Introduction
Breast cancer is the most common cancer in women,
and its incidence shows an increasing trend globally (1–3). Annually,
~1,400,000 women are diagnosed with breast cancer worldwide, of
which there are 500,000 mortalities. The incidence of breast cancer
in China has also increased rapidly, and is a huge threat to the
health of Chinese women (4,5). At present, the treatment of breast
cancer predominantly involves surgery combined with chemotherapy,
endocrine therapy, targeted therapy or immunotherapy, which has
shown good curative effects; however, the current treatment
strategy has not shown good therapeutic effects for all
patients.
Gene therapy is the most promising potential
treatment option for cancer, and is currently undergoing rapid
development. Selecting appropriate therapeutic genes is the first
stage in the development of gene therapy. Various genes have been
investigated for their potential use in gene therapy, including
genes that allow ‘immortalization’ of cells, which is a
prerequisite for tumor formation, and genes that promote or induce
tumor cell apoptosis (6). Apoptin,
which is derived from the chicken anemia virus (CAV), is a highly
conservative protein that has no homology with other animal or
viral sequences (7). Due to its
ability to selectively induce apoptosis in a large panel of human
malignant cells, without affecting normal cells, it has emerged as
an important potential agent for cancer gene therapy (8).
An efficient gene delivery method is crucial for the
implementation of gene therapy. Nonviral gene delivery systems,
which involve using natural or synthetic proteins as delivery
tools, have become increasingly popular due to their advantages,
including safety, stability and relative cheapness (9). Human serum albumin (HSA) is the most
abundant protein in serum, and is the main driving force of the
circulatory system. HSA is also an important source of nutrition
for the cells of the body. As tumor development requires a large
volume of nutrients, HSA has been shown to accumulate in solid
tumors (10). Rhaese et al
(11) successfully constructed a
HSA-DNA complex, and assessments of its stability, cytotoxicity,
nuclease resistance and transfection and expression efficiency
demonstrated that HSA may be an ideal gene delivery tool.
The present study developed a gene therapy approach
incorporating the delivery of the cancer-specific cytotoxic
protein, apoptin, with HSA as a vector for the treatment of breast
cancer. The HSA-polyethylenimine (PEI)-pcDNA-Apoptin construct was
transfected into the MCF-7 breast cancer cell line, and the in
vitro and in vivo effects on MCF-7 cell viability and
apoptosis were determined. When systemically delivered into mice,
the HSA-PEI-pcDNA-Apoptin construct was capable of inducing
apoptosis and causing significant tumor regression, in the absence
of damage to normal tissues.
Materials and methods
Cells and reagents
The MCF-7 breast cancer cell line was obtained from
the Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). MCF-7 cells were cultured in RPMI-1640 medium
supplemented with 10% fetal bovine serum, 100 U/ml penicillin and
100 µg/ml streptomycin (complete medium; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in 5% CO2.
HSA and PEI were purchased from Sigma-Aldrich; Merck Millipore
(Darmstadt, Germany). The enhanced green fluorescent protein
(EGFP)-N1 plasmid (pEGFP-N1) was a gift from Professor Xinhan Zhao
of Xi'an Jiaotong University (Xi'an, China). The study was approved
by the Ethics Committe of Xi'an Jiaotong University.
Construction of the expression
vector
The expression vectors containing apoptin or EGFP
were generated under the control of the cytomegalovirus promoter.
The apoptin gene was synthesized by Shanghai GenePharma Co., Ltd.
(Shanghai, China) based on sequences in GenBank (accession no.
