Introduction
Inhabitants who live at >3,000 m elevation may
experience damage to the heart, brain, lungs and other organs due
to the low inspired PO2 and the reduced barometric
pressure (1–4). Furthermore, Zhang et al
(5) reported that 4,500 m
elevation is the turning point for high-altitude polycythemia
(HAPC) prevalence in Tibetan communities. However, the Mount
Everest (8,848 m), being well renowned as the roof of the world,
appears to be close to the limit of human tolerance to hypoxia
(6,7). Hence, these previous data provide
evidence to suggest that the three altitudes of 3,000, 4,500 and
8,848 (~9,000) m are points of elevation which are notable in
research conducted on plateau hypoxia.
In previous years, an increasing number of
lowlanders have travelled to high-altitude areas for recreation or
work all year round. Unfortunately, individuals travelling to
high-altitude areas from low-altitude areas have the potential risk
of developing mountain sickness due to exposure to a hypobaric
hypoxia environment at a high altitude, including acute mountain
sickness, high-altitude pulmonary edema, high-altitude cerebral
edema, chronic mountain sickness, high-altitude pulmonary
hypertension and HAPC (8–11). These diseases can be
life-threatening. Therefore, the improvement of the endurance of
humans in resisting hypoxia and reducing hypoxia-induced organ
damage is a global challenge.
Considering the issues mentioned above, the purpose
of the present study was to establish an equably simulated acute
plateau anoxia brain injury model of Sprague-Dawley (SD) rats and
reliable methodology validation. The present study aimed to provide
a foundation for the investigation into the mechanisms and
molecular-targeted therapeutic drugs used for hypoxic brain
injury.
Materials and methods
Reagents
Urethane was purchased from Aladdin Shanghai
Biochemical Technology Co., Ltd (Shanghai, China). Hematoxylin and
eosin (H&E) were provided by Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). The total extraction sample kit (cat. no.
AR0101-30), BCA protein assay kit (cat. no. AR0146), sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) protein
sample buffer 2X (denaturation; cat. no. AR0131), broad spectrum
protease inhibitor (cat. no. AR1182-1), broad spectrum phosphatase
inhibitor (cat. no. AR1183), terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) apoptosis detection kit I-POD (cat.
no. MK1025) and primary antibodies against Bcl-2-associated X
protein (Bax; cat. no. BA0315), apoptotic protease activating
factor-1 (Apaf-1; cat. no. BA2373), hypoxia inducible factor
(HIF)-1α (cat. no. PB0245) and cytochrome c (cyto-c; cat.
no. A03529) were obtained from Wuhan Boster Biological Technology,
Ltd. (Wuhan, China). Caspase-3 (cat. no. #9662), and cleaved
caspase-3 (cat. no. #9661) antibodies were procured from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Antibodies against
β-actin (cat. no. GB11001) and horseradish peroxidase-conjugated
goat anti-rabbit immunoglobulin G (H+L; cat. no. GB23303) were
obtained from Servicebio (Wuhan, China). Ultrasignal
electrochemiluminescence (ECL) substrate (cat. no. 4AW011-100) was
obtained from 4A Biotech Co., Ltd (Beijing, China). Enzyme-linked
immunosorbent assay (ELISA) kits for lactate dehydrogenase (LDH;
cat. no. A020-2), superoxide dismutase (SOD; cat. no. A001-3),
malondialdehyde (MDA; cat. no. A003-1) and glutathione/oxidized
glutathione (GSH/GSSG; cat. no. A061-1) were provided by Nanjing
Jiancheng Bioengineering Institute (Nanjing, China).
Animals
A total of 12 male specific pathogen-free SD rats
(weighing 200–220 g, 7 weeks old) were obtained from Chengdu Dashuo
Experimental Animal Co., Ltd (Chengdu, China) and were maintained
in a 12 h light/dark cycle at room temperature (23±2°C) in 50–60%
relative humidity. They were randomly divided into two groups of
six rats each and were fed chow and water ad libitum.
Animals were maintained for one week for the purpose of
acclimatization prior to the experimental treatment. All rats were
anesthetized using an intraperitoneal injection of 20% urethane
(0.5 ml/100 g).
Ethical statement
All procedures were performed in accordance with the
Animal Care Guidelines, conforming to the Health Guide for the Care
and Use of Laboratory Animals (12), and the protocol of this
investigation was ethically approved by the Animal Research Ethics
Committee of Chengdu University of Traditional Chinese Medicine
(Chengdu, China).
Simulated high-altitude hypoxia brain
injury model
Based on a previous investigation on HAPC or
high-altitude cerebral injury rat models (13) and other similar studies (14–17),
a modified version of an animal model of acute hypobaric hypoxia
brain injury model was developed. In brief, subsequent to
acclimatization for 30 min in an animal decompression chamber (AVIC
Guizhou Fenglei Aviation Armament Co., Ltd., Guizhou, China; cat.
no. FLYDWC50-II C), which is able to simulate a ~10,000 m global
atmospheric condition by the reduction of ambient barometric
pressure in combination with precise monitoring temperature
(−40-60°C) and relative humidity (0–100%). An anoxic environment
was simulated by evacuating the air from the chamber using powerful
vacuum pumps. Fresh air was allowed to circulate, and the air flow
was maintained inside the chamber at 0.9l per min to replenish
consumed O2 and remove the produced CO2. The
hypoxic rats were subjected to the following protocols: 3,000 and
4,500 m altitude at a speed of 5 m/s for 30 min and subsequent
simulated 9,000 m altitude for 23 h. The chamber was maintained at
a temperature of 15–17°C and a relative humidity of 55–60%.
