Introduction
Triple-negative breast cancer (TNBC) is one breast
cancer subtype that is characterized by the lack expression of
estrogen receptor (ER), progesterone receptor (PR), and human
epidermal growth factor receptor 2 (HER2) (1,2). TNBC
accounts for 9–21% of all breast cancers and has higher metastatic
and recurrence rates and a poorer prognosis than other breast
cancer subtypes (2–5). Currently, chemotherapy is the only
option for treating malignant breast cancer because of the lack of
targeted molecules for disease treatment (6,7). Among
the chemotherapy drugs, platinum agents have received much
attention in the treatment of TNBC (8). However, the growing chemotherapy
resistance and side effect problems restrict its clinical
application (9,10). Therefore, exploring the underlying
mechanisms of drug resistance and seeking a new target for TNBC
chemotherapy is needed.
TG-interacting factor (TGIF), a member of the
three-amino-acid loop extension (TALE) subfamily of atypical
homeodomain proteins, is involved in multiple signaling pathways
(11–17). Since TGIF was identified to be
involved in holoprosencephaly development (18), it has been implicated in diverse
biological and pathological functions. Recently, numerous studies
have focused on the potential roles of TGIF in tumor development,
invasion and migration in various types of cancers, including lung
cancer (19–21), liver cancer (22,23),
ovarian cancer (24), leukemia
(25), gastric carcinoma (26), upper urinary tract urothelial
carcinoma (27) and esophageal
squamous cell carcinoma (28).
However, research regarding the correlation of TGIF protein with
TNBC is limited. Prof. Atfi's laboratory revealed that TGIF
expression was much higher in TNBC patients compared with normal
tissue or other breast cancer subtypes. Moreover, the elevated
levels of TGIF correlated with a high relapse and mortality rate of
TNBC patients (29,30). Kwon et al reported that
targeted interference of SIN3A-TGIF function by SID decoy treatment
inhibited Wnt signaling and invasion in TNBC cells (31). Together, previous papers suggested
that TGIF protein might be a target for TNBC chemotherapy. To the
best of our knowledge, there is no publication focusing on the
relationship between TGIF silencing and cisplatin-induced apoptotic
sensitivity in TNBC cells. In the present study, we used the
representative TNBC cell line of MDA-MB-231 to observe the effects
of TGIF silencing on cisplatin-induced apoptosis.
Materials and methods
Cell culture and cell
transfection
The MDA-MB-231 cell line was maintained in our
laboratory and cultured in DMEM supplemented with penicillin (100
U/ml), streptomycin (100 µg/ml), 10% fetal bovine serum (FBS), and
2 mM L-glutamine in a humidified atmosphere that contained 5%
CO2 at 37°C. TGIF shRNA human (h) lentiviral particles
(sc-36659-V) and control shRNA lentiviral particles-A (sc-108080)
were obtained from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA). The shRNA lentiviral particles against TGIF were infected
into MDA-MB-231 cells according to the manufacturer's instructions.
Next, the stable clones expressing shRNA were initially selected by
10 µg/ml of puromycin for three weeks (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The TGIF expression level of
the infected cells was detected by western blot to confirm the
transfection efficiency. Cells that were stably transfected with
the TGIF shRNA (h) lentiviral particles and control shRNA
lentiviral particles were named MDA-MB-231-shRNA-TGIF cells and
MDA-MB-231-shRNA-control cells, respectively.
Western blot analysis
Cell lysates were prepared in a RIPA buffer (Pierce;
Thermo Fisher Scientific, Inc.), and a BCA protein assay (Pierce;
Thermo Fisher Scientific, Inc.) was conducted to quantify the
protein concentration. The samples were then separated on a 10%
sodium dodecyl sulfate-polyacrylamide electrophoresis gel
(SDS-PAGE) and proteins were transferred onto a nitrocellulose (NC)
membrane. After blocking with 5% bovine serum albumin
(BSA)/Tris-buffered saline Tween-20 (TBST) for 1 h, the membrane
was incubated with primary antibody overnight at 4°C, followed by
adsorption to peroxidase-coupled protein G (ZSGB-BIO, Beijing,
China) for 1 h at room temperature. Antibodies against TGIF
(sc-9084) and p21 (sc-397) were purchased from Santa Cruz
Biotechnology, Inc., and antibodies against PARP (no. 9532S), Bax
(no. 2772S), caspase-3 (no. 9665S) and caspase-9 (no. 9508S) were
obtained from Cell Signaling Technology. Immunoreactive bands were
visualized with a Bio-Rad Clarity™ western ECL substrate
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Antibody to
β-actin (sc-47778; Santa Cruz Biotechnology, Inc.) was used as a
loading control.
MTT assay
Cell viability was determined by MTT assay. We
collected the MDA-MB-231-shRNA-TGIF cells and
MDA-MB-231-shRNA-control cells at a density of 5×104/ml
and then plated cells in 96-well plates at a density of
5×103 cells per well (6-well per group). After
incubation in culture medium for 24 h, the culture medium was
replaced with the following concentrations of cisplatin at 0, 2.5,
5.0, 7.5 and 12.5 µg/ml and maintained for 48 h. Four h before the
cisplatin treatment finished, 10 µl of 5 mg/ml MTT were added to
each well. Then, 150 µl of DMSO were added to each well and the
absorbance was determined on a micro-plate reader (Multiskan
Ascent;. Thermo Labsystems; Thermo Fisher Scientific, Inc.) at 492
nm.
Annexin V and dead cell assay
Annexin V and dead cell assay was used to determine
the cell populations in the apoptosis stage, including the early
apoptotic cells and the late apoptotic cells. For the induction of
apoptosis, cells were seeded in 60-mm plates and cultured for 24 h
at 37°C and then incubated for 48 h with cisplatin (12.5 µg/ml).
