0.96 | −3.7532 |
|
| USA (ADGC-AA) | – | – | – | – | – | – | – | – | – | – | 1.06 | 0.89, 1.28 | 0.6172 |
|
| USA (ADGC-H) | – | – | – | – | – | – | – | – | – | – | 1.10 | 0.91, 1.32 | 1.1137 |
|
| USA | – | – | – | – | – | – | – | – | – | – | 0.96 | 0.69, 1.32 | −0.2141 |
|
| (ADGC-Wadi
Ara) |
|
|
|
|
|
|
|
|
|
|
|
|
|
| Seshadri et
al, 2010 (24) | Spain (ACE) | 148 | 525 | 467 | 184 | 575 | 450 | 821 | 1459 | 943 | 1475 | 0.88 | 0.78, 0.99 | −2.1150 |
|
|
| (13.0) | (46.1) | (41.0) | (15.2) | (47.6) | (37.2) | (36.0) | (64.0) | (39.0) | (61.0) |
|
|
|
| Carrasquillo et
al, 2010 (13) | USA | 249 | 848 | 722 | 431 | 1,241 | 893 | 1,346 | 2,292 | 2,103 | 3,027 | 0.85 | 0.77, 0.92 | −3.7725 |
|
|
| (13.7) | (46.6) | (39.7) | (16.8) | (48.4) | (34.8) | (37.0) | (63.0) | (41.0) | (59.0) |
|
|
|
| Yu et al,
2010 (25) | China | 2 | 104 | 218 | 12 | 126 | 250 | 108 | 540 | 150 | 626 | 0.83 | 0.64, 1.10 | −1.2983 |
|
|
| (0.6) | (32.1) | (67.3) | (3.1) | (32.5) | (64.4) | (16.7) | (83.3) | (19.3) | (80.7) |
|
|
|
| Schjeide et
al, 2011 (23) | Germany | – | – | – | – | – | – | – | – | – | – | 0.99 | 0.83, 1.27 | −0.0905 |
|
| USA | – | – | – | – | – | – | – | – | – | – | 0.84 | 0.73, 0.96 | −3.4529 |
| Gu et al,
2011 (11) | Indiana | 4 | 72 | 30 | 6 | 67 | 25 | 80 | 132 | 79 | 117 | 0.90 | 0.60, 1.34 | −0.5318 |
|
|
| (3.8) | (67.9) | (28.3) | (6.1) | (68.4) | (25.5) | (37.7) | (62.3) | (40.3) | (59.7) |
|
|
|
| Ohara et al,
2012 (22) | Japan | 60 | 295 | 469 | 242 | 1,156 | 1,535 | 415 | 1,233 | 1,640 | 4,226 | 0.87 | 0.77, 0.98 | −2.2324 |
|
|
| (7.3) | (35.8) | (56.9) | (8.3) | (39.4) | (52.3) | (25.2) | (74.8) | (28.0) | (72.0) |
|
|
|
| Lin et al,
2012 (19) | Taiwan | 3 | 89 | 176 | 29 | 118 | 242 | 95 | 441 | 176 | 602 | 0.74 | 0.56, 0.97 | −2.1521 |
|
|
| (1.1) | (33.2) | (65.7) | (7.5) | (30.3) | (62.2) | (17.7) | (82.3) | (22.6) | (77.4) |
|
|
|
| Chen et al,
2012 (15) | Hong Kong | 15 | 162 | 274 | 24 | 114 | 200 | 192 | 710 | 162 | 514 | 0.86 | 0.68, 1.09 | −1.2616 |
|
|
| (3.3) | (35.9) | (60.8) | (7.1) | (33.7) | (59.2) | (21.3) | (78.7) | (24.0) | (76.0) |
|
|
|
| Bettens et
al, 2012 (12) | Stage 1