Ay171617), and amplified by polymerase chain reaction (PCR) using
Taq DNA polymerase (Sigma-Aldrich; Merck Millipore). The
primers were: Sense, 5′-ACGAATTCCGATGAACGCTCTCCAAGAAGATACTC-3′ and
antisense, 5′-CAGGATCCGTFGACAGTCTFATACACCTTCTTGCG-3′. The terminals
of the primers were used to introduce the Eco RI and
Xba I excision sites. PCR was performed with an initial
denaturation step at 94°C for 5 min, followed by 35 cycles of
denaturation at 94°C for 30 sec, annealing at 56°C for 45 sec and
extension at 72°C for 45 sec. A final extension step at 72°C for 10
min was performed. The PCR products were separated by agarose gel
electrophoresis and the bands were purified from the gels. The PCR
fragments were cloned into pEGFP-N1 using the T4 DNA ligase
(Sigma-Aldrich; Merck Millipore). Subsequently, the insert site was
released from pEGFP-N1 by digestion with Eco RI and
Xba I restriction enzymes (Sigma-Aldrich; Merck Millipore),
and PCR fragments were cloned into pEGFP-N1 using the T4 DNA
ligase; the complex was termed pEGFP-N1-Apoptin, which was
amplified in bacterial cells (E. coli DH5α; Takara Bio,
Inc., Otsu, Japan). The cells were transfected with
pEGFP-N1-Apoptin as follows: Following thawing of competent cells,
2 µl pEGFP-N1-Apoptin was added to 100 µl competent cells and
incubated on ice for 30 min. This was followed by incubation in
water for 45–60 sec at 42°C, then incubation on ice for 2 min,
vortexing for 1 h at 37°C after added 900 µl SOC media. Finally,
100 µl bacteria liquid was added to the Luria-Bertani (LB) media,
which contained 50 µg/ml kanamycin, and was incubated for 12–16 h
at 37°C so that bacterial colonies would appear and select for
positive clones. The pcDNA-EGFP vector was constructed by excising
EGFP from the pEGFP-N1 plasmid using Eco RI/Xba I
restriction enzymes and sub-cloning into the Eco RI and
Xba I sites of pcDNA3.1 (Sigma-Aldrich; Merck Millipore).
The pcDNA-Apoptin vector was constructed by subcloning the apoptin
fragment from pEGFP-Apoptin into pcDNA3.1.
Construction of recombinant
HSA-PEI-pcDNA-Apoptin
HSA (10 mg) was dissolved in 0.05 mol/l
2-(N-Morpholino) ethanesulfonic acid (MES) and 0.05 mol/l NaCl
(MES/NaCl) buffer solution (pH 4). Subsequently, 300 mg
1-ethyl-3-(3-dimethyllaminopropyl)carbodiimide hydrochloride and
0.06 mol/l N-hydroxysuccinimide (NHS) were dissolved in MES/NaCl
buffer to form an NHS-HSA ester. PEI (10 mg) was reacted with
NHS-HSA solution at room temperature, and stirred for 20 h using a
magnetic stirring apparatus. The HiTrap SP FF ion exchange column
reagent kit (GE Healthcare Life Sciences, Shanghai, China) was used
to purify the synthesized HSA-PEI according to the manufacturer's
protocols. Finally, 10 µg HSA-PEI and 10 µg pcDNA-Apoptin or pcDNA
were dissolved in 100 ml pure water. The recombinant
HSA-PEI-pcDNA-Apoptin and HSA-PEI-pcDNA constructs were used for
the following experiments. In order to identify the recombinant
plasmid, the recombinant plasmid was digested using EcoRI
and XbaI restriction enzymes, and the products were
separated by 1% agarose gel electrophoresis.
Cell transfection
MCF-7 cells (2×105) were cultured in
24-well culture plates. Upon reaching the logarithmic growth phase,
various concentrations of HSA-PEI-pcDNA-Apoptin (0, 0.1, 0.25, 0.5
or 1.0 mg/ml) were added to the cells. After 24 h, the cells were
stable, and were permeabilized and washed with phosphate-buffered
saline (PBS), after which apoptin expression was examined under a
fluorescence microscope.
Cell viability assay
The MTT assay (Sigma-Aldrich; Merck Millipore) was
performed to detect cell viability following the transfection of
MCF-7 cells with the HSA-PEI-pcDNA-Apoptin construct. MCF-7 cells
were cultured in 96-well plates at a density of 4×104/ml
(200 µl per well) for 24 h at 37°C. Each plate was divided into
three groups, including the HSA-PEI-pcDNA-Apoptin group, the
HSA-PEI-pcDNA group and the blank control group. Subsequently,
HSA-PEI-pcDNA-Apoptin, HSA-PEI-pcDNA and 0.9% NaCl were added to
the appropriate wells and, after 24 h, 20 µl MTT (5 mg/ml) was
added for 4 h at 37°C. The culture media was removed and the
crystals were dissolved by addition of 150 µl dimethylsulfoxide per
well, and oscillating for 15 min at room temperature. The
absorbance of the reaction solution was measured at 490 nm using a
microplate reader (Multiskan FC; Thermo Fisher Scientific, Inc.).