Following hypobaric exposure for 24 h, the elevation was
immediately lowered at a speed of 5 m/s. Meanwhile, the control
group was subject to similar conditions at an altitude of ~600 m
outside of the chamber, in order to simulate a pure high altitude
environment that is lacking in oxygen and at a low pressure rather
compared with a low altitude environment with normal air
circulation. The protocol of the present study design is presented
in Fig. 1.
Determination of oxidative stress
indicators in serum
The levels of SOD, MDA, GSH, GSSG and LDH in the
sera of six rats that had undergone 24 h simulated high-altitude
hypoxic treatment were measured using ELISA kits according to the
manufacturers' protocol. The absorbance values of SOD and LDH were
read at 450 nm, MDA at 532 nm and GSSG and total (T-)GSH at 405 nm
using a full-wavelength scanning multifunction reading meter
(Varioskan Flash; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The concentration of GSH was calculated by the formula:
GSH=T-GSH-2×GSSG. The values were expressed in terms of units SOD
per ml, nmol MDA per ml, µmol GSH and GSSG per l and units LDH per
l.
Hematoxylin & eosin (H&E)
staining and immunohistochemical detection
The serum was acquired from the abdominal aorta of
the rats. Subsequently, the whole brain was removed from the skull
and fixed using 100 ml 4% paraformaldehyde at a 7.2 pH at 25°C for
24 h. The paraffin-embedded sections of the hippocampal tissue were
sliced into 5 µm thick sections using cryotome (cat. no. RM2235;
Leica Microsystems GmbH, Wetzlar, Germany), baked at 60°C overnight
and dewaxed using xylene I and II for 20 min. Then, the sections
were stained with hematoxylin for 30 min at 25°C, washed with water
for 20 min, stained with eosin for 5 min at 25°C, dehydrated with
100, 95, 80 and 70% ethanol for 5 min each, cleared using xylene
and mounted with neutral balsam.
For immunohistochemical assessment, the protocol was
performed as described previously (18). In brief, subsequent to
deparaffinization, blocking with 10% goat serum at 25°C for 15 min,
the 4-µm-thick sections were incubated with primary antibodies
(1:400 rabbit anti-HIF-1α and 1:1,000 anti-Caspase-3) in 1% bovine
serum albumin, 0.3% triton X-100 and 0.1% sodium azide in phosphate
buffered saline (PBS) at 4°C overnight, washed three times for 3
min each with PBS, and subsequently incubated with goat anti-rabbit
antibody conjugated to horseradish peroxidase (cat. no. 18149A01,
OriGene Technologies, Inc., Beijing, China) diluted 1:300 in bull
serum albumin blocking buffer for 2 h at room temperature, followed
by use of a 3,3′-diaminobenzidine (DAB) kit according to the
manufacturers' protocol.
Finally, images were acquired using a Leica
microscopic imaging system (cat. no. DM1000; Leica Microsystems
GmbH, Wetzlar, Germany) using a CX22 light microscope with ×200
magnification for positive stains (Olympus Corporation, Tokyo
Japan) to record the lesions in the CA1, CA2, CA3 and dentate gyrus
(DG) regions of the hippocampus. The levels of HIF-1α and caspase-3
in the CA1 region were scored using the Image-Pro Plus software
version 6.0 (Media Cybernetics, Inc., Rockville, MD, USA). The
experiment was repeated three times.
Western blot analysis
Frozen brains were homogenized in cold whole cell
lysis buffer (cat. no. AR0101-30; Wuhan Boster Biological
Technology, Ltd., Wuhan, China) and centrifuged at 12,000 × g for
10 min at 4°C. The total protein concentration was determined using
a BCA protein assay kit according to the manufacturer's protocol.
Subsequent to mixing with the loading buffer and boiling at 100°C
for 5 min, 40 µg protein per lane were separated using 10%
SDS-PAGE.
Proteins were electrophoretically transferred onto
polyvinylidene difluoride membranes and then incubated with the
corresponding primary antibodies overnight at 4°C subsequent to
blocking using 5% skim milk for 1 h at room temperature. The
primary antibodies included Apaf-1 (1:300), HIF-1α (1:1,000),
β-actin (1:1,000), caspase-3 and cleaved caspase-3 (1:1,000), Bax
(1:300) and cyto-c (1:200) overnight at 4°C. Membranes were washed
with PBS with 0.1% Tween-20 and subsequently incubated with
horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G
secondary antibodies (1:1,000; cat. no. GB23303; Wuhan Goodbio
Technology Co., Ltd., Wuhan, China) for 2 h at room temperature.
The blots were developed using ultrasignal Western ECL substrate
(4A Biotech Co., Ltd.) and visualized using ChampChemi 610 Plus
(Beijing Sage Creation Science Co., Ltd., Beijing, China). The
relative expression levels of the protein bands of interest were
quantified and normalized to β-actin using Image-Pro Plus software
version 6.0 (Media Cybernetics, Inc.).
Nissl and TUNEL staining
To detect the histomorphological changes in the
hippocampal neurons, Nissl staining was performed as previously
described (19). In brief, the
4-µm sections were incubated with 0.1% toluidine blue for 10 min at
room temperature, rinsed in double distilled water followed by
treatment with 70 and 95% ethanol, dehydrated in 100% ethanol,
cleared using xylene and coated with neutral resin.