(Belgium) | – | – | – | – | – | – | 676 | 1,232 | 630 | 990 | 0.79 | 0.68, 0.93 | −2.1199 |
|
|
|
|
|
|
|
|
| (35.4) | (64.6) | (38.9) | (61.1) |
|
|
|
|
| Stage 2
(France) | – | – | – | – | – | – | 875 | 1,707 | 452 | 764 | 0.93 | 0.79, 1.10 | −1.9789 |
|
|
|
|
|
|
|
|
| (33.9) | (66.1) | (37.2) | (62.8) |
|
|
|
|
| Stage 2
(Canada) | – | – | – | – | – | – | 236 | 372 | 179 | 299 | 1.00 | 0.77, 1.31 | 0.4606 |
|
|
|
|
|
|
|
|
| (38.8) | (61.2) | (37.4) | (62.6) |
|
|
|
| Kamboh et
al, 2012 (18) | USA | 179 | 623 | 542 | 195 | 636 | 519 | 981 | 1707 | 1,026 | 1,674 | 0.94 | 0.84, 1.05 | −1.1420 |
|
|
| (13.3) | (46.4) | (40.3) | (14.4) | (47.1) | (38.4) | (36.5) | (63.5) | (38.0) | (62.0) |
|
|
|
| Ferrari et
al, 2012 (16) | UK | – | – | – | – | – | – | 254 | 430 | 242 | 312 | 0.76 | 0.60, 0.96 | −2.3357 |
|
|
|
|
|
|
|
|
| (37.1) | (62.9) | (43.7) | (56.3) |
|
|
|
| Ma et al,
2012 (20) | China | 7 | 39 | 81 | 5 | 58 | 80 | 53 | 201 | 68 | 218 | 0.85 | 0.56, 1.27 | −0.8090 |
|
|
| (5.5) | (30.7) | (63.8) | (3.5) | (40.6) | (55.9) | (20.9) | (79.1) | (23.8) | (76.2) |
|
|
|
| Miyashita et
al, 2013 (21) | Stage 1
(Japan) | – | – | – | – | – | – | – | – | – | – | 0.87 | 0.78, 0.97 | −2.0100 |
|
| Stage 3
(Korean) | – | – | – | – | – | – | – | – | – | – | 0.79 | 0.63, 0.98 | −2.3006 |
| Carrasquillo et
al, 2014 (14) | USA | 4 | 25 | 25 | 416 | 1,165 | 843 | 33 | 75 | 1,997 | 2,851 | 0.60 | 0.39, 0.92 | −2.2044 |
|
|
| (7.4) | (46.3) | (46.3) | (17.2) | (48.0) | (34.8) | (30.6) | (69.4) | (41.2) | (58.8) |
|
|
|
| Lu et al,
2014 (26) | Stage 2
(China) | 18 | 156 | 319 | 23 | 161 | 399 | 192 | 794 | 207 | 959 | 1.12 | 0.90, 1.39 | 1.0224 |
|
|
| (3.7) | (31.6) | (64.7) | (4.0) | (27.6) | (68.4) | (19.5) | (80.5) | (17.8) | (82.2) |
|
|
|
Considering ethnic differences,
subgroup meta-analyses were conducted according to five ethnicities
(Caucasian, Asian, African descent, Hispanic and other/mixed
ethnicities)
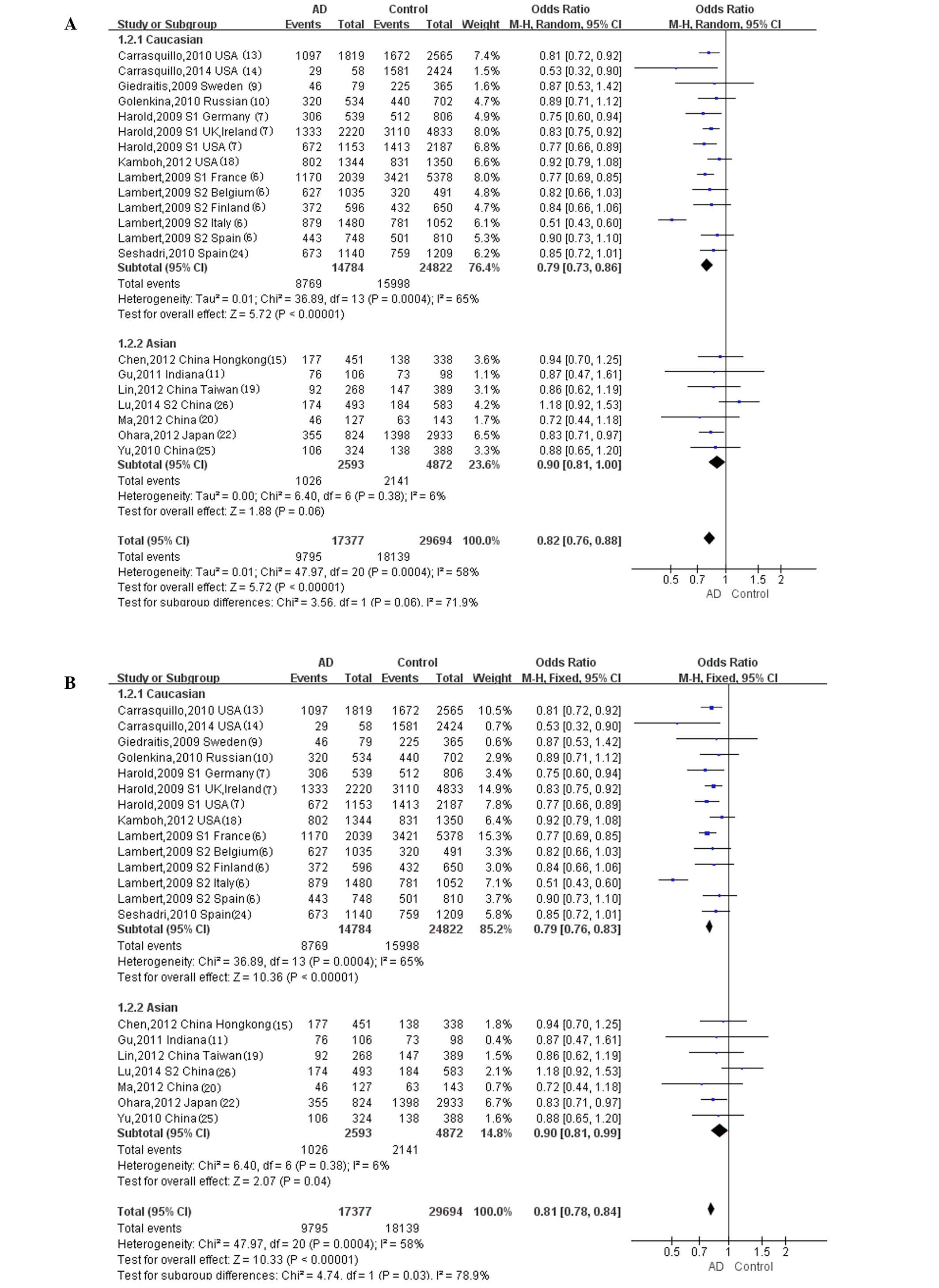
Using the (TT+TC)/CC genotype model (Fig. 2), a significant heterogeneity was
observed in the Caucasian population with P=0.0004, although no
significant heterogeneity was observed in the Asian population with
P=0.38. Using the T/C allele model (Fig.