Cells were trypsinized and resuspended in at least 1% FBS. The cell
samples were incubated with Muse™ annexin V and dead
cell reagent for 20 min at room temperature in the dark. The
apoptosis rate was measured by the Muse™ Cell Analyzer
with the Muse™ annexin V and dead cell software
module.
Cell apoptosis analysis by hoechst
33258
The Hoechst staining kit (Beyotime Institute of
Biotechnology, Haimen, China) was used to detect the state of
nucleus condensation. Cells were seeded onto cover slides in 6-well
plates overnight at 37°C and then treated with 12.5 µg/ml of
cisplatin for 48 h followed by adding 0.5 ml of fixation fluid for
30 min. After washing with phosphate-buffered saline (PBS) twice,
0.5 ml of Hoechst 33258 were added to the plate and incubated for 5
min. The stained cells were washed with PBS twice and then the
nuclear morphology was observed under a fluorescence microscope
(Zeiss AG, Oberkochen, Germany). The apoptotic cells showed
condensed and fragmented nuclei.
Colony formation assay
A total of 400 cells were plated in 60-mm plates.
After plating for 72 h, cells were treated with 2.5 µg/ml of
cisplatin for 4 h and washed with HANKs three times. Cells were
maintained in an incubator of 5% CO2 at 37°C for 18
days. The culture medium was changed every three days. At the end
of the experiments, cells were washed with PBS and fixed with
methanol for 30 min at room temperature and then stained with
Giemsa for 30 min at room temperature. Colonies with more than 50
cells were counted under an inverted microscope (Leica Microsystems
GmbH, Wetzlar, Germany).
Statistical analysis
SPSS 13.0 software (SPSS, Inc., Chicago, IL, USA)
was used for statistical analyses. All data were expressed as the
mean ± standard deviation (SD). A Student's t test was applied to
evaluate the difference of relative cell viability between groups
at each dose of cisplatin treatment (0, 2.5, 7.5 and 12.5 µg/ml)
and one-way ANOVA (LSD for post-hoc test) was applied to evaluate
the difference among groups in additional experiments. P<0.05
was considered to indicate a statistically significant
difference.
Results
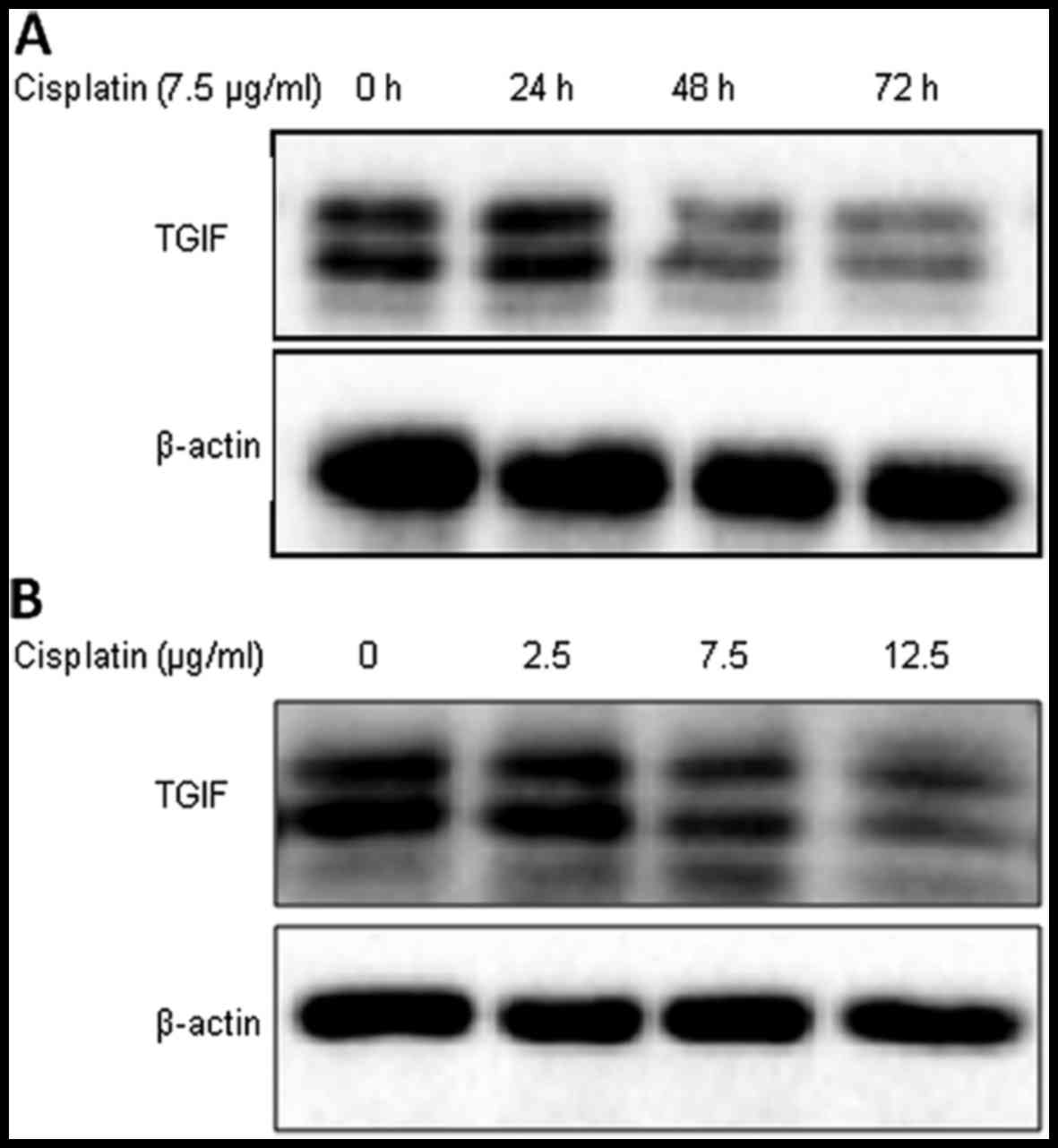
Cisplatin decreased the expression of
TGIF protein in MDA-MB-231 cells
As shown in Fig. 1,
the expression of TGIF protein was suppressed by cisplatin
treatment from 48 h in MDA-MB-231 cells (Fig. 1A). We also tested the level of TGIF
protein expression under different concentrations of cisplatin;
Fig. 1B shows that cisplatin
treatment suppressed TGIF protein expression in a dose-dependent
manner in MDA-MB-231 cells.