Apoptosis was evaluated using a TUNEL apoptosis
detection kit according to the manufacturers' protocol. In brief,
the 5 µm paraffin-embedded brain sections were fully dewaxed and
hydrated, soaked in 3% hydrogen peroxide for 10 min at 25°C, and
washed with distilled water three times for 2 min each time.
Subsequently, the sections were treated with proteinase K solution
at 37°C for 15 min and washed with 0.01 M tris buffered saline
(TBS) thrice for 2 min each time. Then the sections were tagged for
2 h at 37°C with 20 µl mixed labeling buffer (1 µl TdT:1 µl
DIG-dUTP:18 µl labeling buffer) for each section and washed with
0.01 M TBS thrice for 2 min each time. Subsequently, they were
blocked at room temperature for 30 min with 50 µl blocking reagent.
A total of 50 µl biotinylated anti-digoxin antibody (1:100) was
added to the sections at 37°C for 30 min and washed with 0.01 M TBS
thrice for 2 min each time. Then, the sections were incubated with
50 µl streptavidin biotin complexes (1:100) at 37°C for 30 min,
washed with 0.01 M TBS four times for 5 min each time, treated
using DAB at 20°C for 10 min, subjected to mild hematoxylin
restaining at 25°C for 2 min, rinsed in 0.01 M TBS followed with
distilled water, dehydrated in 100% ethanol, cleared using xylene
and coated using neutral resins. Blocking reagent, biotinylated
anti-digoxin antibody, streptavidin biotin complexes were all taken
from apoptosis detection kit I-POD (cat. no. MK1025; Wuhan Boster
Biological Technology, Ltd.) and were used strictly according with
the manufacturer's protocols. All sections were randomly observed
under a CX22 light microscope with a ×200 magnification (Olympus
Corporation). The number of Nissl's bodies and the TUNEL positive
apoptotic cell rate were calculated using Image-Pro Plus software
version 6.0 (Media Cybernetics, Inc.).
Statistical analysis
Statistical analysis was conducted using SPSS 17.0
(SPSS, Inc., Chicago, IL, USA). Results were presented as the mean
± standard deviation (n=6 with replicas for each sample). The
statistical analysis was performed using analysis of variance
followed by a Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Physical behavioral performance
Following 30 min acclimatization in a
multifunctional hypobaric chamber, a hypoxic protocol was conducted
on six rats, while the control group of rats were simultaneously
treated outside of the chamber as a parallel control group. During
the first 5 min, the abnormal activities or behavioral performance
of the rats were observed in the hypoxic group. Briefly, an
increased activity frequency, increased respiratory rate, head-up
and neck extension, hind legs straight and nasal flaring allegro
were observed in the hypoxic group compared with the control group.
In addition, the physical activities reduced gradually as the
hypoxic time extended. Following 0.5 h, all the rats stood still,
huddled together, trembled and fell to one side occasionally.
Ultimately, the semiclosed eyes and extremely fast belly breathing
continued until the end of the hypoxic treatment. However, no
abnormal activities or behavioral performance were observed in the
control group. No rats succumbed to mortality either among the
control or hypoxic group in a 24-h hypoxic duration at 9,000 m.
Hence, the present hypoxic protocol that was applied is able to
induce abnormal activities or affect the behavioral performance of
rats.
Effect of hypoxia on oxidative stress
indicators in serum
The antioxidant activities of MDA, GSSG, LDH and
reduced SOD along with GSH in the rats stimulated by hypobaric
hypoxia are summarized in Table I.
The results revealed significantly higher levels of serum MDA, LDH
and GSSG (P<0.01), and significantly lower levels of serum SOD
(P<0.05) and GSH (P<0.01) in the model group compared with
that in the control group. The statistical results are presented in
Fig. 2.
 | Table I.MDA, GSH, GSSG, SOD and LDH
activities in the serum of rats (mean ± standard deviation,
n=6). |
Table I.
MDA, GSH, GSSG, SOD and LDH
activities in the serum of rats (mean ± standard deviation,
n=6).