3), statistically significant heterogeneity was not observed in
the Caucasian population with P=0.57, nor in the Asian (P=0.47) or
other/mixed ethnicity populations (P=0.27). No heterogeneity tests
were required in the ethnic subgroups in which only one study was
eligible.
Meta-analysis
The random-effect model was used to test for the
overall effect of the (TT+TC)/CC genotype model in the Caucasian
population (Fig. 2A), in which a
statistically significant difference was observed (overall OR,
0.79; 95% CI, 0.73–0.86; P<0.00001). The fixed-effect model was
used in the Asian population for the (TT+TC)/CC genotype model
(Fig. 2B), and a statistically
significant difference was also observed (overall OR, 0.90; 95% CI,
0.81–0.99; P=0.04). The fixed-effect model was used in the
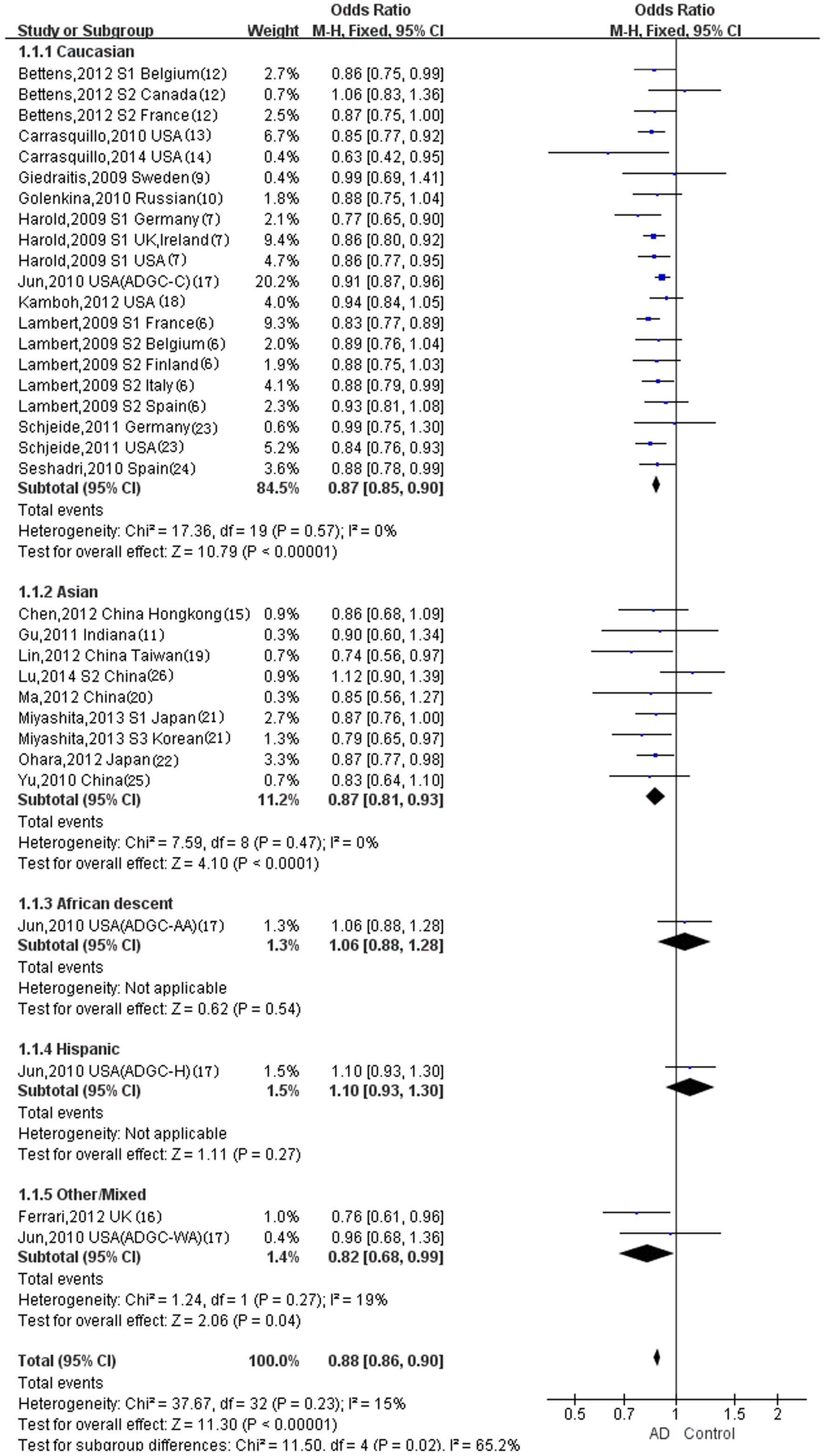
Caucasian, Asian and other/mixed ethnicity populations for the T/C
allele model (Fig. 3), and
statistically significant differences were observed for all
ethnicities (Caucasian: Overall OR, 0.87; 95% CI, 0.85–0.90;
P<0.00001; Asian: Overall OR, 0.87; 95% CI, 0.81–0.93;
P<0.0001; other/mixed ethnicities: Overall OR, 0.82; 95% CI,
0.68–0.99; P=0.04). Statistically significant differences were not
however observed in the populations of African descent (overall OR,
1.06; 95% CI, 0.88–1.28; P=0.54) or Hispanics (overall OR, 1.10;
95% CI, 0.93–1.30; P=0.27) for the T/C allele model (Fig. 3).