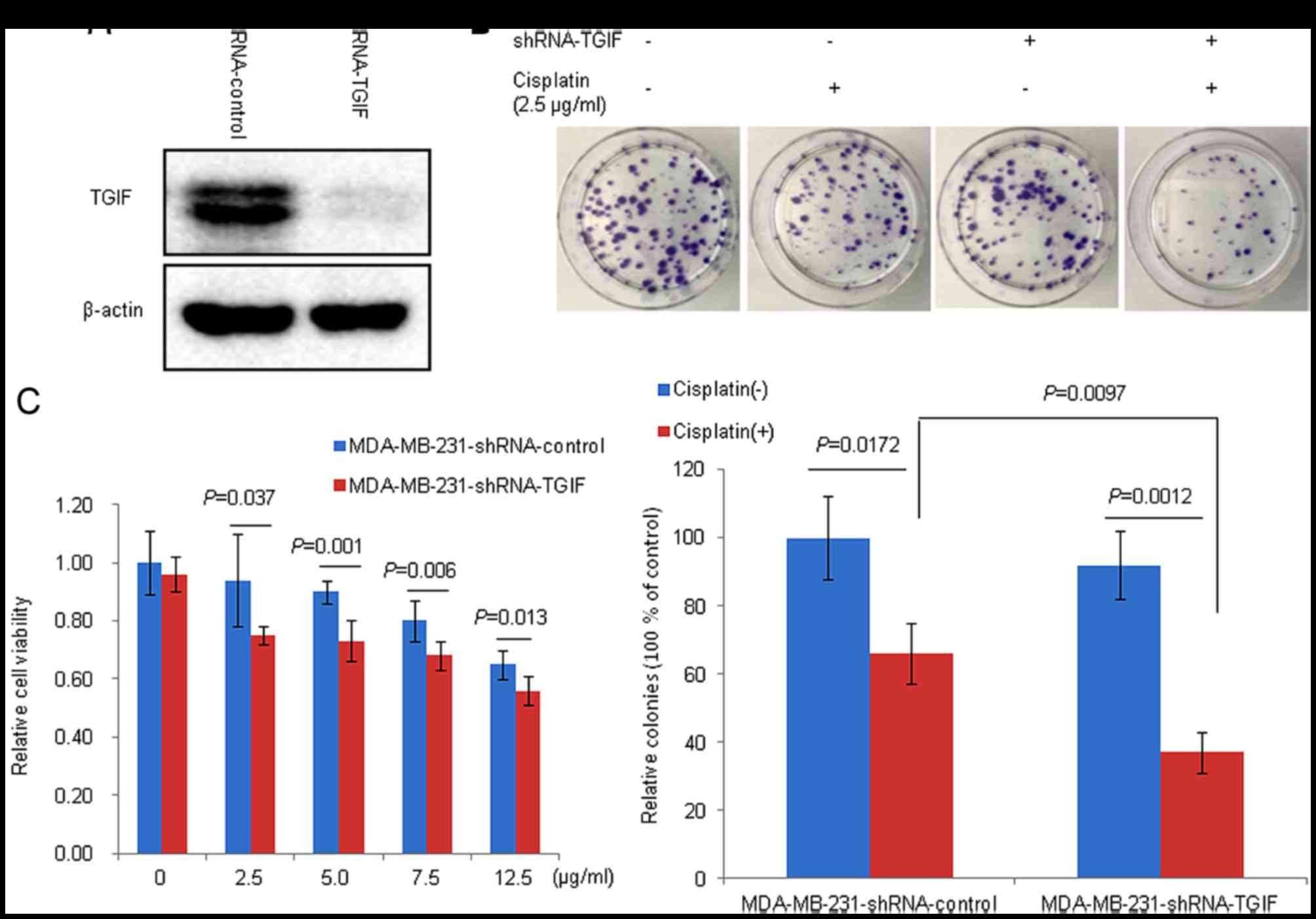
TGIF silencing efficiency in
MDA-MB-231 cells
Fig. 2A shows that
the level of TGIF protein expression in shRNA-TGIF infected cells
was markedly decreased compared with control cells. This suggested
a successful construction of a stable TGIF-silenced MDA-MB-231 cell
line.
Silencing TGIF increased the
cytotoxicity of MDA-MB-231 cells to cisplatin
Fig. 2B and C
demonstrated the effects of TGIF silencing on cisplatin-induced
cytotoxicity. Our data showed that TGIF silencing significantly
impeded cell proliferation in MDA-MB-231 cells compared with the
control groups in the presence of cisplatin (Fig. 2C). Furthermore, we observed that
there were significantly decreased colonies in the
MDA-MB-231-shRNA-TGIF group compared with the
MDA-MB-231-shRNA-control group in the presence of cisplatin
(Fig. 2B).