| Parameter | Control group | Model group |
|---|
| SOD (U/ml) | 25.38±0.69 |
21.65±0.14a |
| MDA (nmol/ml) | 12.91±0.40 |
26.12±1.62b |
| GSH (µmol/l) | 10.89±0.30 |
1.74±0.40b |
| GSSG (µmol/l) | 1.79±0.43 |
6.21±0.67b |
| LDH (U/l) | 572.77±30.66 |
872.50±37.21b |
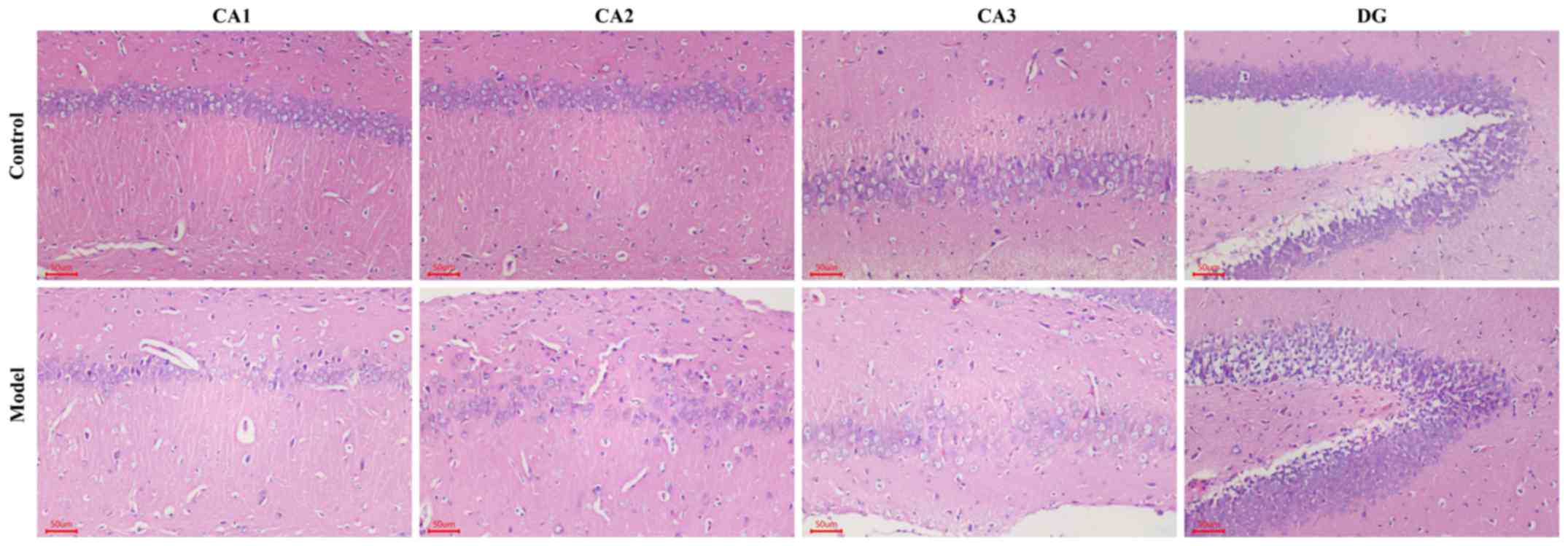
Hypobaric hypoxia induces neuronal
damage in the hippocampus of rats
H&E staining revealed that the nerve cell
arrangement was well organized in the hippocampus CA1, CA2, CA3 and
DG regions in the control group. Conversely, a remarkable neuron
loss, cellular swelling, widened pericellular spaces and shrunken
neurons with darkly stained pyknotic nuclei in the hippocampi and
cortex following hypoxic treatment were observed in the hypoxic
group (Fig. 3). As presented in
Fig. 4A and B, the number of
surviving neurons stained by Nissl's presented a significantly
decreased integrated optical density (IOD) in the CA1 region in
hypobaric hypoxic rats compared with that in the control group
(P<0.01). In addition, the TUNEL positive apoptotic cell rate
was significantly higher in the hypoxic group compared with that in
the control group (P<0.01).
Hypoxia enhances the expression of
Apaf-1, HIF-1α, caspase-3, cleaved caspase-3, Bax and cyto-c
The HIF-1α and apoptosis-associated protein
expression levels are presented in Figs. 4 and 5. Western blot analysis revealed that the
protein expression levels of Apaf-1, HIF-1α, caspase-3, cleaved
caspase-3, Bax and cyto-c in the model group were significantly
higher compared with that in the control group (P<0.01; Fig. 4C and D). Significantly stronger
immunopositive caspase-3 and HIF-1α levels were also observed in
the model group compared with that in the control group (P<0.01;
Fig. 5).
Discussion
Behavioral investigation in animals has revealed
that abnormal activity performances are associated with brain
damage (20–22). A previous study has revealed a
series of affected physical activities in rats subjected to middle
cerebral artery occlusion and reperfusion injury (23). In the present experiment,
hypoxia-associated abnormal performances, including nasal flaring
allegro, head-up and neck extension, hind legs straightening and an
increased respiratory rate were also presented in
hypobaric-hypoxia-induced rat brain injuries. During the hypoxic
period, biomarkers in the serum or injured tissues including
increased MDA, GSSG and LDH levels and decreased SOD and GSH levels
have been demonstrated to alter substantially (24–27).
The results of the present study indicate an elevation of MDA, GSSG
and LDH levels in the serum compared with that in the control
group, whereas SOD and GSH levels decreased in the model. The
results are presented in Table I
and Fig. 2. Therefore, this
indicates that the buffering system or redox enzyme in serum may be
used for diagnosis of early brain injury.
Mounting evidence suggests that hypoxic or ischemic
insult may induce morphological deterioration in the rat
hippocampus, including sparse cell arrangement in the pyramidal
layer of the hippocampus and karyopyknosis at the microscale
(28–30). The present results of H&E
staining (Fig. 3) of the rat
hippocampus revealed similar pathological phenomena in the CA1,
CA2, CA3 and DG in the hypoxic rat hippocampus. In addition,
substantially decreased quantities and sizes of Nissl bodies in the
hippocampus were observed in the hypoxic group, which were
consistent with the results of previous studies (Fig. 4C) (31–33).
HIFs, which consist of HIF-1 and HIF-2 isoforms,
respond to the prevailing level of oxygen. Functioning as a master
switch in the human response to hypoxia, a substantially higher
protein expression of HIF-1α is produced during hypoxia.