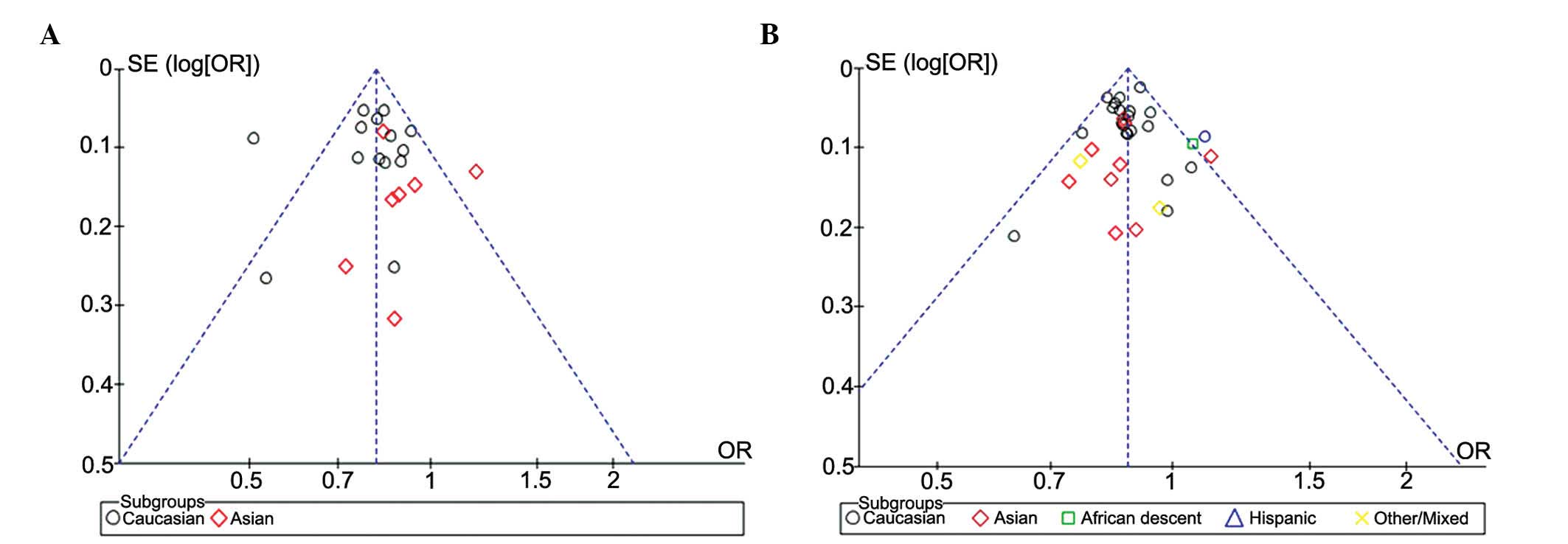
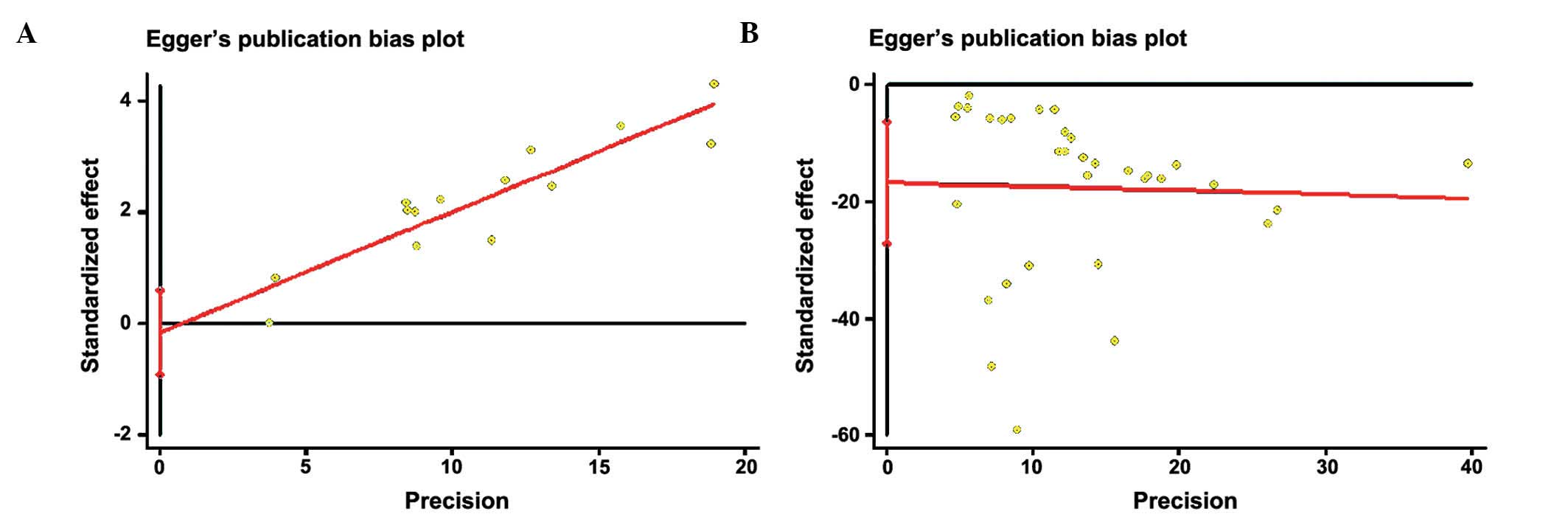
Evaluation of publication bias
The RevMan funnel plot for the genotype model
exhibited relative symmetry, whereas the allele model exhibited
incomplete symmetry (Fig. 4).