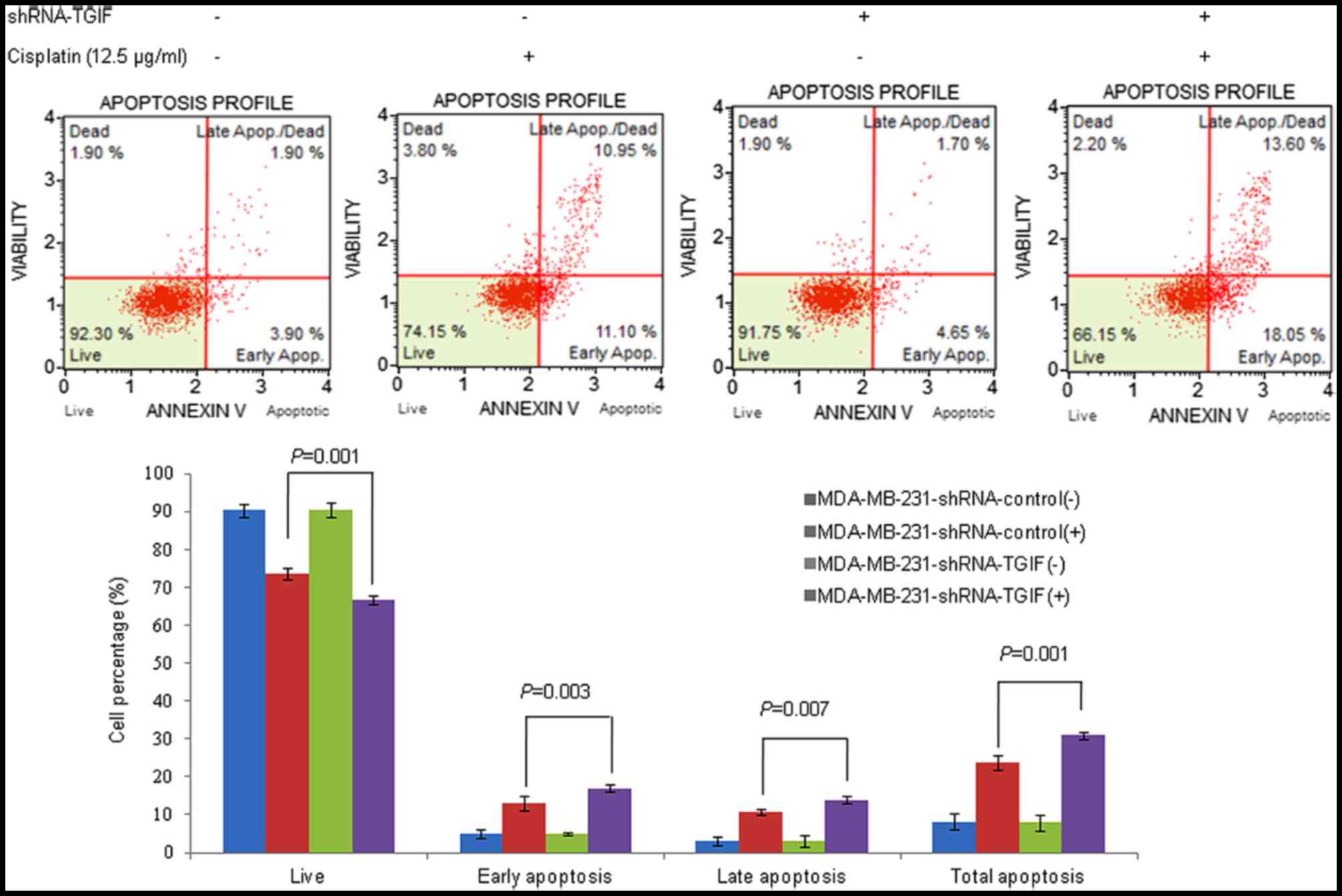
Silencing TGIF increased the apoptotic
sensitivity of MDA-MB-231 cells to cisplatin
Fig. 3 indicated the
effects of TGIF silencing on cisplatin-induced apoptosis in
MDA-MB-231 cells by annexin V and dead cell assay. Our results
showed that the percentage of apoptosis in MDA-MB-231-shRNA-TGIF
cells was much higher than that in MDA-MB-231-shRNA-control cells
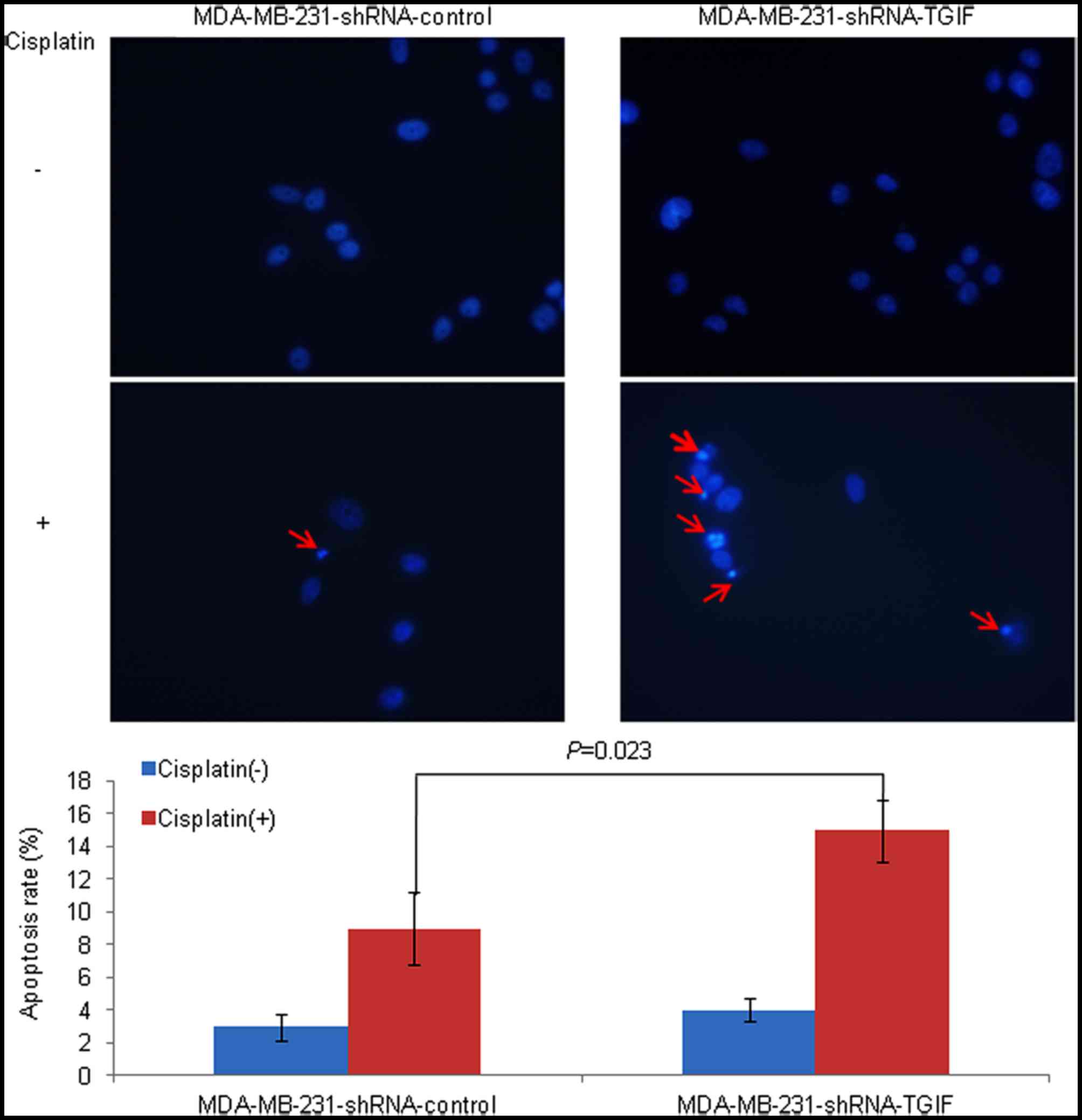
with the treatment of 12.5 µg/ml cisplatin (Fig. 3). Similar results were obtained from
the Hoechst 33258 staining assay. The MDA-MB-231-shRNA-TGIF cells