Subsequently, it translocates from the cytoplasm to the nucleus
(34–35). Finally, the HIF-1α protein binds to
specific DNA sequences of downstream targets, thereby controlling
the transcription rate of >200 genes (36–39).
Zheng et al (40) reported
that salidroside induced the inhibition of HIF-1α protein
degradation rather than its gene overexpression. As a result,
HIF-1α protein expression tended to increase. However, a higher
gene expression of HIF-1α stimulated by hypoxia in addition to
salidroside was confirmed previously (41,42).
Thus, HIF-1α may be viewed as a notable molecule that may be
affected by drugs or oxygen concentration, while the affected
biosynthesis or degradation of the HIF-1α protein in vivo
requires further investigation. In the present study, the increased
accumulation of the HIF-1α protein was detected by western blot
analysis in the hypoxic group compared with that in the control
group. Furthermore, it was previously reported that hypoxia or
ischemic brain injuries induce a decrease of mitochondrial membrane
potential (43). Increased protein
levels of cyto-c, Bax and Apaf-1 were also demonstrated in the
present study. Thus, the apoptotic body consisting of Apaf-1,
cyto-c and caspase-9 in the cytoplasm accelerates the process of
cell apoptosis through activating the caspase-dependent apoptotic
signaling pathway (44). A graphic
illustration of the involved molecular mechanism is presented in
Fig. 6. The protein expression
levels of HIF-1α, caspase-3 and cleaved caspase-3 were assessed
using immunohistochemistry and western blot analysis in the present
study. The results are presented in Figs. 4C-D and 5. Cyto-c, Bax and Apaf-1 protein
expression levels were consistently increased in the hypoxic group
compared with that in the control group; the results are presented
in Fig. 5B. Furthermore, TUNEL
staining was used to evaluate whether hypobaric hypoxia induced
neuronal apoptosis, as presented in Fig. 4B, a large number of positive
staining apoptotic cells appeared in hippocampal neurons of the
hypoxic group.
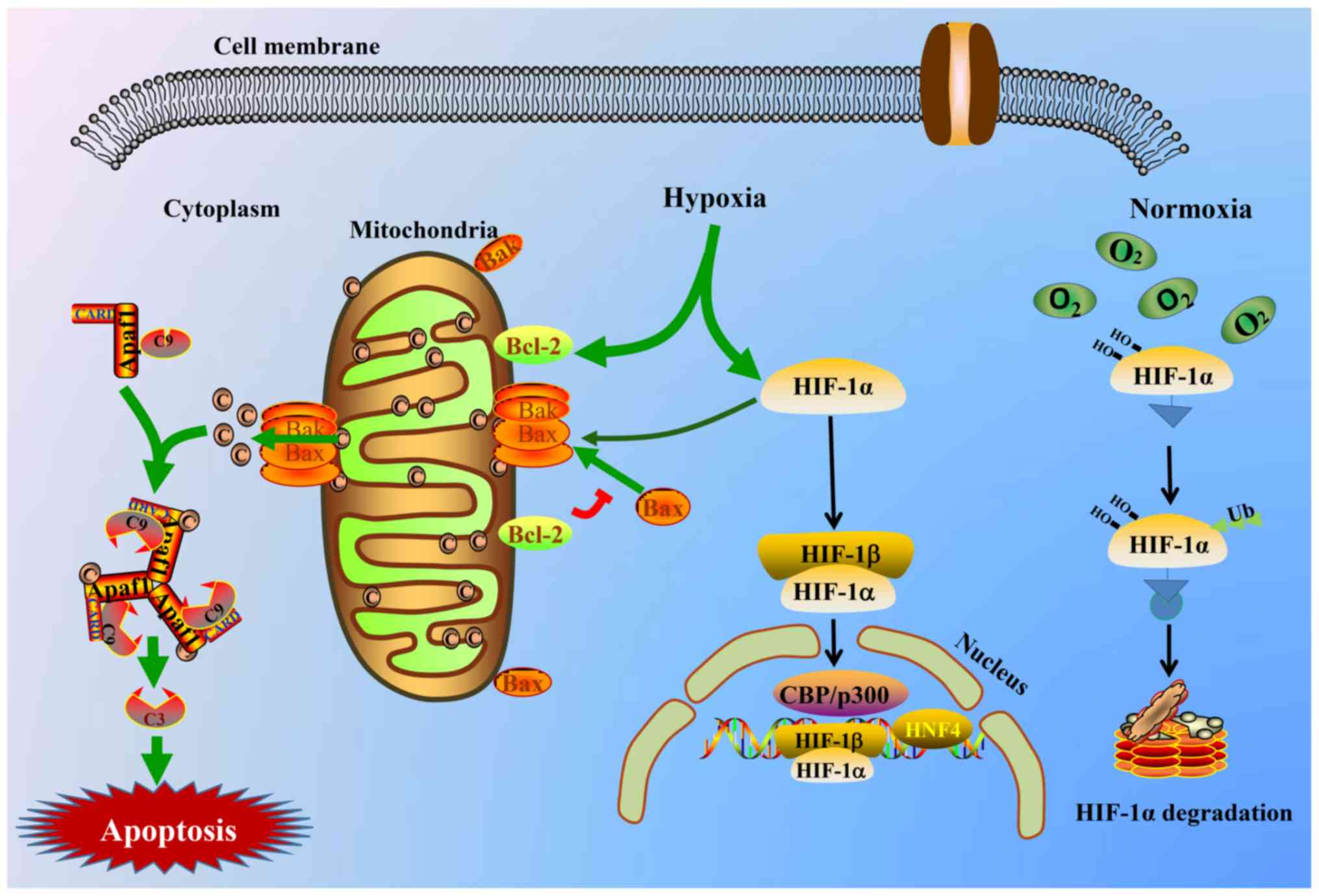 | Figure 6.Schematic view of HIF-1α and the
intrinsic triggering pathway of the apoptotic signaling cascade.
