Introduction
Oestrogen deficiency has been suggested to be the
predominant cause of post-menopausal osteoporosis (1). Accompanied by the withdrawal of sex
steroids in mammals, serum levels of the pituitary hormone
follicle-stimulating hormone (FSH) are markedly increased (2,3). FSH
exhibits osteoclastogenic and proresorptive actions in a
Gi2α-coupled FSH receptor (FSHR) identified in human and mouse
osteoclasts and their oestrogen-independent precursors (4). In osteoclasts, FSHR activation
enhances the phosphorylation of downstream receptor activator of
nuclear factor κB ligand (RANKL)-sensitive kinases, extracellular
signal-regulated kinases, protein kinase B and inhibitor of κB to
transduce the proresorptive actions of RANKL (5). The highly circulating FSH exhibits a
strong positive correlation with bone remodelling markers;
alterations in FSH circulating levels can predict bone mass
remodeling (6). A recent study
additionally observed that FSH-mediated effects on the secretion of
inflammatory factors, particularly interleukin-1 (IL-1) and tumour
necrosis factor α (TNF-α), may be partly involved in regulating the
resorptive activity of osteoclasts (7).
Periapical bone loss is a prominent pathological and
clinical feature of periapical periodontitis, which begins as a
bacterial infection in the dental pulp; inflammatory cells are then
recruited and cytokines are subsequently generated (8). It has been identified that the
regulatory mechanisms of apical periodontitis are complex, and
remain to be fully elucidated. Bone-resorptive cytokines, including
IL-1 and TNF-α, and bone resorptive factors, in particular RANKL,
serve key roles in pulpal and periapical pathogenesis, including
bone destruction (9). In addition,
systemic factors, such as hormones, are able to influence
periapical pathogenesis. In the periapical lesions of
ovariectomised (OVX) rats, more severe bone destruction has been
observed (10). Increased RANKL
expression levels have been observed in periapical lesions of OVX
rats, which are attributed to oestrogen deficiency (11). Further studies have indicated that
OVX mice yield high FSH levels as oestrogen levels are reduced.
FSH, independent of oestrogen, can aggravate alveolar bone loss
through FSHR in periodontitis; the FSH inhibitor leuprorelin (LE)
elicits protective effects on alveolar bone loss resulting from
experimental periapical lesions of OVX rats (12). However, it remains unclear whether
FSH serves a direct role in the progression of apical
periodontitis.
The current study aimed to assess the effects and
the associated mechanisms of FSH on apical periodontitis-associated
bone loss. Enzyme histochemical examination, radioimmunoassay and
immunohistochemical staining were conducted in the present study in
order to detect the following: FSHR, a novel positive biomarker of
osteoclasts; RANKL, a positive regulator of osteoclastogenesis and
osteoclastic activity; osteoprotegerin (OPG), a negative regulator
of osteoclastogenesis and osteoclastic activity; and TNF-α and
IL-1β, inflammatory cytokines.
Materials and methods
Animal experimental groups
A total of 90 12-week-old female Sprague-Dawley
rats, weighing 180–200 g, were purchased from the Laboratory Animal
Centre of Wuhan University (Wuhan, China) and were housed with a
temperature of 24–26°C and a 12 h light/dark cycle. The rats were
randomly assigned into the following six groups with 15 rats/group:
i) Group 1, Sham surgery + 0.1 mol/l phosphate-buffered saline
(PBS; pH 7.4) as a vehicle, ii) group 2, bilateral OVX + vehicle,
iii) group 3, bilateral OVX + 1.6 mg/kg LE (Takeda Pharmaceutical
Company, Ltd., Osaka, Japan), iv) group 4, bilateral OVX + 1.6
mg/kg LE + 1.85 µg/kg luteinizing hormone (LH; Serono; Merck
Millipore, Geneva, Switzerland), v) group 5, bilateral OVX + 1.6
mg/kg LE + 3 µg/kg FSH (Serono; Merck Millipore) and vi)
group 6, bilateral OVX + 3 µg/kg FSH. The experimental
protocols were approved by the Institutional Animal Care and Use
Committee of Wuhan University.
Ovariectomy for rats
Rats were anaesthetised by intraperitoneal injection
3 mg/ml pentobarbital (10 ml/kg) (Veterinary Institute of Military
Supplies University, Changchun, China). Subsequently, the rats in
groups 2–6 were subjected to bilateral OVX. The rats in group 1
were subjected to Sham surgeries with similar-sized fatty tissues
near to the removed ovaries. Subsequent to surgery, LE, Vehicle, LH
or FSH were subcutaneously injected into the rats in groups 3–5,
groups 1 and 2, group 4 or groups 5 and 6 at the relevant
concentrations, respectively. As a slow-release formulation, LE can
maintain effective blood concentration and inhibit increases of
circulating LH and FSH levels for a minimum of 4 weeks subsequent
to injection.
Induction of experimental periapical
lesions
At the 7th day following surgery, all rats were
anaesthetised as previously described (13,14).
The pulps of the mandibular first molars were exposed using a #1/2
round bur under a surgical microscope (M320 F12, Leica Microsystems
GmbH, Wetzlar, Germany). The exposure size was equal to the
diameter of the bur to avoid furcal perforation. The exposed pulp
chambers were left to the oral environment without any restoration
during the entire experimental period. After pulpal exposure, FSH,
LH, LE and vehicle were subcutaneously injected in the back of rats
daily.
Measurement of estradiol (E2), FSH and LH
serum levels
On the 7th, 14th and 21st day after pulpal exposure,
the venous blood samples were collected and centrifuged at 1,000 ×
g for 10 min at 4°C. The serum was then separated and acquired to
test E2, FSH and LH levels using a radioimmunoassay kit (North
institute of Biological Technology, Beijing, China) according to
the manufacturer's instructions.
Histological observation
On the 7th, 14th and 21st day after pulpal exposure,
rats were anaesthetised with an intraperitoneal injection of 3
mg/ml pentobarbital (10 ml/kg; Veterinary Institute of Military
Supplies University, Changchun, China) and sacrificed by cervical
dislocation. Bilateral mandibles were removed and fixed with 4%
paraformaldehyde at 4°C for 2 days. In order for histological
observation to be conducted, samples were decalcified with 10%
ethylenediaminetetraacetic acid (EDTA) at 4°C for ~6 weeks. The
decalcified specimens were then embedded in paraffin and cut into
pieces at a thickness of 4 µm using a microtome from Jung AG
(Heidelberg, Germany). The sections were then stained with
haematoxylin/eosin or immunohistochemistry antibodies. Subsequent
to haematoxylin/eosin staining, samples were photographed under a
light microscope. The areas of periapical lesions were measured by
SPOT RT software, version 3.5 (Diagnostic Instruments, Inc.,
Sterling Heights, MI, USA) in three random HE-stained sections in a
double-blind manner as previously described (10,15).