treated with cisplatin contained more cells in the apoptotic
morphological stages, while the control cells treated with
cisplatin were uniformly stained. After counting the cells, a
significantly increased percentage of apoptotic monocytes was
observed in MDA-MB-231-shRNA-TGIF cells compared with
MDA-MB-231-shRNA-control cells with the treatment of cisplatin
(Fig. 4).
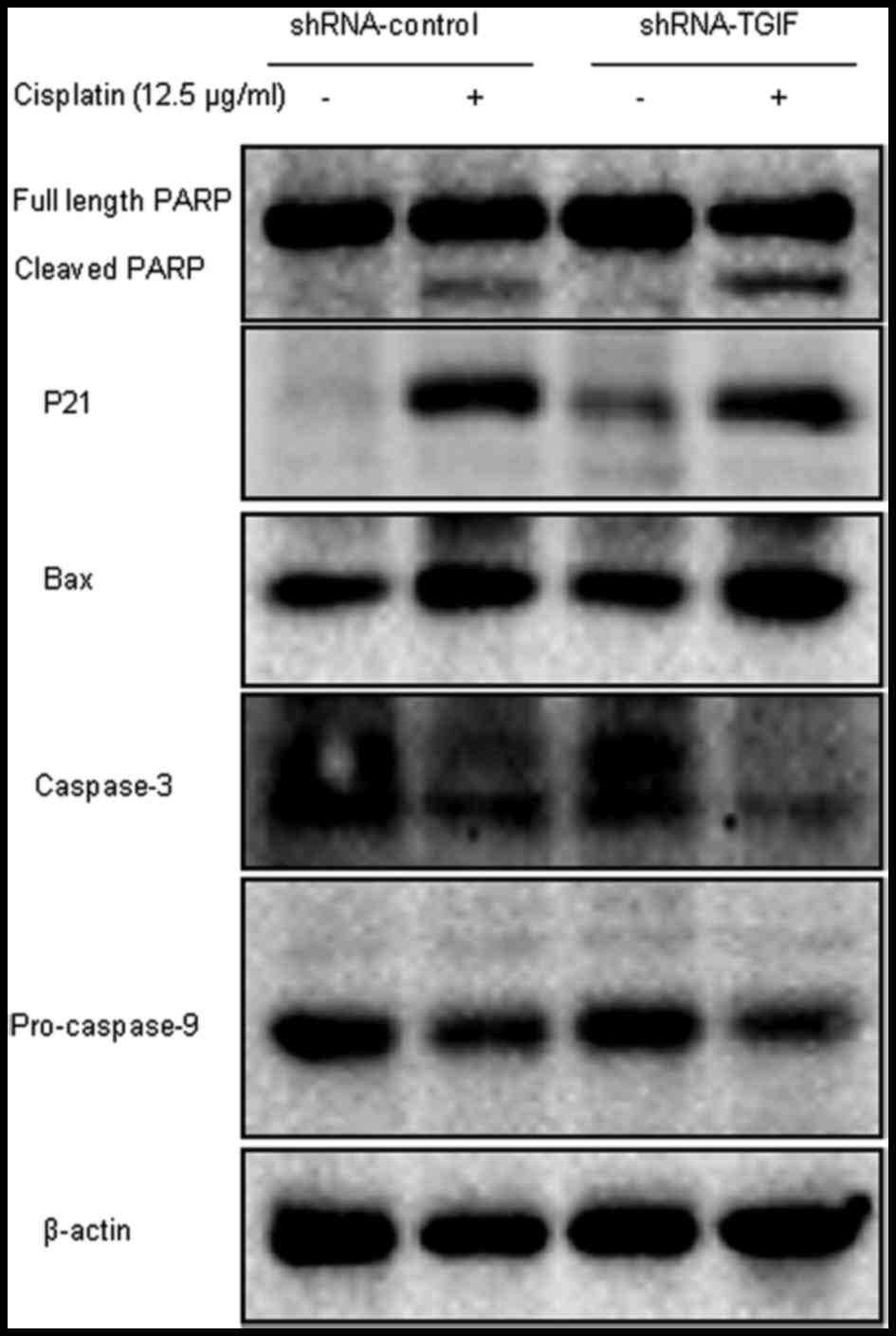
Silencing TGIF regulated the
expression of cell apoptosis-related proteins with the treatment of
cisplatin
As illustrated in Fig.