Hypoxia stimulates the HIF-1α and mitochondrial apoptotic signaling
pathway and triggers the release of cyto-c from the mitochondrial
matrix to the cytoplasm, thereby binding to the Apaf-1 protein,
inducing its conformational change and activating it. Thus, the
apoptosome consisting of cyto-c/Apaf-1/caspase-9 is yielded and is
able to cleave or activate the executioner caspase-3. The activated
caspase-3 complex then results in apoptosis. C9, caspase-9; C3,
caspase-3; HIF-1α, hypoxia inducible factor-1α; cyto-c, cytochrome
c; Apaf-1, apoptotic protease activating factor-1; CARD, caspase
recruitment domain containing; Bak, Bcl2 antagonist/killer; Bax,
Bcl-2-associated X protein; Bcl-2, Bcl2, apoptosis regulator; CBP,
CREB binding protein; HIF-1β, hypoxia inducible factor-1β; HNF4,
hepatocyte nuclear factor 4α. |
In conclusion, a hypobaric hypoxia brain injury rat
model induced by a multifunctional hypobaric chamber was
established successfully at 9,000 m for 24 h. The redox enzymes in
the serum, HIF-1α and mitochondrial apoptosis-associated proteins,
along with H&E and Nissl's staining, may be applied to evaluate
the degree of a hypoxic injury. This simulated high-altitude
hypoxia brain injury model in SD rats would be beneficial for
progressing high-altitude hypoxia drug discovery. In addition, the
increased HIF-1α protein expression induced by high-altitude
hypoxia may stimulate the expression of the microRNA-210 gene,
thereby inhibiting the expression of iron-sulfur cluster assembly
enzyme 1/2 and COX10, Heme A:Farnesyltransferase cytochrome
c oxidase assembly factor, which affect the process of
mitochondrial energy metabolism (45,46).
Furthermore, the hypoxic-activated
phosphoinositide-3-kinase/protein kinase B signaling pathway may
also increase HIF-1α overexpression, and bind to the hypoxia
response element in the nucleus and trigger downstream target gene
expression, including vascular endothelial growth factor,
erythropoietin, heme oxygenase l, inducible nitric oxide synthase
and nuclear factor-κB (47). Thus,
it is reasonable to believe that HIF-1α is a core component of the
body's response to hypoxia. The degree and duration of hypoxia
determine its protein synthesis and degradation. The association
between HIF-1α and hypoxia, and additional treatment strategies
urgently require further investigation and development.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81203000), National
Natural Science Foundation of China (grant no. 81403187), State
Administration of Traditional Chinese Medicine of the People's
Republic of China, Special Scientific Research in the Traditional
Chinese Medicine field (grant no. 201507002) and Supported by
Sichuan Science and Technology Program (grant no. 19YYJC1922).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YH designed the present study, performed the animal
experiments, analyzed the data and wrote the manuscript. XW, XC and
JZ performed the animal experiments, the physiological test and the
H&E staining. XA, TK and YH analyzed the data, organized the
images and improved the language. YL and YY performed the protein
detection and immunohistochemical analysis. YZ and XM performed
serum biochemical indexes detection, Nissl's and TUNEL staining.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The protocol of this investigation was ethically
approved by the Animal Research Ethics Committee of Chengdu
University of Traditional Chinese Medicine (Chengdu, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Young AJ, Berryman CE, Kenefick RW,
Derosier AN, Margolis LM, Wilson MA, Carrigan CT, Murphy NE,
Carbone JW, Rood JC, et al: Altitude acclimatization alleviates the
hypoxia-induced suppression of exogenous glucose oxidation during
steady-state aerobic exercise. Front Physiol. 9:8302018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gonzalez Garay A, Molano Franco D, Nieto
Estrada VH, Martí-Carvajal AJ and Arevalo-Rodriguez I:
Interventions for preventing high altitude illness: Part 2. Less
commonly-used drugs. Cochrane Database Syst Rev.
3:CD0129832018.PubMed/NCBI
|
|
3
|
Paul S, Gangwar A, Bhargava K, Khurana P
and Ahmad Y: Diagnosis and prophylaxis for high-altitude
acclimatization: Adherence to molecular rationale to evade
high-altitude illnesses. Life Sci. 203:171–176. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xiang K, Ouzhuluobu, Peng Y, Yang Z, Zhang
X, Cui C, Zhang H, Li M, Zhang Y, Bianba, et al: Identification of
a Tibetan-specific mutation in the hypoxic gene EGLN1 and its
contribution to high-altitude adaptation. Mol Biol Evol.