Immunohistochemistry
Subsequent dewaxing and rehydration, the slides were
treated with 3% hydrogen peroxide at 37°C for 15 min to inhibit any
endogenous peroxidase activity. The slices were then incubated
separately with the following primary antibodies: Polyclonal goat
anti-FSHR (1:200; Santa Cruz Biotechnology, Inc., Dallas, TX, USA;
cat. no. sc-7797), polyclonal goat anti-RANKL (1:100; Santa Cruz
Biotechnology, Inc.; cat. no. sc-7628), polyclonal goat anti-OPG
(1:200; Santa Cruz Biotechnology, Inc.; cat. no. sc-8468),
polyclonal rabbit anti-TNF-α (1:300; Bioss, Beijing, China; cat.
no. 2081R), monoclonal hamster anti-IL-1β (1:300; Biolegend, Inc.,
San Diego, CA, USA; cat. no. 503501) for 24 h at 4°C. Subsequent to
incubation, the primary antibody was removed by washing the slides
thrice with PBS for 5 min. The sections were incubated with
secondary antibody kits from ZSGB-Bio (Beijing, China) according to
the manufacturer's protocols. The kits were as follows: Polink-2
Plus® Polymer HRP Detection system for the antibodies
raised in goat; and SPlink Detection kit (Biotin-Streptavidin HRP
Detection system) for the remaining primary antibodies. The slides
were stained for approximately 30–40 sec with diaminobenzidine and
the nucleus was counterstained with haematoxylin for 10 sec.
Enzyme histochemical examination
Periapical region slices were used to detect TRAP
activity using the TRAP Detection kit (Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany). In brief, EDTA-decalcified tissue
sections were rehydrated and incubated for 1 h at 37°C with the
TRAP staining solution. Following completion of incubation, these
sections were stained with haematoxylin. Certain sections were
incubated in a substrate-free medium, which were used as controls
for TRAP activity.
Statistical analysis
For each specimen, positive cells from five randomly
selected areas at the periapical region were counted at a
magnification of ×400 under a light microscope. GraphPad Prism
software, version 5.0 for Windows (GraphPad Software, Inc., La
Jolla, CA, USA) was used to analyse the data. The differences in
lesion size and the average number per high-power field were
analysed by one-way analysis of variance and post-hoc Tukey or
Bonferroni multiple comparison tests were conducted between the
subgroups.
Results
Levels of E2, FSH and LH in serum
From 7 days after ovariectomy, compared with the
Sham group, levels of E2 in the serum of all OVX groups were
reduced significantly and plateaued at a level approximately 1/4 of
that of the Sham group (P<0.01), with no statistical differences
identified among the OVX groups (P>0.05; Fig. 1A). The levels of FSH and LH
observed in the serum were increased significantly, when compared
with that of the Sham group subsequent to ovariectomy, however,
this increase was prevented by the application of LE (P<0.05;
Fig. 1B and C). The levels of FSH
were increased markedly subsequent to application of FSH in the
FSH-treated group, with the levels observed approximately 10x
greater than that observed in the OVX group (P<0.05; Fig. 1B). The levels of LH measured in the
OVX + LE + LH group were increased rapidly and plateaued from the
third week subsequent to administration of LH (P<0.05; Fig. 1C).
 | Figure 1Levels of E2, FSH and LH in serum. (A)
From 7 days after ovariectomy, the levels of E2 were significantly
reduced in the serum of all OVX groups vs. Sham group, and
plateaued at a level ~1/4 of that of the Sham group (P<0.01),
with no statistical differences among the OVX groups (P>0.05).
(B and C) Levels of FSH and LH in serum were significantly
increased vs. the Sham group after ovariectomy, and this increase
was prevented by application of LE (P<0.05). Levels of FSH
rapidly increased following application in the FSH-treated group,
with levels approximately 10× greater than that of the OVX group
(P<0.05). Levels of LH in the OVX + LE + LH group rapidly
increased and plateaued from the third week after administration of
LH (P<0.05). ◊P<0.05 vs. the OVX-only and FSH
treatment groups; #P<0.05 vs. the non-FSH treatment
group; ΔP<0.05 vs. the non-LH treatment groups. E2,
estradiol; FSH, follicle-stimulating hormone; LH, luteinizing
hormone; OVX, ovariectomised; LE, leuprorelin. |
Histological analysis
The histological analysis is presented in Fig. 2. Compared with those in the Sham
group, specimens in the OVX groups exhibited significant increases
in bone loss of periapical lesions (P<0.05; Fig. 2A, B and G), which were reversed by
administration of LE or LE + LH (P>0.05; Fig. 2D–G). Compared with those in the
Sham and OVX groups, bone loss of periapical lesions were
significantly increased following administration of FSH (P<0.05;
Fig. 2C, E and G).