The inhibition rate of cell growth was calculated as follows:
Inhibition rate (%) = (1 - absorbance of experimental group /
absorbance of control group) × 100.
Flow cytometric analysis
The effect of HSA-PEI-pcDNA-Apoptin on cell
apoptosis was determined by flow cytometry using the Annexin
V-phycoerythrin (PE)/7-aminoactinomycin D (7-AAD) Apoptosis kit
(Genechem, Co., Ltd., Shanghai, China), according to the
manufacturer's protocol. Briefly, MCF-7 cells were rinsed twice
using PBS, digested with trypsin and centrifuged at 400 × g
for 5 min. The cells were incubated with 50 µl Binding Buffer and 5
µl 7-AAD for 5–15 min in the dark at room temperature.
Subsequently, 450 µl Binding Buffer and 1 µl Annexin V-PE were
added to the cells for 5–15 min. The samples were analyzed using
the BD FACSAria™ III Cell Sorter (BD Biosciences, San Jose, CA,
USA), and the data were analyzed using FlowJo 7.6.1 software
(FlowJo, LLC, Ashland, OR, USA).
Semi-quantitative reverse
transcription (RT)-PCR
To detect the mRNA expression of apoptin in MCF-7
cells, total RNA was extracted using TRIzol reagent (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. RT-PCR
was performed using the OneStep RT-PCR kit (Thermo Fisher
Scientific, Inc.) and the following manually designed PCR primers:
Apoptin sense, 5′-CGGCCGGCGTGGGATGAACGATCTCCAAGAAGATAC-3′ and
antisense, 5′-CAAGCTTCAGTCTTATACGCCTTCTTGCGGTTC-3′; and β-actin
sense, 5′-TGACGGGGTCACCCACACTG-3′ and antisense,
5′-TAGAAGCATTTGCGGTGGAC-3′. The PCR cycling conditions were as
follows: 35 cycles of 94°C for 30 sec, 56°C for 30 sec and 72°C for
30 sec, followed by an extension step at 72°C for 10 min.
Subsequently, the PCR products were separated by 1.5% agarose gel
electrophoresis, with β-actin as an internal marker. Images of the
gel were captured, and the apoptin gene was analyzed, using the
GeneGenius Bio Imaging system (Syngene, Frederick, MD, USA).
Western blot analysis
MCF-7 cells were washed three times with cooled PBS
and lysed using radioimmunoprecipitation assay lysis buffer (Xi'an
Heart Biotechnology, Co., Ltd., Xi'an, China) containing 2 mg/ml
protease inhibitors for 30 min. The lysates were then centrifuged
at 25,000 × g for 20 min at 4°C, followed by boiling in
loading buffer for 5 min. Total protein concentrations were
determined using a Bradford protein assay (Sigma-Aldrich; Merck
Millipore), and 150 µg protein samples were separated by 10%
SDS-PAGE and transferred onto polyvinylidene difluoride membranes.