Following Egger's linear regression analysis and Begg's rank
correlation test, it was demonstrated that the studies on genotype
had no statistically significant publication bias (bias factors,
−0.156; P=0.658 of Egger's; P=0.584 of Begg's), whereas those on
alleles did have some bias (bias factors, −16.716; P=0.002 of
Egger's; P=0.009 of Begg's) (Table
III and Fig. 5).
Nfs0.05 was equal to 750.604, which was large enough to
warrant ignoring the publication bias.
| Table III.Results of Egger's linear regression
analysis and Begg's rank correlation test for publication bias. |
Table III.
Results of Egger's linear regression
analysis and Begg's rank correlation test for publication bias.
|
| Egger's test | Begg's test |
|---|
|
|
|
|
|---|
|
| Std-Eff | Coef.a | SE | t | P>|t| | 95% CI | Adj. Kendall's
score (P-Q) | SD of score | Number of
studies | Z-value | Pr>|z| |
|---|
| Genotype | Slope | 0.2160835 | 0.0287501 | 7.52 | 0.000 | 0.1534424,
0.2787246 | −11 | 18.27 | 14 | 0.55 | 0.584 |
|
| bias | −0.1556121 | 0.3430764 | −0.45 | 0.658 | −0.9031114,
0.5918871 |
|
|
|
|
|
| Allele | Slope | −0.0658025 | 0.3326355 | −0.20 | 0.844 | −0.744217,
0.0612612 | −170 | 64.54 | 33 | 2.62 | 0.009 |
|
| bias | −16.71626 | 5.075586 | −3.29 | 0.002 | −27.06799,
−6.364535 |
|
|
|
|
|
Discussion
The CLU gene is located on p21-p12 of human
chromosome 8, with CLU as its encoded product, which has various
physiological functions, including participating in lipid
metabolism (28), oxidative stress
reaction (29), and cell cycle
regulation (30). CLU is
highly expressed in cerebrospinal fluid and amyloid plaques in
brain tissues, and is involved in the pathogenesis of AD (4,5,31). Yerbury et al (32) demonstrated that the deposition of CLU
in senile plaques and neurofibrillary tangles of AD. Howlett et
al (33) further reported a
correlation between CLU and senile plaque Aβ40 in the brain cortex
of patients with AD. Martin-Rehrmann et al (34) demonstrated the presence of
dysfunctional neurons with phosphorylated tau protein surrounding
the senile plaques in 71% of CLU-positive patients with AD.
Furthermore, they also showed that the tau and phosphorylated tau
protein were significantly increased in the rat hippocampus,
following the injection of a CLU-rich solution (34). It was suggested that CLU may be
involved in the formation of Aβ and dysfunctional neurons in the
pathological state of AD (5). Wahrle
et al (35) reported that CLU
regulated the expression and metabolism of APOE in the brain.
Indeed, the levels of cholesterol and APOE were decreased in the
mouse following CLU gene knocked out. Aβ and clusterin
combine together in normal cerebrospinal fluid, which suggested
that CLU may be involved in the metabolism, transportation and
clearance of Aβ in the brain (30).
Bell et al (36) demonstrated
that the clearance of Aβ42 through the blood brain barrier was
increased by 83% when it was combined with CLU. Thambisetty et
al (37) demonstrated the
important role of CLU in the pathogenesis of AD. They showed that
CLU was associated with atrophy of the entorhinal cortex and rapid
clinical progression of AD (37).
The higher the plasma concentration levels of CLU, the more
significant the entorhinal cortex atrophy, and the more severe the
cognitive impairment (37). It was
also demonstrated that increased plasma concentration levels of CLU
was predictive of greater Aβ burden in the medial temporal lobe
(31). In view of the above results,
it was hypothesized that gene mutations or specific environmental
factors could induce changes in CLU gene expression or
function, which have an important role in the pathogenesis of
AD.
Since October 2009, association studies between the
CLU gene polymorphism and LOAD susceptibility have rapidly
progressed worldwide, and have been conducted in numerous
populations, including Caucasian, Asian, African descent, and
Hispanic populations (6,7,9–26). Notably, SNP rs11136000 in the
CLU gene was demonstrated to be significantly associated
with LOAD by two GWAS with large sample sizes and two stages, of
which one was conducted by Lambert et al (6) (OR, 0.86; 95% CI, 0.81–0.90;
P=7.5×10−9) and the other by Harold et al
(7) (OR, 0.86; 95% CI, 0.82–0.90;
P=8.5×10−10). The meta-analysis carried out by Jones
et al (38) also supported
the above conclusion (P=1.72 × 10−15). From then on,
numerous other studies obtained concordant or non-concordant
results (9–26). An issue is that many of the studies
differ in their selection of ethnic genetic backgrounds and sample
sizes, which leads to inconsistent or contrary conclusions
regarding the association between gene polymorphism and diseases.