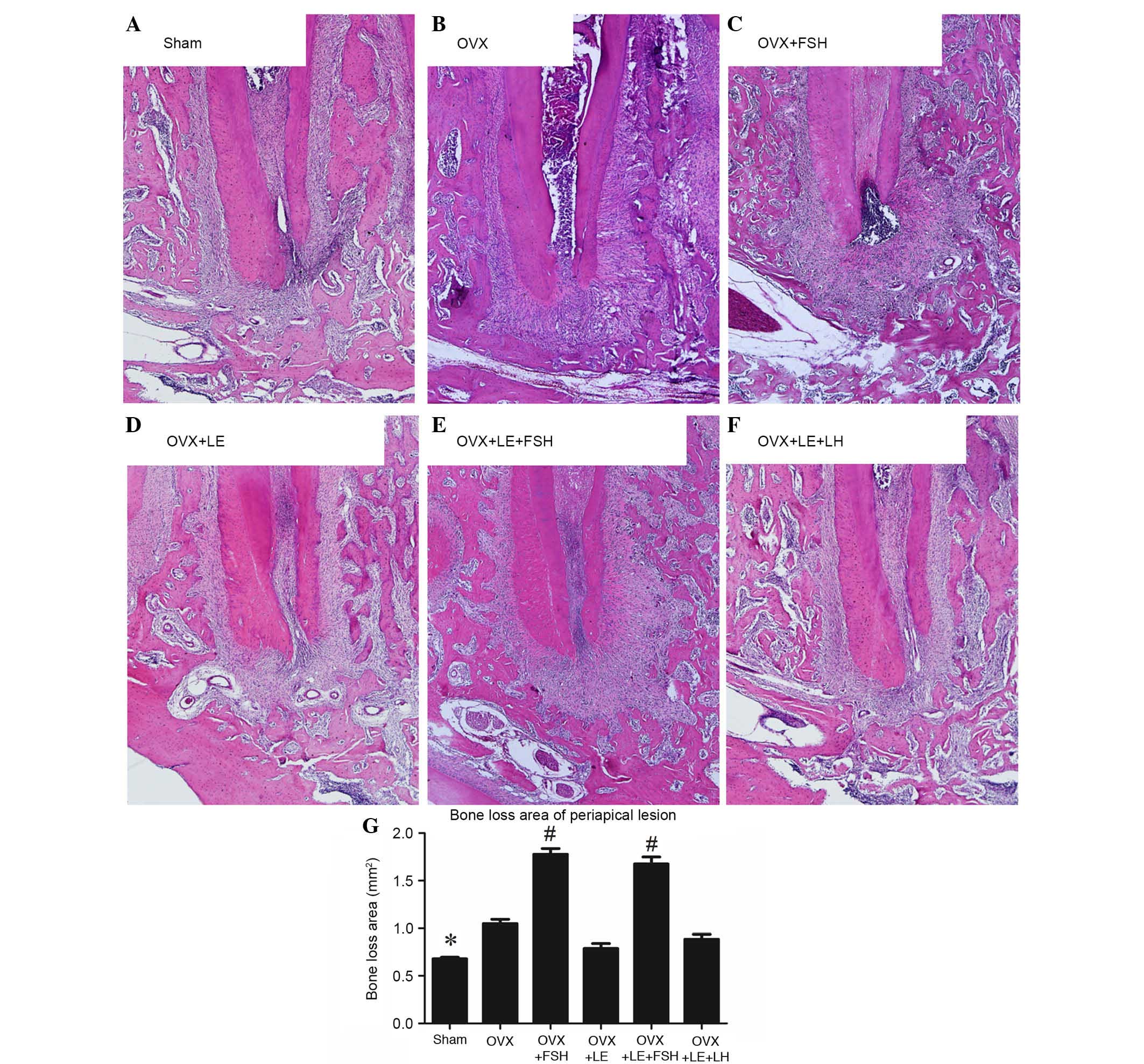 | Figure 2Histological analysis of periapical
lesions in each group. (A) Sham, (B) OVX, (C) OVX + FSH, (D) OVX +
LE, (E) OVX + LE + FSH and (F) OVX + LE + LH groups. (G)
Quantitative analysis of periapical lesions in each group. Compared
with those in the Sham group, specimens in the OVX groups exhibited
significant increases in bone loss of periapical lesions
(P<0.05; A, B and G), which were reversed by the administration
of LE or LE + LH (P>0.05; D–G). Bone loss of periapical lesions
was significantly increased vs. those in the Sham and OVX groups
after administration of FSH (P<0.05; C, E and G). Data is
presented as the mean ± standard deviation. *P<0.05
vs. the OVX only group and the FSH treatment group;
#P<0.05 vs. non-FSH treatment groups. OVX,
ovariectomised; FSH, follicle-stimulating hormone; LE, leuprorelin;
LH, luteinizing hormone. |
Immunohistochemical observation of
osteoclasts and FSHR expression
TRAP-positive cells were stained dark red to purple,
and multinuclear cells were considered osteoclasts (Fig. 3A–F). On the 7th, 14th and 21st day
following pulpal exposure, the numbers of osteoclasts in the OVX
group were higher than that of Sham group (P<0.01; Fig. 3A, B and G). The numbers of
osteoclasts in OVX + LE + LH and the OVX + LE groups were
significantly lower than those in the OVX group, with no
significant differences between the OVX + LE + LH and OVX + LE
groups themselves (P<0.01; Fig. 3C,
D and G). The numbers of osteoclasts were significantly greater
in the OVX + FSH and OVX + LE + FSH groups than those in OVX and
OVX + LE groups (P<0.01; Fig. 3E, F
and G).
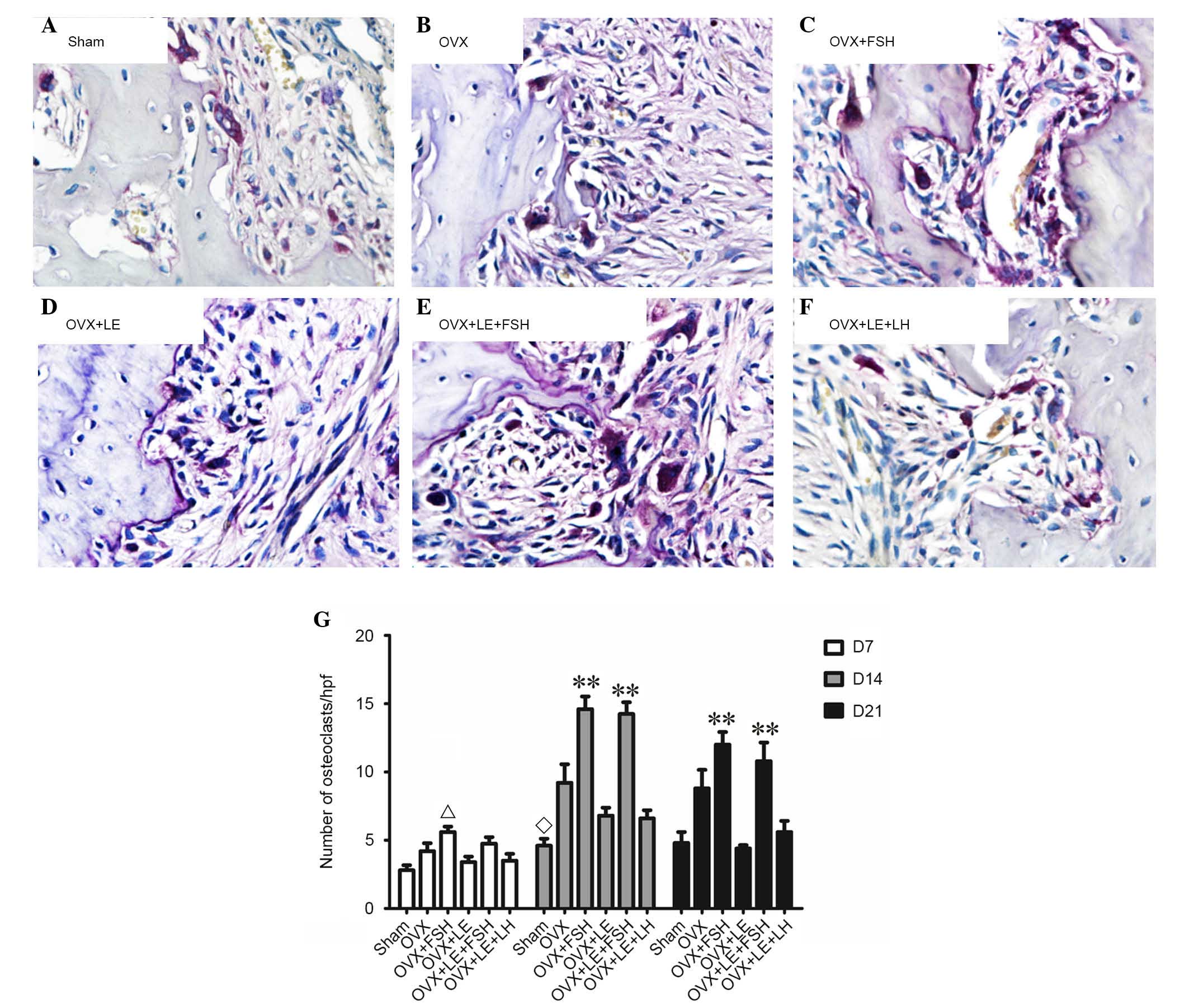 | Figure 3Immunohistochemical observation of
osteoclasts. (A) Sham, (B) OVX, (C) OVX + FSH, (D) OVX + LE, (E)
OVX + LE + FSH and (F) OVX + LE + LH groups. Tartrate-resistant
acid phosphatase-positive cells were stained dark red to purple,
and multinuclear cells were considered osteoclasts. (G)
Quantitative analysis of osteoclast number in each group. On the
7th, 14th and 21st day after pulpal exposure, the numbers of
osteoclasts in OVX group were increased compared with that of the
Sham group (P<0.01; A, B and G). The numbers of osteoclasts in
the OVX + LE + LH and OVX + LE groups were significantly greater
than those in the OVX groups (P<0.01, C, D and G), with no
significant differences observed between them. The numbers of
osteoclasts were significantly greater in the OVX + FSH and OVX +
LE + FSH groups compared with those in the OVX and OVX + LE groups
(P<0.01, E, F and G). Data is presented as the mean ± standard
deviation. ΔP<0.05 vs. the sham D7 group;
◊P<0.05 vs. the OVX D14 group and the FSH treatment
group; **P<0.01 vs. non-FSH treatment groups on D14