30:1889–1898. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang H, He Y, Cui C, Ouzhuluobu,
Baimakangzhuo, Duojizhuoma, Dejiquzong, Bianba, Gonggalanzi, Pan Y,
et al: Cross-altitude analysis suggests a turning point at the
elevation of 4,500 m for polycythemia prevalence in Tibetans. Am J
Hematol. 92:E552–E554. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gonggalanzi, Labasangzhu, Nafstad P,
Stigum H, Wu T, Haldorsen ØD, Ommundsen K and Bjertness E: Acute
mountain sickness among tourists visiting the high-altitude city of
Lhasa at 3658 m above sea level: A cross-sectional study. Arch
Public Health. 74:232016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
West JB: High-altitude medicine. Lancet
Respir Med. 3:12–13. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Basnyat B and Murdoch DR: High-altitude
illness. Lancet. 361:1967–1974. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Azad P, Stobdan T, Zhou D, Hartley I,
Akbari A, Bafna V and Haddad GG: High-altitude adaptation in
humans: From genomics to integrative physiology. J Mol Med (Berl).
95:1269–1282. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Davis C and Hackett P: Advances in the
prevention and treatment of high altitude illness. Emerg Med Clin
North Am. 35:241–260. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Horscroft JA, Kotwica AO, Laner V, West
JA, Hennis PJ, Levett DZH, Howard DJ, Fernandez BO, Burgess SL,
Ament Z, et al: Metabolic basis to Sherpa altitude adaptation. Proc
Natl Acad Sci USA. 114:6382–6387. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ward PA, Blanchard RJ and Bolivar V:
Recognition and alleviation of distress in laboratory animals.
National Academies Press. 44:3802007.
|
|
13
|
Zhang Y, Meng X, Wu W, Lai X, Wang Y,
Zhang J and Wang Z: Duoxuekang, a traditional Tibetan medicine,
reduces hypoxia-induced high-altitude polycythemia in rats. A
Sponsored Supplement to Science. 338:63–64. 2012.
|
|
14
|
Zhang XY, Zhang XJ, Xv J, Jia W, Pu XY,
Wang HY, Liang H, Zhuoma-Lamao and Lu DX: Crocin attenuates acute
hypobaric hypoxia-induced cognitive deficits of rats. Eur J
Pharmacol. 818:300–305. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gong G, Yin L, Yuan L, Sui D, Sun Y, Fu H,
Chen L and Wang X: Ganglioside GM1 protects against high altitude
cerebral edema in rats by suppressing the oxidative stress and
inflammatory response via the PI3K/AKT-Nrf2 pathway. Mol Immunol.
95:91–98. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Song TT, Bi YH, Gao YQ, Huang R, Hao K, Xu
G, Tang JW, Ma ZQ, Kong FP, Coote JH, et al: Systemic
pro-inflammatory response facilitates the development of cerebral
edema during short hypoxia. J Neuroinflammation. 13:63–76. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo P, Luo H, Fan Y, Luo Y and Zhou Q:
Establishment and evaluation of an experimental animal model of
high altitude cerebral edema. Neurosci Lett. 547:82–86. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gu J, Su S, Guo J, Zhu Y, Zhao M and Duan
JA: Anti-inflammatory and anti-apoptotic effects of the combination
of Ligusticum chuanxiong and Radix Paeoniae against focal cerebral
ischaemia via TLR4/MyD88/MAPK/NF-κB signalling pathway in MCAO
rats. J Pharm Pharmacol. 70:268–277. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang P, Xie ZD, Xie CN, Lin CW, Wang JL,
Xuan LN, Zhang CW, Wang Y, Huang ZH and Teng HL: AMP-activated
protein kinase-dependent induction of autophagy by erythropoietin
protects against spinal cord injury in rats. CNS Neurosci Ther.
24:1185–1195. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Song H, Konan LM, Cui J, Johnson CE,
Langenderfer M, Grant D, Ndam T, Simonyi A, White T, Demirci U, et
al: Ultrastructural brain abnormalities and associated behavioral
changes in mice after low-intensity blast exposure. Behav Brain
Res. 347:148–157. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kaplan GB, Leite-Morris KA, Wang L,
Rumbika KK, Heinrichs SC, Zeng X, Wu L, Arena DT and Teng YD:
Pathophysiological bases of comorbidity: Traumatic brain injury and
post-traumatic stress disorder. J Neurotrauma. 35:210–225. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mancuso M, Abbruzzese L, Canova S, Landi
G, Rossi S and Santarnecchi E: Transcranial random noise
stimulation does not improve behavioral and neurophysiological
measures in patients with subacute vegetative-unresponsive
wakefulness state (VS-UWS). Front Hum Neurosci. 11:5242017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao Q, Wang X, Chen A, Cheng X, Zhang G,
Sun J, Zhao Y, Huang Y and Zhu Y: Rhein protects against cerebral
ischemic-/reperfusion-induced oxidative stress and apoptosis in
rats. Int J Mol Med. 41:2802–2812. 2018.PubMed/NCBI
|
|
24
|
Ren H, Meng X, Yin J, Sun J, Huang Q and
Yin Z: Ganoderma lucidum polysaccharide peptide attenuates skin
flap ischemia-reperfusion injury in a thioredoxin-dependent manner.