The present study performed a meta-analysis to systematically
review the published articles regarding the association between the
CLU polymorphism rs11136000 and LOAD susceptibility, with
increased sample size to improve and enhance test efficiency and
ethnic subgroups to eliminate possible ethnic differences. This
investigation aimed to provide objective results of evidence-based
medicine for the association between CLU gene polymorphisms
and AD susceptibility. The results of the present study
demonstrated a positive association between CLU gene
polymorphisms and LOAD susceptibility, as expected.
In the present study, 20 studies of interest were
included in a meta-analysis, covering 33 independent study
populations from various regions, ethnicities and stages, all
meeting the predetermined inclusion criteria. The results
demonstrated that there were some differences in the association
between rs11136000 and LOAD among the various ethnicities.
Meta-analysis supported the following conclusions: 1) TT+TC
genotypes and the T allele of rs11136000 in Caucasian populations
are associated with LOAD, as for the overall-effect test P<0.01;
2) TT+TC genotypes and the T allele of rs11136000 in Asian
populations is associated with LOAD, as for the overall-effect test
both P<0.05; 3) the T allele of rs11136000 in the population of
African descent and the Hispanic population was not associated with
LOAD, as for the single tests P>0.05; and 4) the T allele of
rs11136000 in the other/mixed ethnicity group was associated with
LOAD, as for the overall-effect test P<0.05. To summarize, the
meta-analysis demonstrated that TT+TC genotypes and (or) the T
allele of rs11136000 in Caucasian, Asian, and other/mixed ethnicity
populations were associated with LOAD (P<0.05), and the
population carrying the T allele would likely suffer lower LOAD
risk (OR<1). The results therefore demonstrated that the T
allele of rs11136000 had a protective effect on LOAD. The results
also demonstrated the impact of racial differences on association
studies of gene polymorphisms and disease.
In addition to ethnic differences, sample size also
had an important effect on the study results. In the present
meta-analysis, there was only one study population of African
descent or Hispanic population included (17), with a relatively small sample size,
which may not have provided accurate results. For instance, in the
studies conducted in the Chinese Han population, Yu et al
(25), Chen et al (15) and Ma et al (20) did not report any statistical
significance in the T allele distribution of rs11136000 (P=0.21,
0.22, 0.42, respectively). However, when sample size was enlarged
through meta-analysis and the three studies were included, a weak
association was revealed (P=0.048 in the overall effect test). The
majority of the studies and those with the largest sample size were
conducted in the Caucasian population in the current meta-analysis,
and therefore the results obtained for the Caucasian population are
the most reliable. Further studies are required in other ethnic
groups with larger sample sizes in order to improve the reliability
of the study findings.
It is not only rs11136000 but also other
polymorphisms of the CLU gene may contribute to LOAD
susceptibility. For instance, the study carried out by Yu et
al (25) in the Chinese Han
population did not find an association between rs11136000 and LOAD,
although the results indicated instead an association between
rs9331888 and LOAD, with OR=1.39 (95% CI, 1.13–1.72) and P=0.002.
Other studies also demonstrated the association between
polymorphisms of rs9331888 (6,11,25),
rs2279590 (6,15,21,23),
rs7982 (7,17), rs7012010 (7,17),
rs9331908 (12), rs1532278 (12,39), and
rs9331949 (40) and LOAD, although
these studies were few and inconsistent in their results. The
rs11136000 polymorphism systematically reviewed in the present
meta-analysis is likely one of the factors that influences LOAD
susceptibility. The precise forms of the genetic variants require
further investigation. The meta-analysis only involved rs11136000,
the most well-established CLU gene polymorphism, but was not
representative of the entire CLU gene. The results of the
meta-analysis supported that the CLU gene polymorphism was
associated with LOAD susceptibility in Caucasian and Asian
populations, but there was no such definitive association in
populations of African descent or Hispanic populations.
In conclusion, although the studies included in the
meta-analysis of the present investigation were less than those
conducted on the APOE gene, all were published in authorized
journals after 2009, which indicated that the research on the
CLU gene polymorphism was novel and promising. It is likely
that the CLU polymorphism will be the subject of numerous
studies on the genetic susceptibility to AD in the future. Further
investigations are required in various ethnic population. The
present meta-analysis will be regularly updated in order to draw
more scientific and reliable conclusions regarding the association
between the CLU gene polymorphism and susceptibility to
LOAD.
There are clinical significances and wider
implications of the findings in the present study. For example,
there may be a use in monitoring disease progression in the elderly
at risk. Furthermore, understanding the influence of the LOAD risk
variants on cognitive decline may provide additional information
with regard to their plausible mechanism of action.
References
|
1
|
Ossenkoppele R, van der Flier WM, Zwan MD,
Adriaanse SF, Boellaard R, Windhorst AD, Barkhof F, Lammertsma AA,
Scheltens P and van Berckel BN: Differential effect of APOE
genotype on amyloid load and glucose metabolism in AD dementia.
Neurology. 80:359–365. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tycko B, Feng L, Nguyen L, Francis A, Hays
A, Chung WY, Tang MX, Stern Y, Sahota A, Hendrie H and Mayeux R:
Polymorphisms in the human apolipoprotein-J/clusterin gene: Ethnic
variation and distribution in Alzheimer's disease. Hum Genet.