and D21. OVX, ovariectomised; FSH, follicle-stimulating hormone;
LE, leuprorelin; LH, luteinizing hormone. |
FSHR-positive cells were stained brown (Fig. 4A–F). On the 7th, 14th and 21st day
subsequent to pulpal exposure, the numbers of FSHR-positive cells
in the OVX group were observed to be greater than that of the Sham
group (P<0.05; Fig. 4A, B and
G). In the FSH-treated groups, the number of FSHR-positive
cells was significantly increased compared with those in the Sham,
OVX or OVX + LE groups (P<0.05; Fig. 4C–E and G). There was no difference
between the OVX + LE and OVX + LE + LH groups (P>0.05; Fig. 4D, F and G).
 | Figure 4Immunohistochemical observation of
FSHR expression. (A) Sham, (B) OVX, (C) OVX + FSH, (D) OVX + LE,
(E) OVX + LE + FSH and (F) OVX + LE + LH groups. (G) Quantitative
analysis of FSHR expression in each group. FSHR-positive cells were
stained brown. On the 7th, 14th and 21st day after pulpal exposure,
the numbers of FSHR-positive cells in OVX group were significantly
greater than that of the Sham group (P<0.05; A, B and G). In the
FSH-treated groups, the number of FSHR-positive cells was increased
significantly compared with those in the Sham, OVX or OVX + LE
groups (P<0.05; C–E and G). There was no difference between the
OVX + LE and OVX + LE + LH groups (P>0.05; D, F and G). Data is
presented as the mean ± standard deviation. **P<0.01
vs. non-FSH treatment groups on D7, D14 and D21. FSH,
follicle-stimulating hormone; FSHR, FSH receptor; OVX,
ovariectomised; LE, leuprorelin; LH, luteinizing hormone. |
Comparison of RANKL and OPG
expressions
RANKL- and OPG-positive cells were stained brown
(Figs. 5A–F and 6A–F). On the 7th, 14th and 21st days
after pulpal exposure, the number of RANKL- and OPG-positive cells
in each group reached maximal values on day 14 (P<0.05; Figs. 5G and 6G). At each time point, the number of
RANKL-positive cells in the FSH-treated groups was significantly
greater than those in the corresponding non-FSH treatment groups
(P<0.05; Fig. 5B–E). There were
no significant differences identified between the FSH-treated
groups and the corresponding groups in the number of OPG-positive
cells (P>0.05; Fig. 6B–E). The
numbers of RANKL- and OPG-positive cells were not significantly
different between the OVX + LE and OVX + LE + LH groups at each
time point (P>0.05; Figs. 5F and
G and 6F and G).
 | Figure 5Immunohistochemical observation of
RANKL expression. (A) Sham, (B) OVX, (C) OVX + FSH, (D) OVX + LE,
(E) OVX + LE + FSH and (F) OVX + LE + LH groups. (G) Quantitative
analysis of RANKL-positive expression in each group. RANKL-positive
cells were stained brown. On the 7th, 14th and 21st day after
pulpal exposure, the numbers of RANKL-positive cells in each group
respectively reached the maximum value at 14th day (P<0.05; G).