5, the increased expression of cleaved PARP, p21 and Bax
proteins and decreased expression of caspase-3 and pro-casepase-9
proteins were observed in MDA-MB-231-shRNA-control cells compared
with the cisplatin-untreated group with the treatment of 12.5 µg/ml
cisplatin. However, only PARP and caspase-3 were correlated with
the cisplatin-induced apoptosis mediated by TGIF protein in
MDA-MB-231 cells.
Discussion
Recently, several documents have shown that TGIF
might mediate a critical feature of chemoresistance in response to
the chemotherapy drugs of arsenic trioxide
(As2O3, ATO) and gemcitabine. Published
studies showed that ATO induced cell apoptosis via the ERK1/2
signaling and the TGF-β/Smad pathway, and enhanced
p21WAF1/CIP1 (p21) expression in human keratinocytes and
hepatocellular carcinoma cells. However, this effect could be
inhibited by ATO-induced TGIF expression through the recruitment of
the HDAC1/TGIF complex to the p21 promoter (32,33). In
addition, among the patients with upper tract urothelial carcinoma
treated with the gemcitabine and cisplatin systemic chemotherapy,
increased expression of TGIF was significantly associated with
worse oncological outcomes. Higher disease progression and
cancer-related death rates in the TGIF positive group were 100 and
83.3%, respectively, compared with 63.6 and 27.3% in the TGIF
negative group (34).
Cisplatin, also called cis-diamminedichloroplatinum
(II), is a representative of the platinum drugs. Since its
cytotoxic properties were discovered in the 1960s, it has gained
attention in the systemic treatment of cancer cells and now it has
been one of the most compelling chemotherapy drugs (6,35).
Cisplatin crosslinks with the purine bases on the DNA in the
nucleus and yields four main cisplatin-DNA adducts, including
interstrand crosslink, 1,2-intrastrand cross-link, 1,3-intrastrand
crosslink and DNA-protein cross-links (36,37).
Cisplatin-DNA adducts can be recognized by several families of
proteins and then can interfere with DNA repair mechanisms, induce
cell cycle arrest and DNA damage, and subsequently induce cell
apoptosis in cancer cells (38).
Previous studies have shown that ectopic expression
of several genes is associated with cisplatin therapeutic
sensitivity in MDA-MB-231 cells. Zhou et al reported that
silencing eIFE4 enhanced the chemosensitivity of MDA-MB-231 cells
to cisplatin (39). Yang et
al showed that knockdown of HAX-1 via RNA interference
decreased the IC50 level of cisplatin by 70.91% in
MDA-MB-231/CR cells (40). Hong
et al demonstrated that knockdown of BCL2L12 and BCL2L12A
dramatically inhibited cisplatin-induced apoptosis in MDA-MB-231
cells (41). Zhang et al
indicated that MiR-363 sensitized cisplatin-induced apoptosis
targeting in Mcl-1 in MDA-MB-231 cells; moreover, cells transfected
with Mcl-1 expression plasmid abolished the sensitization effects
of MiR-363 to cisplatin-inducing cytotoxicity (42). Wu et al showed that MiR-153
promoted MDA-MB-231 cell apoptosis treated with cisplatin by
targeting HECTD3. Moreover, stable overexpression of HECTD3
abrogated the sensitization effects of MiR-153 to cisplatin
treatments (43). Together, combined
treatment with gene therapy and chemotherapeutic drugs might
effectively enhance chemotherapy-induced cytotoxicity in TNBC cells
and may have great clinical significance.
In the present study, we explored the correlation of
TGIF silencing with cisplatin chemosensitivity in MDA-MB-231 cells.
Our data indicated that TGIF protein expression was markedly
decreased in a time- and dose-dependent manner in MDA-MB-231 cells
with the treatment of cisplatin, however, we did not observe
cisplatin had obvious effects on the expression of TGIF mRNA (0,
2.5, 7.5 and 12.5 µg/ml for 48 h or 7.5 µg/ml for 0, 24, 48 and 72
h; data not shown). The mechanism underlying cisplatin repressing
TGIF expression should be explored in future studies. We observed
that cell proliferation was significantly inhibited in
TGIF-silenced cells treated by cisplatin. Furthermore, silencing
TGIF reduced the formation of colonies of MDA-MB-231 cells in the
presence of cisplatin compared with controls. In addition, TGIF
silencing dramatically increased the cisplatin-induced apoptosis
rate as measured by flow cytometry assay as well as the Hoechst
staining assay. These data revealed that TGIF might be a potential
therapeutic target for cisplatin treatment in MDA-MB-231 cells. For
further investigation, the expression of apoptosis-related proteins
was measured by western blot analysis. The expression of PARP and
caspase-3 proteins was involved in the event that silencing TGIF
protein enhanced cisplatin-induced apoptosis.
As is well-known, caspase-3 is the key molecule in
the execution phases of apoptosis (44,45).
Caspase-3 is usually present in the cytoplasm in the form of an
inactive 32-kDa precursor; after the stimulation of the apoptosis
signal, it can produce 17 kDa (p17) and 12 kDa (p12) subunits and
later form a mature enzyme that is responsible for the proteolytic
cleavage of many key proteins. PARP is one of the main cleavage
targets of caspase-3 in vivo; the cleavage is typically a
marker of cells undergoing apoptosis. When a cell is engaged in
cell apoptosis, autoribosylated PARP is cleaved by caspases into
the 89 kDa C-terminal fragment and the 24 kDa N-terminal fragment;
these fragments interact with intact PARP and block the function of
PARP on DNA repair and the maintenance of genomic stability
(46,47). Presently, we found that caspase-3 and
PARP cleavage were involved in cisplatin-induced apoptosis mediated
by TGIF protein. However, the mechanisms that are involved should
be elucidated further.