The membranes were blocked using 5% (w/v) skimmed milk powder
(Inner Mongolia Yili Industrial Group, Co., Ltd., China) dissolved
in Tris-buffered saline containing 0.1% Tween-20 (TBS-T) for 1 h at
room temperature. The membranes were incubated with anti-apoptin
(1:1,000 dilution; catalog no. ab193612; Abcam, Cambridge, MA, USA)
and anti-β-actin (1:1,000 dilution; catalog no. sc-47778; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA) antibodies at 4°C
overnight. After washing three times with TBS-T, the membranes were
incubated with the goat anti-mouse IgG1-HRP (1:10,000 dilution;
catalog no. sc-2060; Santa Cruz Biotechnology, Inc.) and goat
anti-rabbit IgG-HRP (1:5,000 dilution; catalog no. sc-2030; Santa
Cruz Biotechnology, Inc.) secondary antibodies for 2 h at room
temperature. The membranes were then washed with TBS-T three times
for 10 min, then either applied with enhanced chemiluminescence
reagents (Merck Millipore), according to the manufacturer's
protocols, and exposed to X-ray film, or applied with DAB (from
which brown staining was visualized). Autoradiogram signals were
quantified using a gel densitometric scanning program. The relative
expression of protein was determined from the optical density ratio
of the corresponding protein bands, as quantified using Quantity
One 4.6.2 software (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Experiments on a nude mouse model in
vivo
A total of 15 BALB/c nu/nu athymic nude female mice
(4–5 weeks-of-age) were purchased from Shanghai Silaike Laboratory
Animal Co., Ltd. (Shanghai, China). The mice were maintained at
Xi'an Jiaotong University in a sterile environment, with a
temperature of 22–23°C, relative humidity of 45–55%, light/dark
cycles of 12/12h, and food and water provided daily, in compliance
with the Institutional Animal Care and Use Committee regulations.
The mice were subcutaneously injected with 1×106 MCF-7
cells (0.1 ml) into the left side of the second breast fat pad.
When the tumors had reached ~100 mm3 in volume, the mice
were randomized into three groups, as follows: i) The