At each time point, the number of RANKL-positive cells in
FSH-treated groups was significantly greater compared with those in
the corresponding non-FSH treatment groups (P<0.05; B–E). The
numbers of RANKL-positive cells were not observed to be
significantly different between the OVX + LE and OVX + LE + LH
groups at each time point (P>0.05; F and G). Data is presented
as the mean ± standard deviation. ◊P<0.05 vs. the
OVX-only groups and the FSH treatment groups on D7, D14 and D21;
**P<0.01 vs. non-FSH treatment groups on D7, D14 and
D21. RANKL, receptor activator of nuclear factor κB ligand; OVX,
ovariectomised; FSH, follicle-stimulating hormone; LE, leuprorelin;
LH, luteinizing hormone. |
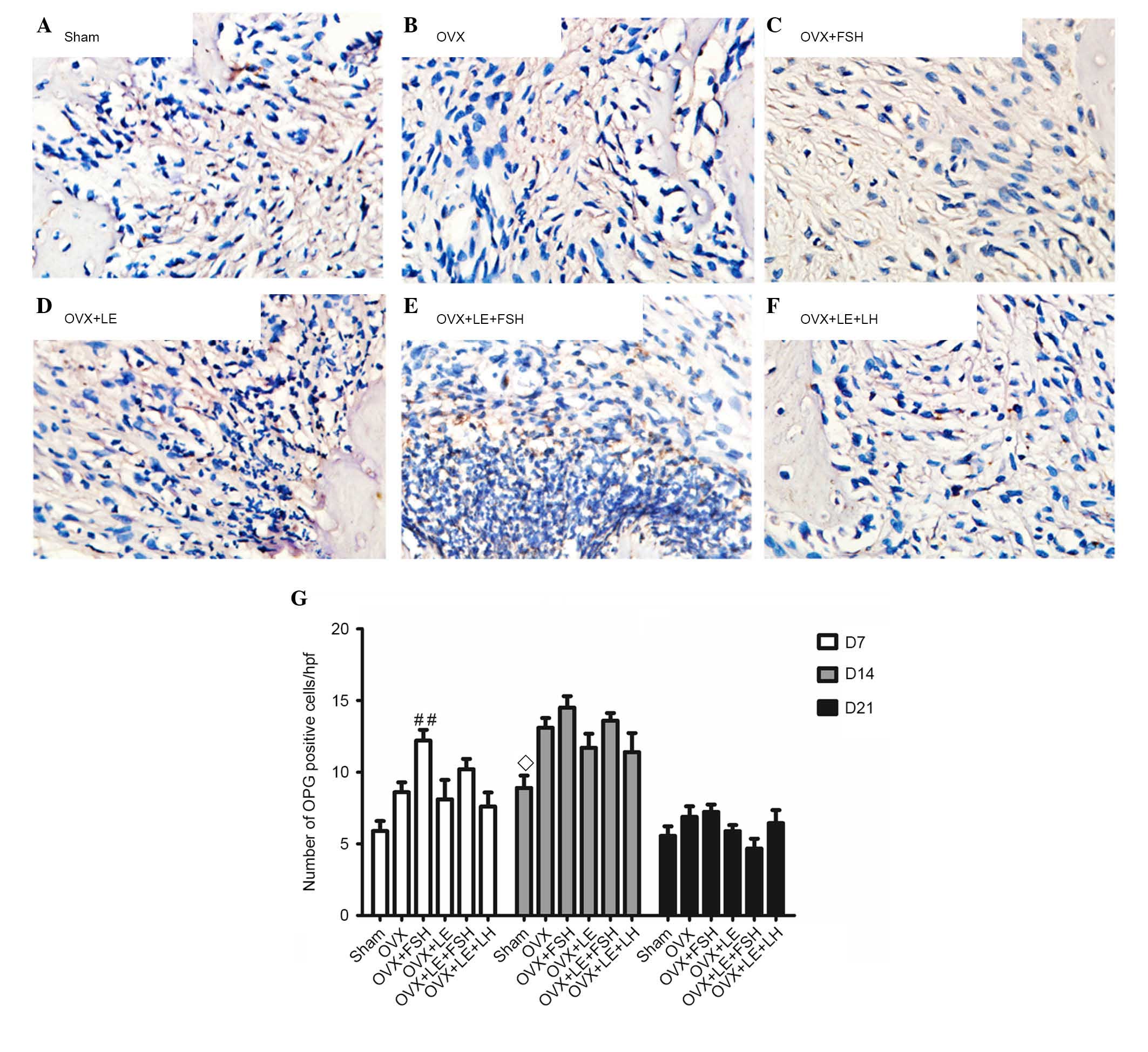 | Figure 6Immunohistochemical observation of OPG
expression. (A) Sham, (B) OVX, (C) OVX + FSH, (D) OVX + LE, (E) OVX
+ LE + FSH and (F) OVX + LE + LH groups. (G) Quantitative analysis
of OPG-positive expression in each group. OPG-positive cells were
stained brown. On the 7th, 14th and 21st day after pulpal exposure,
the numbers of OPG-positive cells in each group peaked at day 14
(P<0.05; G). There were no significant differences observed
between FSH-treated groups and corresponding groups in the number
of OPG-positive cells (P>0.05; B–E). The numbers of OPG-positive
cells exhibited no significant differences between OVX + LE and OVX
+ LE + LH groups at each time point (P>0.05; F and G). Data is
presented as the mean ± standard deviation. ◊P<0.05
vs. the OVX D14 group; ##P<0.01 vs. the sham D7
group. OPG, osteoprotegerin; OVX, ovariectomised; FSH,
follicle-stimulating hormone; LE, leuprorelin; LH, luteinizing
hormone. |
Comparison of TNF-α and IL-1β
expressions
TNF-α- and IL-1β-positive cells were stained brown
and increased as the lesion expanded (Figs. 7A–F and 8A–F). The majority of these cells were
likely inflammatory cells predominantly centralised at the
periapical bone destruction areas and periodontal ligaments. The
numbers of TNF-α- and IL-1β-positive cells in the OVX group were
increased compared with those in the Sham groups at each time point
(P<0.05; Figs. 7A and B and
8A and B). The numbers of TNF-α-
and IL-1β-positive cells were increased in the FSH-treated mice
when compared with those in the corresponding non-FSH-treated mice
(P<0.05; Figs. 7B–E and
8B–E). The numbers of TNF-α- and
IL-1β-positive cells were not significantly different between the
OVX + LE and OVX + LE + LH groups at each time point (P>0.05;
Figs. 7F and G and 8F and G).