We should acknowledge that there are some
limitations in our present study. First, only one cell line of
MDA-MB-231 human TNBC has been applied to investigate the effects
of TGIF knockdown on cisplatin-induced apoptosis; other breast
cancer cell lines representing different types of breast cancer
should be used to verify our current results. Second, in
vitro experiments, such as cell proliferation assay, colony
formation assay and apoptosis assay, were performed to observe the
effects of TGIF silencing on cytotoxicity and apoptosis in
MDA-MB-231 cells induced by cisplatin; a tumor xenograft model
should be performed in future studies to confirm our findings.
Third, it would be interesting to investigate the expression of
TGIF in breast cancer patients who develop resistance to cisplatin
in clinical treatment. Fourth, a TGIF-overexpressed cell line
should be constructed in future studies to observe whether TGIF
overexpression could result in breast cancer cells being resistant
to cisplatin; this may verify our present data in other
aspects.
In summary, our results showed, for the first time,
that silencing TGIF by RNA intervention promoted cell apoptosis and
cell cytotoxicity to cisplatin treatment in MDA-MB-231 cells. In
addition, this effect might be attributed to caspase-3 protein and
its cleavage substrate PARP. Given the association of TGIF protein
with a chemotherapeutic response in MDA-MB-231 TNBC cells, targeted
therapies directed at TGIF may be a potential novel strategy for
improving the chemotherapy response in TNBC.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. U1404815).
References
|
1
|
Agarwal G, Nanda G, Lal P, Mishra A,
Agarwal A, Agrawal V and Krishnani N: Outcomes of triple-negative
breast cancers (TNBC) compared with non-TNBC: Does the survival
vary for all stages? World J Surg. 40:1362–1372. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lehmann BD, Bauer JA, Chen X, Sanders ME,
Chakravarthy AB, Shyr Y and Pietenpol JA: Identification of human
triple-negative breast cancer subtypes and preclinical models for
selection of targeted therapies. J Clin Invest. 121:2750–2767.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kumar P and Aggarwal R: An overview of
triple-negative breast cancer. Arch Gynecol Obstet. 293:247–269.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Palma G, Frasci G, Chirico A, Esposito E,
Siani C, Saturnino C, Arra C, Ciliberto G, Giordano A and D'Aiuto
M: Triple negative breast cancer: Looking for the missing link
between biology and treatments. Oncotarget. 6:26560–26574. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yadav BS, Chanana P and Jhamb S:
Biomarkers in triple negative breast cancer: A review. World J Clin
Oncol. 6:252–263. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Merrouche Y, Fabre J, Cure H, Garbar C,
Fuselier C, Bastid J, Antonicelli F, Al-Daccak R, Bensussan A and
Giustiniani J: IL-17E synergizes with EGF and confers in vitro
resistance to EGFR-targeted therapies in TNBC cells. Oncotarget.
7:53350–53361. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Von Minckwitz G and Martin M: Neoadjuvant
treatments for triple-negative breast cancer (TNBC). Ann Oncol. 23
Suppl 6:vi35–vi39. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Siddik ZH: Cisplatin: Mode of cytotoxic
action and molecular basis of resistance. Oncogene. 22:7265–7279.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen Y, Gao Y, Zhang K, Li C, Pan Y, Chen
J, Wang R and Chen L: MicroRNAs as regulators of cisplatin
resistance in lung cancer. Cell Physiol Biochem. 37:1869–1880.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bartholin L, Powers SE, Melhuish TA, Lasse
S, Weinstein M and Wotton D: TGIF inhibits retinoid signaling. Mol
Cell Biol. 26:990–1001. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ferrand N, Demange C, Prunier C, Seo SR
and Atfi A: A mechanism for mutational inactivation of the
homeodomain protein TGIF in holoprosencephaly. FASEB J. 21:488–496.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hu Y, Yu H, Shaw G, Renfree MB and Pask
AJ: Differential roles of TGIF family genes in mammalian
reproduction. BMC Dev Biol. 11:582011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Massagué J: TGFbeta in cancer. Cell.
134:215–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sharma M and Sun Z: 5′TG3′ interacting
factor interacts with Sin3A and represses AR-mediated
transcription. Mol Endocrinol. 15:1918–1928. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wotton D, Lo RS, Lee S and Massagué J: A
smad transcriptional corepressor. Cell. 97:29–39. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zerlanko BJ, Bartholin L, Melhuish TA and
Wotton D: Premature senescence and increased TGFβ signaling in the
absence of Tgif1. PLoS One. 7:e354602012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gripp KW, Wotton D, Edwards MC, Roessler
E, Ades L, Meinecke P, Richieri-Costa A, Zackai EH, Massagué J,
Muenke M and Elledge SJ: Mutations in TGIF cause holoprosencephaly
and link NODAL signalling to human neural axis determination. Nat
Genet. 25:205–208. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Wang H, Gao H, Xu B, Zhai W, Li J
and Zhang C: Elevated expression of TGIF is involved in lung
carcinogenesis. Tumour Biol. 36:9223–9231. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Pan T, Wang H, Li L, Li J, Zhang C
and Yang H: Silencing of TGIF attenuates the tumorigenicity of A549
cells in vitro and in vivo. Tumour Biol. 37:12725–12730. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiang G, Yi Y, Weiwei H and Weiming W:
TGIF1 promoted the growth and migration of cancer cells in nonsmall
cell lung cancer. Tumour Biol. 36:9303–9310. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Borlak J, Meier T, Halter R, Spanel R and
Spanel-Borowski K: Epidermal growth factor-induced hepatocellular
carcinoma: Gene expression profiles in precursor lesions, early
stage and solitary tumours. Oncogene. 24:1809–1819. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu ZM, Tseng HY, Tsai HW, Su FC and Huang
HS: Transforming growth factor β-interacting factor-induced
malignant progression of hepatocellular carcinoma cells depends on
superoxide production from Nox4. Free Radic Biol Med. 84:54–64.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Imoto I, Pimkhaokham A, Watanabe T,
Saito-Ohara F, Soeda E and Inazawa J: Amplification and
overexpression of TGIF2, a novel homeobox gene of the TALE
superclass, in ovarian cancer cell lines. Biochem Biophys Res
Commun. 276:264–270. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hamid R and Brandt SJ: Transforming
growth-interacting factor (TGIF) regulates proliferation and
differentiation of human myeloid leukemia cells. Mol Oncol.