98:430–436. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bertram L and Tanzi RE: Alzheimer's
disease: New light on an old CLU. Nat Rev Neurol. 6:11–13. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Malkki H: Alzheimer disease: Chaperone
protein clusterin is involved in amyloid-β-associated entorhinal
atrophy in early AD. Nat Rev Neurol. 10:602014. View Article : Google Scholar
|
|
5
|
Nuutinen T, Suuronen T, Kauppinen A and
Salminen A: Clusterin: A forgotten player in Alzheimer's disease.
Brain Res Rev. 61:89–104. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lambert JC, Heath S, Even G, Campion D,
Sleegers K, Hiltunen M, Combarros O, Zelenika D, Bullido MJ,
Tavernier B, et al: Genome-wide association study identifies
variants at CLU and CR1 associated with Alzheimer's disease. Nat
Genet. 41:1094–1099. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Harold D, Abraham R, Hollingworth P, Sims
R, Gerrish A, Hamshere ML, Pahwa JS, Moskvina V, Dowzell K,
Williams A, et al: Genome-wide association study identifies
variants at CLU and PICALM associated with Alzheimer's disease. Nat
Genet. 41:1088–1093. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Butler AW, Ng MY, Hamshere ML, Forabosco
P, Wroe R, Al-Chalabi A, Lewis CM and Powell JF: Meta-analysis of
linkage studies for Alzheimer's disease-a web resource. Neurobiol
Aging. 30:1037–1047. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Giedraitis V, Kilander L,
Degerman-Gunnarsson M, Sundelöf J, Axelsson T, Syvänen AC, Lannfelt
L and Glaser A: Genetic analysis of Alzheimer's disease in the
Uppsala Longitudinal Study of Adult Men. Dement Geriatr Cogn
Disord. 27:59–68. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Golenkina SA, Gol'tsov AIu, Kuznetsova IL,
Grigorenko AP, Andreeva TV, Reshetov DA, Kunizheva SS, Shagam LI,
Morozova IIu, Goldenkova-Pavlova IV, et al: Analysis of clustering
gene (CLU/APOJ) polymorphism in Alzheimer's disease patients and in
normal cohorts from Russian populations. Mol Biol (Mosk).
44:620–626. 2010.(In Russian). View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gu H, Wei X, Chen S, Kurz A, Müller U,
Gasser T, Dodel RC, Farlow MR and Du Y: Association of clusterin
gene polymorphisms with late-onset Alzheimer's disease. Dement
Geriatr Cogn Disord. 32:198–201. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bettens K, Brouwers N, Engelborghs S,
Lambert JC, Rogaeva E, Vandenberghe R, Le Bastard N, Pasquier F,
Vermeulen S, Van Dongen J, et al: Both common variations and rare
non-synonymous substitutions and small insertion/deletions in CLU
are associated with increased AD risk. Mol Neurodegener. 7:32012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Carrasquillo MM, Belbin O, Hunter TA, Ma
L, Bisceglio GD, Zou F, Crook JE, Pankratz VS, Dickson DW,
Graff-Radford NR, et al: Replication of CLU, CR1 and PICALM
associations with alzheimer's disease. Arch Neurol. 67:961–964.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carrasquillo MM, Khan Qu, Murray ME,
Krishnan S, Aakre J, Pankratz VS, Nguyen T, Ma L, Bisceglio G,
Petersen RC, et al: Late-onset Alzheimer disease genetic variants
in posterior cortical atrophy and posterior AD. Neurology.
82:1455–1462. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen LH, Kao PY, Fan YH, Ho DT, Chan CS,
Yik PY, Ha JC, Chu LW and Song YQ: Polymorphisms of CR1, CLU and
PICALM confer susceptibility of Alzheimer's disease in a southern
Chinese population. Neurobiol Aging. 33:210.e1–210.e7. 2012.
View Article : Google Scholar
|
|
16
|
Ferrari R, Moreno JH, Minhajuddin AT,
O'Bryant SE, Reisch JS, Barber RC and Momeni P: Implication of
common and disease specific variants in CLU, CR1 and PICALM.
Neurobiol Aging. 33:1846.e7–1846.e18. 2012. View Article : Google Scholar
|
|
17
|
Jun G, Naj AC, Beecham GW, Wang LS, Buros
J, Gallins PJ, Buxbaum JD, Ertekin-Taner N, Fallin MD, Friedland R,
et al: Meta-analysis confirms CR1, CLU and PICALM as alzheimer
disease risk loci and reveals interactions with APOE genotypes.
Arch Neurol. 67:1473–1484. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kamboh MI, Minster RL, Demirci FY, Ganguli
M, Dekosky ST, Lopez OL and Barmada MM: Association of CLU and
PICALM variants with Alzheimer's disease. Neurobiol Aging.
33:518–521. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin YL, Chen SY, Lai LC, Chen JH, Yang SY,
Huang YL, Chen TF, Sun Y, Wen LL, Yip PK, et al: Genetic
polymorphisms of clusterin gene are associated with a decreased
risk of Alzheimer's disease. Eur J Epidemiol. 27:73–75. 2012.
View Article : Google Scholar
|
|
20
|
Ma JF, Liu LH, Zhang Y, Wang Y, Deng YL,
Huang Y, Wang G, Xu W, Cui PJ, Fei QZ, et al: Association study of
clusterin polymorphism rs11136000 with late onset Alzheimer's
disease in Chinese Han population. Am J Alzheimers Dis Other Demen.
26:627–630. 2012. View Article : Google Scholar
|
|
21
|
Miyashita A, Koike A, Jun G, Wang LS,
Takahashi S, Matsubara E, Kawarabayashi T, Shoji M, Tomita N, Arai
H, et al: SORL1 is genetically associated with late-onset
Alzheimer's disease in Japanese, Koreans and Caucasians. PLoS One.