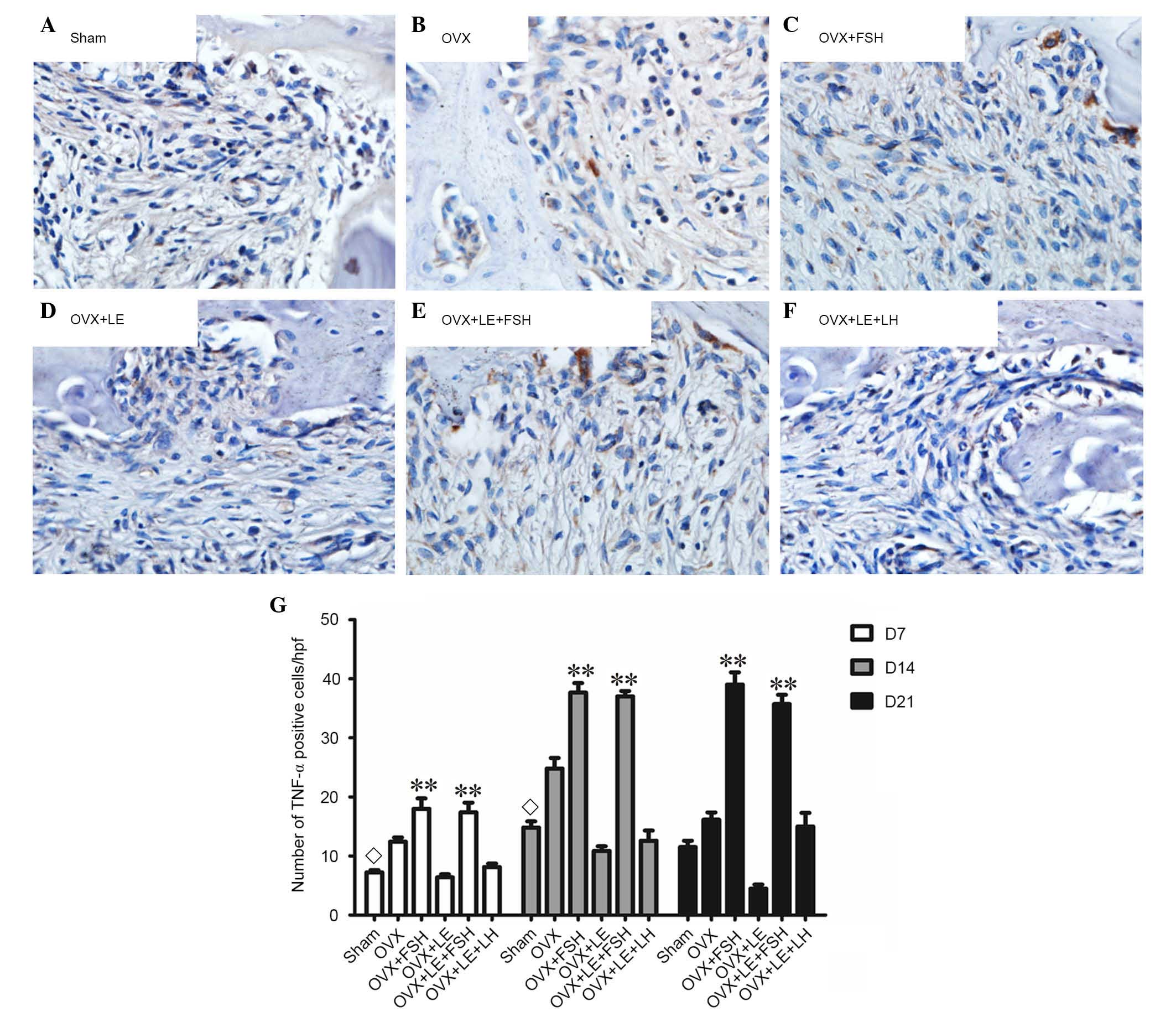 | Figure 7Immunohistochemical observation of
TNF-α expression. (A) Sham, (B) OVX, (C) OVX + FSH, (D) OVX + LE,
(E) OVX + LE + FSH and (F) OVX + LE + LH groups. (G) Quantitative
analysis of TNF-α-positive expression in each group. TNF-α-positive
cells were stained brown and increased as the lesion expanded. The
numbers of TNF-α-positive cells in the OVX group were larger than
those in Sham groups at each time point (P<0.05; A and B). The
numbers of TNF-α-positive cells were larger in FSH-treated mice
than those in the corresponding non-FSH-treated mice (P<0.05;
B–E). The numbers of TNF-α-positive cells were not significantly
different between the OVX + LE and OVX + LE + LH groups at each
time point (P>0.05; F and G). Data is presented as the mean ±
standard deviation. ◊P<0.05 vs. the OVX-only group on
D7 and D14; **P<0.01 vs. the non-FSH treatment groups
on D7, D14 and D21. TNF-α, tumour necrosis factor α; OVX,
ovariectomised; FSH, follicle-stimulating hormone; LE, leuprorelin;
LH, luteinizing hormone. |
 | Figure 8Immunohistochemical observation of
IL-1β expression. (A) Sham, (B) OVX, (C) OVX + FSH, (D) OVX + LE,
(E) OVX + LE + FSH and (F) OVX + LE + LH groups. (G) Quantitative
analysis of IL-1β-positive expression in each group. IL-1β-positive
cells were stained brown and increased as the lesion expanded. The
numbers of IL-1β-positive cells in OVX group were greater than
those in the Sham groups at each time point (P<0.05; A and B).
The numbers of IL-1β-positive cells were greater in FSH-treated
mice when compared with those in the corresponding non-FSH-treated
mice (P<0.05; B–E). The numbers of IL-1β-positive cells were not
observed to be significantly different between the OVX + LE and OVX
+ LE + LH groups at each time point (P>0.05; F and G). Data is
presented as the mean ± standard deviation. ◊P<0.05
vs. the OVX-only group on D7, D14 and D21; **P<0.01
vs. the non-FSH treatment groups on D7, D14 and D21. IL-1β,
interleukin 1β; OVX, ovariectomised; FSH, follicle-stimulating
hormone; LE, leuprorelin; LH, luteinizing hormone. |
Discussion
Post-menopausal osteoporosis, a global public health
problem, has been attributed to reducing oestrogen levels. FSH
levels increase rapidly as oestrogen levels are reduced during the
post-menopausal period (16). FSH
directly affects skeletal remodelling by stimulating the formation
and the function of osteoclasts in vitro and in vivo
(16). In the current study, a rat
model was established to verify the effects of FSH in experimental
periapical lesions. The results indicated that FSH aggravated the
bone loss of periapical lesions. In addition, RANKL-, TRAP-, TNF-α-
and IL-1β-positive cells were increased significantly in the
FSH-treated groups, which indicated that the function of FSH in
bone loss may be mediated via the increasing activity of
osteoclasts and the increased secretion of inflammatory
cytokines.