3:451–463. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hu ZL, Wen JF, Xiao DS, Zhen H and Fu CY:
Effects of transforming growth interacting factor on biological
behaviors of gastric carcinoma cells. World J Gastroenterol.
11:84–88. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yeh BW, Wu WJ, Li WM, Li CC, Huang CN,
Kang WY, Liu ZM and Huang HS: Overexpression of TG-interacting
factor is associated with worse prognosis in upper urinary tract
urothelial carcinoma. Am J Pathol. 181:1044–1055. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nakakuki K, Imoto I, Pimkhaokham A, Fukuda
Y, Shimada Y, Imamura M, Amagasa T and Inazawa J: Novel targets for
the 18p11.3 amplification frequently observed in esophageal
squamous cell carcinomas. Carcinogenesis. 23:19–24. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang MZ, Ferrigno O, Wang Z, Ohnishi M,
Prunier C, Levy L, Razzaque M, Horne WC, Romero D, Tzivion G, et
al: TGIF governs a feed-forward network that empowers Wnt signaling
to drive mammary tumorigenesis. Cancer Cell. 27:547–560. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Razzaque MS and Atfi A: TGIF function in
oncogenic Wnt signaling. Biochim Biophys Acta. 1865:101–104.
2016.PubMed/NCBI
|
|
31
|
Kwon YJ, Leibovitch BA, Bansal N, Pereira
L, Chung CY, Ariztia EV, Zelent A, Farias EF and Waxman S: Targeted
interference of SIN3A-TGIF1 function by SID decoy treatment
inhibits Wnt signaling and invasion in triple negative breast
cancer cells. Oncotarget. 8:88421–88436. 2016.PubMed/NCBI
|
|
32
|
Huang HS, Liu ZM and Hong DY: Blockage of
JNK pathway enhances arsenic trioxide-induced apoptosis in human
keratinocytes. Toxicol Appl Pharmacol. 244:234–241. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu ZM, Tseng JT, Hong DY and Huang HS:
Suppression of TG-interacting factor sensitizes arsenic
trioxide-induced apoptosis in human hepatocellular carcinoma cells.
Biochem J. 438:349–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yeh BW, Li WM, Li CC, Kang WY, Huang CN,
Hour TC, Liu ZM, Wu WJ and Huang HS: Histone deacetylase inhibitor
trichostatin A resensitizes gemcitabine resistant urothelial
carcinoma cells via suppression of TG-interacting factor. Toxicol
Appl Pharmacol. 290:98–106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tsimberidou AM, Braiteh F, Stewart DJ and
Kurzrock R: Ultimate fate of oncology drugs approved by the us food
and drug administration without a randomized Trial. J Clin Oncol.
27:6243–6250. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gonzalez VM, Fuertes MA, Alonso C and
Perez JM: Is cisplatin-induced cell death always produced by
apoptosis? Mol Pharmacol. 59:657–663. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cepeda V, Fuertes MA, Castilla J, Alonso
C, Quevedo C and Pérez JM: Biochemical mechanisms of cisplatin
cytotoxicity. Anticancer Agents Med Chem. 7:3–18. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Galluzzi L, Senovilla L, Vitale I, Michels
J, Martins I, Kepp O, Castedo M and Kroemer G: Molecular mechanisms
of cisplatin resistance. Oncogene. 31:1869–1883. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou FF, Yan M, Guo GF, Wang F, Qiu HJ,
Zheng FM, Zhang Y, Liu Q, Zhu XF and Xia LP: Knockdown of eIF4E
suppresses cell growth and migration, enhances chemosensitivity and
correlates with increase in Bax/Bcl-2 ratio in triple-negative
breast cancer cells. Med Oncol. 28:1302–1307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang J, Wu Y, Wang X, Xu L, Zhao X and
Yang Y: Chemoresistance is associated with overexpression of HAX-1,
inhibition of which resensitizes drug-resistant breast cancer cells
to chemotherapy. Tumour Biol. 39:10104283176922282017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hong Y, Yang J, Wu W, Wang W, Kong X, Wang
Y, Yun X, Zong H, Wei Y, Zhang S and Gu J: Knockdown of BCL2L12
leads to cisplatin resistance in MDA-MB-231 breast cancer cells.
Biochim Biophys Acta. 1782:649–657. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang R, Li Y, Dong X, Peng L and Nie X:
MiR-363 sensitizes cisplatin-induced apoptosis targeting in Mcl-1
in breast cancer. Med Oncol. 31:3472014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu X, Li L, Li Y and Liu Z: MiR-153
promotes breast cancer cell apoptosis by targeting HECTD3. Am J
Cancer Res. 6:1563–1571. 2016.PubMed/NCBI
|
|
44
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Del Bello B, Valentini MA, Mangiavacchi P,
Comporti M and Maellaro E: Role of caspases-3 and −7 in Apaf-1
proteolytic cleavage and degradation events during
cisplatin-induced apoptosis in melanoma cells. Exp Cell Res.
293:302–310. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Soldani C and Scovassi AI:
Poly(ADP-ribose) polymerase-1 cleavage during apoptosis: An update.
Apoptosis. 7:321–328. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Michels J, Vitale I, Galluzzi L, Adam J,
Olaussen KA, Kepp O, Senovilla L, Talhaoui I, Guegan J, Enot DP, et
al: Cisplatin resistance associated with PARP hyperactivation.
Cancer Res. 73:2271–2280. 2013. View Article : Google Scholar : PubMed/NCBI
|