8:e586182013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ohara T, Ninomiya T, Hirakawa Y, Ashikawa
K, Monji A, Kiyohara Y, Kanba S and Kubo M: Association study of
susceptibility genes for late-onset Alzheimer's disease in the
Japanese population. Psychiatr Genet. 22:290–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schjeide BM, Schnack C, Lambert JC, Lill
CM, Kirchheiner J, Tumani H, Otto M, Tanzi RE, Lehrach H, Amouyel
P, et al: The role of clusterin, complement receptor 1 and
phosphatidylinositol binding clathrin assembly protein in Alzheimer
disease risk and cerebrospinal fluid biomarker levels. Arch Gen
Psychiatry. 68:207–213. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Seshadri S, Fitzpatrick AL, Ikram MA,
DeStefano AL, Gudnason V, Boada M, Bis JC, Smith AV, Carassquillo
MM, Lambert JC, et al: Genome-wide analysis of genetic loci
associated with Alzheimer disease. JAMA. 303:1832–1840. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu JT, Li L, Zhu QX, Zhang Q, Zhang W, Wu
ZC, Guan J and Tan L: Implication of CLU gene polymorphisms in
Chinese patients with Alzheimer's disease. Clin Chim Acta.
411:1516–1519. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lu SJ, Li HL, Sun YM, Liu ZJ, Yang P and
Wu ZY: Clusterin variants are not associated with southern Chinese
patients with Alzheimer's disease. Neurobiol Aging.
35:2656.e9–2656.e11. 2014. View Article : Google Scholar
|
|
27
|
McKhann G, Drachman D, Folstein M, Katzman
R, Price D and Stadlan EM: Clinical diagnosis of Alzheimer's
disease: report of the NINCDS-ADRDA work group under the auspices
of Department of Health and Human Services Task Force on
Alzheimer's Disease. Neurology. 1984.34:939–944. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Leeb C, Eresheim C and Nimpf J: Clusterin
is a ligand for apolipoprotein E receptor 2 (ApoER2) and very low
density lipoprotein receptor (VLDLR) and signals via the
Reelin-signaling pathway. J Biol Chem. 289:4161–4172. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Trougakos IP: The molecular chaperone
apolipoprotein J/clusterin as a sensor of oxidative stress:
Implications in therapeutic approaches-a mini-review. Gerontology.
59:514–523. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Song HB, Jun HO and Kim JH, Yu YS, Kim KW,
Min BH and Kim JH: Anti-apoptotic effect of clusterin on
cisplatin-induced cell death of retinoblastoma cells. Oncol Rep.
30:2713–2718. 2013.PubMed/NCBI
|
|
31
|
Li X, Ma Y, Wei X, Li Y, Wu H, Zhuang J
and Zhao Z: Clusterin in Alzheimer's disease: A player in the
biological behavior of amyloid-beta. Neurosci Bull. 30:162–168.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yerbury JJ, Poon S, Meehan S, Thompson B,
Kumita JR, Dobson CM and Wilson MR: The extracellular chaperone
clusterin influences amyloid formation and toxicity by interacting
with prefibrillar structures. FASEB J. 21:2312–2322. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Howlett DR, Hortobágyi T and Francis PT:
Clusterin associates specifically with Aβ40 in Alzheimer's disease
brain tissue. Brain Pathol. 23:623–632. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Martin-Rehrmann MD, Hoe HS, Capuani EM and
Rebeck GW: Association of apolipoprotein J-positive beta-amyloid
plaques with dystrophic neurites in Alzheimer's disease brain.
Neurotox Res. 7:231–242. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wahrle SE, Jiang H, Parsadanian M,
Legleiter J, Han X, Fryer JD, Kowalewski T and Holtzman DM: ABCA1
is required for normal central nervous system ApoE levels and for
lipidation of astrocyte-secreted ApoE. J Biol Chem.
279:40987–40993. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bell RD, Sagare AP, Friedman AE, Bedi GS,
Holtzman DM, Deane R and Zlokovic BV: Transport pathways for
clearance of human Alzheimer's amyloid beta-peptide and
apolipoproteins E and J in the mouse central nervous system. J
Cereb Blood Flow Metab. 27:909–918. 2007.PubMed/NCBI
|
|
37
|
Thambisetty M, Simmons A, Velayudhan L,
Hye A, Campbell J, Zhang Y, Wahlund LO, Westman E, Kinsey A,
Güntert A, et al: Association of plasma clusterin concentration
with severity, pathology and progression in Alzheimer disease. Arch
Gen Psychiatry. 67:739–748. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jones L, Harold D and Williams J: Genetic
evidence for the involvement of lipid metabolism in Alzheimer's
disease. Biochim Biophys Acta. 1801:754–761. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Naj AC, Jun G, Beecham GW, Wang LS,
Vardarajan BN, Buros J, Gallins PJ, Buxbaum JD, Jarvik GP, Crane
PK, et al: Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1
are associated with late-onset Alzheimer's disease. Nat Genet.
43:436–441. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yu JT, Ma XY, Wang YL, Sun L and Tan L, Hu
N and Tan L: Genetic variation in Clusterin gene and Alzheimer's
disease risk in Han Chinese. Neurobiol Aging. 34:1921.e17–1921.e23.
2013. View Article : Google Scholar
|