Although ovariectomies markedly affect
mineralisation density of mandibular and maxillary alveolar bones,
ovariectomy alone cannot elucidate the effects of FSH (17). In the current study, a rat model
was successfully established to verify the independent role of FSH
by administering FSH, LH and LE based on OVX (12). In the current study, E2 levels did
not differ among all the OVX groups. However, FSH levels
significantly differed among the different OVX groups. For example,
the comparison results of OVX vs. OVX + FSH group and OVX + LE vs.
OVX + LE + FSH group indicated the effects of extrinsic FSH in
periapical bone loss.
Ovariectomy in mice has been reported to exacerbate
induced periapical lesions; this condition is attributed to
oestrogen deficiency (10,18). In the present study, similar
alterations in alveolar bone loss were observed in the OVX group
compared with the Sham group. In addition, significant differences
in the numbers of osteoclasts and the areas of bone loss were
observed in the OVX group vs. OVX + FSH group and in OVX + LE group
vs. OVX + LE + FSH group. The bone loss on day 21 in the OVX + FSH
group was the most pronounced among all of the groups. Furthermore,
the administration of LE, an FSH inhibitor, resulted in a reduction
of bone loss in the OVX + LE and OVX + LE + LH groups compared with
the OVX group. No significant differences were identified between
the OVX + LE and OVX + LE + LH groups. Therefore, it is suggested
that FSH, independent of oestrogen, can aggravate alveolar bone
loss of induced periapical lesions.
In the current study, it was additionally identified
that administration of FSH may significantly increase the
expression of RANKL. The numbers of FSHR-positive cells were
positively correlated with those of RANKL- and TRAP-positive cells.
RANKL/OPG is an important factor influencing the production of
osteoclasts (19,20). FSH may also influence RANKL
signalling via FSHR due to the fact that FSH and RANKL served an
important role in osteoclastogenesis (6). RANKL expression in bone marrow cells
of post-menopausal women has been reported to be greater than that
of pre-menopausal women (21–23).
Furthermore, RANKL is implicated in mediating increased bone
resorption in early postmenopausal women (24). An additional study reported that
RANKL-stimulated osteoclastogenesis is enhanced by FSH in human
peripheral blood mononuclear cells (25). As an osteoblastic factor, OPG
expression in the OVX + FSH group was unexpectedly greater than
those in other groups at day 7, which indicated that
osteoblastogenesis was also accelerated in the early stages of high
FSH level administration, as a type of reactive-protective
response. In other groups, no significant differences in the
expression of OPG were observed, which suggested that FSH may alter
the balance of the RANKL/OPG system by increasing RANKL expression
levels.
The results of the current study suggested that
higher expression levels of TNF-α and IL-1β were present in the
FSH-treated groups, when compared with those in the non-FSH-treated
groups. This indicated that the effects of FSH on periapical
lesions may be associated with the upregulation of TNF-α and IL-1β
expression. Although oestrogen deficiency increases TNF-α
expression in certain instances, such as after ovariectomy, FSH is
required to increase TNF-α expression similar to accompanying bone
loss (26). A previous study
reported that despite severe hypogonadism, FSH stimulates TNF-α
production, as demonstrated by loss-of-function studies indicating
the presence of lowered TNF-α levels in FSH β-deficient mice
(27). FSH triggers the production
of TNF-α from macrophages and granulocytes in the bone marrow
(27). In turn, TNF-α are able to
stimulate osteoclastogenesis directly and strongly synergise with
IL-1 and RANKL to promote osteoclast differentiation and activation
(28). FSH may influence bone
mineral density by modulating the activity of the osteoresorptive
cytokine IL-1β. Furthermore, FSH increases the secretion of IL-1β,
TNF-α and IL-6 as surface expression of FSHR on monocytes increases
(29).
FSH accelerated the progression of induced
experimental periapical lesions, which may be mediated by
increasing secretion of osteoresorptive cytokines (TNF-α and IL-1β)
and altered RANK/OPG ratios. The findings of the present study
suggest that FSH, independent of oestrogen, may aggravate
periapical bone loss by FSH receptors, which may serve an important
role in the immune and inflammatory response of the host to root
canal and periradicular infection during menopause.
Acknowledgments
The current study was supported by grants from the
National Natural Science Foundation of China (grant no.
81120108010) and the Pre-National Basic Research Program of China
(973 Plan; grant no. 2012CB722404).
References
|
1
|
Han NR, Kim HY, Yang WM, Jeong HJ and Kim
HM: Glutamic acid ameliorates estrogen deficiency-induced
menopausal-like symptoms in ovariectomized mice. Nutr Res.
35:774–783. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wei S, Gong Z, An L, Zhang T, Dai H and
Chen S: Cloprostenol and pregnant mare serum gonadotropin promote
estrus synchronization, uterine development, and
follicle-stimulating hormone receptor expression in mice. Genet Mol
Res. 14:7184–7195. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Santi D, Granata AR and Simoni M: FSH
treatment of male idiopathic infertility improves pregnancy rate: A
meta-analysis. Endocr Connect. 4:R46–R58. 2015.PubMed/NCBI
|
|
4
|
Herrera-Luna CV, Scarlet D, Walter I and
Aurich C: Effect of stallion age on the expression of LH and FSH
receptors and aromatase P450 in equine male reproductive tissues.
Reprod Fertil Dev. 6–Jul;2015.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cannon JG, Kraj B and Sloan G:
Follicle-stimulating hormone promotes RANK expression on human
monocytes. Cytokine. 53:141–144. 2011. View Article : Google Scholar :
|
|
6
|
Robinson LJ, Tourkova I, Wang Y, Sharrow
AC, Landau MS, Yaroslavskiy BB, Sun L, Zaidi M and Blair HC:
FSH-receptor isoforms and FSH-dependent gene transcription in human
monocytes and osteoclasts. Biochem Biophys Res Commun. 394:12–17.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Petrova NL, Petrov PK, Edmonds ME and
Shanahan CM: Inhibition of TNF-α Reverses the pathological
resorption pit profile of osteoclasts from patients with acute
Charcot osteoarthropathy. J Diabetes Res. 2015:9179452015.