Plast Reconstr Surg. 142:23e–33e. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang X, Rui L, Wang M, Lian H and Cai L:
Sinomenine attenuates chronic intermittent hypoxia-induced lung
injury by inhibiting inflammation and oxidative stress. Med Sci
Monit. 24:1574–1580. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang G, Min D, Yan J, Yang M and Lin G:
Protective role and mechanism of snakegourd peel against myocardial
infarction in rats. Phytomedicine. 42:18–24. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu H, Luo D, Li C, Zhang H, Shunxian A,
Zhang Y and Sun C: Chicoric acid improves heart and blood responses
to hypobaric hypoxia in tibetan yaks. Am J Chin Med. 46:339–355.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yan YT, Li SD, Li C, Xiong YX, Lu XH, Zhou
XF, Yang LQ, Pu LJ and Luo HY: Panax notoginsenoside saponins Rb1
regulates the expressions of Akt/mTOR/PTEN signals in the
hippocampus after focal cerebral ischemia in rats. Behav Brain Res.
345:83–92. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ramli Y, Alwahdy AS, Kurniawan M, Juliandi
B, Wuyung PE and Susanto YDB: Intravenous versus intraarterial
transplantation of human umbilical cord blood mononuclear cells for
brain ischemia in rats. Hayati J Biosci. 24:187–194. 2017.
View Article : Google Scholar
|
|
30
|
Sun J, Wang F, Li H, Zhang H, Jin J, Chen
W, Pang M, Yu J, He Y, Liu J and Liu C: Neuroprotective effect of
sodium butyrate against cerebral ischemia/reperfusion injury in
mice. Biomed Res Int. 2015:3958952015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zuo D, Lin L, Liu Y, Wang C, Xu J, Sun F,
Li L, Li Z and Wu Y: Baicalin attenuates ketamine-induced
neurotoxicity in the developing rats: Involvement of PI3K/Akt and
CREB/BDNF/Bcl-2 pathways. Neurotox Res. 30:159–172. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu Z, Wu P, Zuo X, Yu N, Qin Y, Xu Q, He
S, Cen B, Liao W and Ji A: LncRNA-N1LR enhances neuroprotection
against ischemic stroke probably by inhibiting p53 phosphorylation.
Mol Neurobiol. 54:7670–7685. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang JZ, Jing L, Ma Y, Guo FY, Chang Y
and Li PA: Monosialotetrahexosy-1 ganglioside attenuates diabetes-
enhanced brain damage after transient forebrain ischemia and
suppresses phosphorylation of ERK1/2 in the rat brain. Brain Res.
1344:200–208. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bahrami A, Atkin SL, Majeed M and Sahebkar
A: Effects of curcumin on hypoxia-inducible factor as a new
therapeutic target. Pharmacol Res. 137:159–169. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gonzalez FJ, Xie C and Jiang C: The role
of hypoxia-inducible factors in metabolic diseases. Nat Rev
Endocrinol. 15:21–32. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Choudhry H and Harris AL: Advances in
hypoxia-inducible factor biology. Cell Metab. 27:281–298. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
West JB: Physiological effects of chronic
hypoxia. N Engl J Med. 376:1965–1971. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee KE and Simon MC: SnapShot:
Hypoxia-inducible factors. Cell. 163:1288–1288.e1. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
West JB: High altitude medicine. Am J
Respir Crit Care Med. 186:1229–1237. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zheng KY, Zhang ZX, Guo AJ, Bi CW, Zhu KY,
Xu SL, Zhan JY, Lau DT, Dong TT, Choi RC and Tsim KW: Salidroside
stimulates the accumulation of HIF-1α protein resulted in the
induction of EPO expression: A signaling via blocking the
degradation pathway in kidney and liver cells. Eur J Pharmacol.
679:34–39. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Long Z, Li J and Meng X: The protective
effects of active ingredients isolated from Rhodiola on the
myocardial cell damage induced by hypoxia/glucose deprivation.
Pharmacology and Clinics of Chinese Materia Medica. 26:24–25.
2010.(In Chinese).
|
|
42
|
Wang Y, Zhang Y, Feng X, Meng X, Long Y
and Zhang J: Bioactive chemical constituents from Rhodiola
crenulata and effect on hypoxia inducible factor-1α expression.
West China J Pharmaceutical Sci. 24:021–024. 2009.(In Chinese).
|
|
43
|
Hu Y, Lv X, Zhang J and Meng X:
Comparative study on the protective effects of salidroside and
hypoxic preconditioning for attenuating anoxia-induced apoptosis in
pheochromocytoma (PC12) cells. Med Sci Monit. 22:4082–4091. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao Q, Cheng X, Wang X, Wang J, Zhu Y and
Ma X: Neuroprotective effect and mechanism of Mu-Xiang-You-Fang on
cerebral ischemia-reperfusion injury in rats. J Ethnopharmacol.
192:140–147. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Qiu J, Zhou XY, Zhou XG, Cheng R, Liu HY
and Li Y: Neuroprotective effects of microRNA-210 against
oxygen-glucose deprivation through inhibition of apoptosis in PC12
cells. Mol Med Rep. 7:1955–1959. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
He M, Lu Y, Xu S, Mao L, Zhang L, Duan W,
Liu C, Pi H, Zhang Y, Zhong M, et al: MiRNA-210 modulates a
nickel-induced cellular energy metabolism shift by repressing the
iron-sulfur cluster assembly proteins ISCU1/2 in Neuro-2a cells.
Cell Death Dis. 5:e10902014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ye Z, Guo Q, Xia P, Wang N, Wang E and
Yuan Y: Sevoflurane postconditioning involves an up-regulation of
HIF-1α and HO-1 expression via PI3K/Akt pathway in a rat model of
focal cerebral ischemia. Brain Res. 1463:63–74. 2012. View Article : Google Scholar : PubMed/NCBI
|