HSA-PEI-pcDN-Apoptin (0.5 mg/ml) group; ii) the HSA-PEI-pcDNA (0.5
mg/ml) group; and iii) the saline (control) group. The mice were
injected (0.5 ml) into the tail vein with HSA-PEI-pcDN-Apoptin,
HSA-PEI-pcDNA or saline once per day, and the tumor volumes were
measured every 3 days. Tumor volumes were calculated using the
equation: Tumor volume (mm3) = length ×
width2 × 0.5. After 25 days, the mice were sacrificed by
cervical dislocation. The liver, lungs, kidneys and tumor tissues
were harvested, fixed in 4% paraformaldehyde solution and embedded
in paraffin. The liver and lung paraffin sections were stained with
hematoxylin and eosin (HE), and were independently evaluated by two
pathologists. The tissue proteins were extracted for western blot
assay.
The handling of animals was in accordance with the
Animal Care Guidelines of Xi'an Jiaotong University of Medical
Sciences Ethical Committee. All the experiments were performed to
minimize the stress and pain experienced by the animals.
Statistical analysis
Statistical analyses were performed using SPSS 13.0
software for Windows (SPSS Inc., Chicago, IL, USA). Two-tailed
Student's t-tests were used for comparisons between two groups.
Data are expressed as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
Successful establishment of the
HSA-PEI-pcDNA-Apoptin expression plasmid
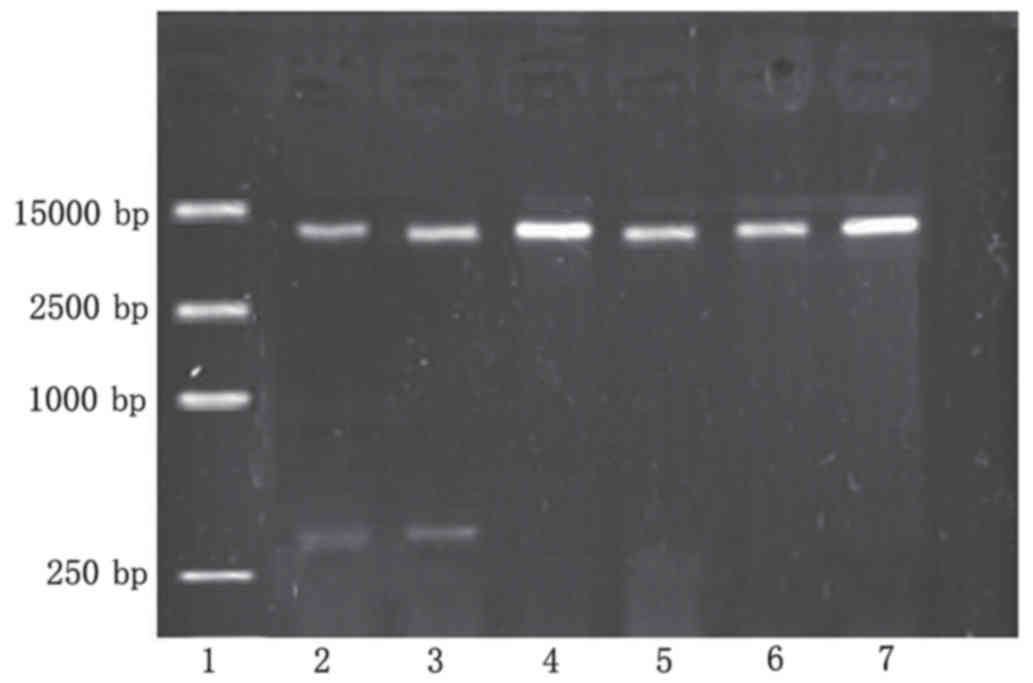
The recombinant plasmid was digested using
Eco RI and Xba I restriction enzymes, and the
products were separated by 1% agarose gel electrophoresis, which
produced two fragments: Apoptin (376 bp) and pEGFP-N1 (5,000 bp)
(Fig. 1). Sequencing by Sigma-Aldrich
(Thermo Fisher Scientific, Inc.) demonstrated that the apoptin gene
sequence of the recombinant plasmid was the same as the apoptin
sequence in GenBank (accession no. Ay171617). The apoptin DNA and
EGFP DNA sequences were shown to form a complete reading frame,
without frameshift mutations, which conformed to the experimental
design.
Apoptin expression in MCF-7 cells
transfected with HSA-PEI-pcDNA-Apoptin
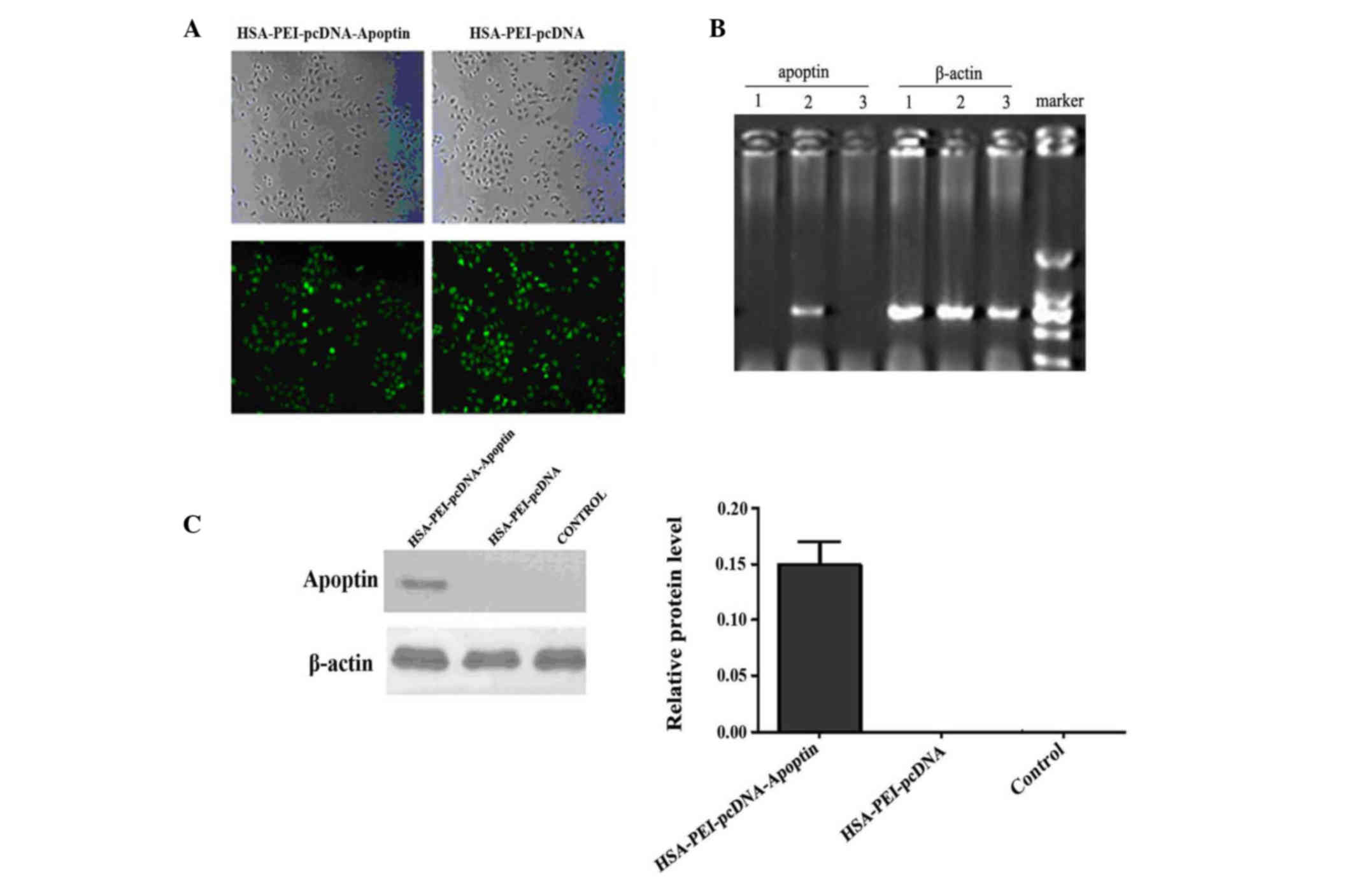
To determine whether HSA-PEI-pcDNA-Apoptin was