View Article : Google Scholar
|
|
8
|
Tolias D, Koletsi K, Mamai-Homata E,
Margaritis V and Kontakiotis E: Apical periodontitis in association
with the quality of root fillings and coronal restorations: A
14-year investigation in young Greek adults. Oral Health Prev Dent.
10:297–303. 2012.PubMed/NCBI
|
|
9
|
Zhang X and Peng B: Immunolocalization of
receptor activator of NF kappa B ligand in rat periapical lesions.
J Endod. 31:574–577. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiong H, Peng B, Wei L, Zhang X and Wang
L: Effect of an estrogen-deficient state and alendronate therapy on
bone loss resulting from experimental periapical lesions in rats. J
Endod. 33:1304–1308. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang X, Peng B, Fan M, Bian Z and Chen Z:
The effect of estrogen deficiency on receptor activator of nuclear
factor kappa B ligand and osteoprotegerin synthesis in periapical
lesions induced in rats. J Endod. 33:1053–1056. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu S, Cheng Y, Fan M, Chen D and Bian Z:
FSH aggravates periodontitis-related bone loss in ovariectomized
rats. J Dent Res. 89:366–371. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Suzuki N, Takimoto K and Kawashima N:
Cathepsin K inhibitor regulates inflammation and bone destruction
in experimentally induced rat periapical lesions. J Endod.
41:1474–1479. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Velickovic M, Pejnovic N, Mitrovic S,
Radosavljevic G, Jovanovic I, Kanjevac T, Jovicic N and Lukic A:
ST2 deletion increases inflammatory bone destruction in
experimentally induced periapical lesions in mice. J Endod.
41:369–375. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin SK, Hong CY, Chang HH, Chiang CP, Chen
CS, Jeng JH and Kuo MYP: Immunolocalization of macrophages and
transforming growth factor-β1 in induced rat periapical lesions. J
Endod. 26:335–340. 2000. View Article : Google Scholar
|
|
16
|
Sun L, Peng Y, Sharrow AC, Iqbal J, Zhang
Z, Papachristou DJ, Zaidi S, Zhu LL, Yaroslavskiy BB, Zhou H, et
al: FSH directly regulates bone mass. Cell. 125:247–260. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rawlinson SC, Boyde A, Davis GR, Howell
PG, Hughes FJ and Kingsmill VJ: Ovariectomy vs. hypofunction: Their
effects on rat mandibular bone. J Dent Res. 88:615–620. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gomes-Filho JE, Wayama MT, Dornelles RC,
Ervolino E, Yamanari GH, Lodi CS, Sivieri-Araújo G, Dezan-Júnior E
and Cintra LT: Raloxifene modulates regulators of
osteoclastogenesis and angiogenesis in an oestrogen deficiency
periapical lesion model. Int Endod J. 48:1059–1068. 2015.
View Article : Google Scholar
|
|
19
|
Pitari MR, Rossi M, Amodio N, Botta C,
Morelli E, Federico C, Gullà A, Caracciolo D, Di Martino MT,
Arbitrio M, et al: Inhibition of miR-21 restores RANKL/OPG ratio in
multiple myeloma-derived bone marrow stromal cells and impairs the
resorbing activity of mature osteoclasts. Oncotarget.
6:27343–27358. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sağlam M, Köseoğlu S, Hatipoğlu M, Esen HH
and Köksal E: Effect of sumac extract on serum oxidative status,
RANKL/OPG system and alveolar bone loss in experimental
periodontitis in rats. J Appl Oral Sci. 23:33–41. 2015.
|
|
21
|
Duan P, Wang ZM, Liu J, Wang LN, Yang Z
and Tu P: Gene polymorphisms in RANKL/RANK/OPG pathway are
associated with ages at menarche and natural menopause in Chinese
women. BMC Womens Health. 15:322015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Duan P, Wang ZM, Liu J, Wang LN, Yang Z
and Tu P: Association of gene polymorphisms in RANKL/RANK/OPG
system with hypertension and blood pressure in Chinese women. J Hum
Hypertens. 29:749–753. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tu P, Duan P, Zhang RS, Xu DB, Wang Y, Wu
HP, Liu YH and Si L: Polymorphisms in genes in the RANKL/RANK/OPG
pathway are associated with bone mineral density at different
skeletal sites in post-menopausal women. Osteoporos Int.
26:179–185. 2015. View Article : Google Scholar
|
|
24
|
Xu XJ, Shen L, Yang YP, Zhu R, Shuai B, Li
CG and Wu MX: Serum β-Catenin levels associated with the ratio of
RANKL/OPG in patients with postmenopausal osteoporosis. Int J
Endocrinol. 2013:5343522013. View Article : Google Scholar
|
|
25
|
Marazzi J, Kleyer J, Paredes JM and
Gertsch J: Endocannabinoid content in fetal bovine sera-unexpected
effects on mononuclear cells and osteoclastogenesis. J Immunol
Methods. 373:219–228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cenci S, Toraldo G, Weitzmann MN, Roggia
C, Gao Y, Qian WP, Sierra O and Pacifici R: Estrogen deficiency
induces bone loss by increasing T cell proliferation and lifespan
through IFN-gamma-induced class II transactivator. Proc Natl Acad
Sci USA. 100:10405–10410. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Karadag F, Ozcan H, Karul AB, Yilmaz M and
Cildag O: Sex hormone alterations and systemic inflammation in
chronic obstructive pulmonary disease. Int J Clin Pract.
63:275–281. 2009. View Article : Google Scholar
|
|
28
|
Kobayashi K, Takahashi N, Jimi E, Udagawa
N, Takami M, Kotake S, Nakagawa N, Kinosaki M, Yamaguchi K, Shima
N, et al: Tumor necrosis factor alpha stimulates osteoclast
differentiation by a mechanism independent of the ODF/RANKL-RANK
interaction. J Exp Med. 191:275–286. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lam J, Takeshita S, Barker JE, Kanagawa O,
Ross FP and Teitelbaum SL: TNF-alpha induces osteoclastogenesis by
direct stimulation of macrophages exposed to permissive levels of
RANK ligand. J Clin Invest. 106:1481–1488. 2000. View Article : Google Scholar : PubMed/NCBI
|