successfully transfected into MCF-7 human breast cancer cells, the
cells were transfected with various concentrations of
HSA-PEI-pcDNA-Apoptin or HSA-PEI-pcDNA, and were observed by
fluorescence microscopy after 24 h. Approximately 90% of cells
transfected with 0.5 or 1.0 mg/ml constructs showed positive green
fluorescence staining (Fig. 2A). For
economic reasons, 0.5 mg/ml HSA-PEI-pcDNA-Apoptin was used in the
subsequent in vivo experiments. The mRNA and protein
expression of apoptin in MCF-7 cells was confirmed by
semi-quantitative RT-PCR and western blotting. Apoptin transcripts
were detected in MCF-7 cells at 24 h following transfection.
Apoptin mRNA was only detected in MCF-7 cells transfected with
HSA-PEI-pcDNA-Apoptin (Fig. 2B);
β-actin was an internal marker (Fig.
2C). Western blot analysis confirmed that apoptin was only
expressed in the MCF-7 cells transfected with
HSA-PEI-pcDNA-Apoptin, and not in the HSA-PEI-pcDNA-transfected or
blank control cells (Fig. 2D). These
results suggest that the HSA-PEI-pcDNA-Apoptin construct can be
transfected into breast cancer cells, resulting in the stable
expression of apoptin.
HSA-PEI-pcDNA-Apoptin reduces the
viability and induces the apoptosis of MCF-7 cells
To identify whether HSA-PEI-pcDNA-Apoptin was able
to reduce the viability of MCF-7 cells, MCF-7 cells were
transfected with HSA-PEI-pcDNA-Apoptin or HSA-PEI-pcDNA, or treated
with saline (blank control), and MTT assays were performed to
assess the cytotoxic effects. The cell viability of the
HSA-PEI-pcDNA-Apoptin group was decreased by 14.3% following
transfection with HSA-PEI-pcDNA-Apoptin, whereas the viability of
the other two groups did not change (Fig.
3A). MCF-7 cells were positively stained with Annexin V-PE and
7-ADD, and flow cytometric analysis showed that the apoptosis of
MCF-7 cells transfected with HSA-PEI-pcDNA-Apoptin was
significantly increased from 9.13±1.65 to 27.23±7.23% (P=0.013 vs.
control; Fig. 3B). These results
suggest that HSA-PEI-pcDNA-Apoptin has a significant effect on the
apoptosis and viability of breast cancer cells.
HSA-PEI-pcDNA-Apoptin effectively
inhibits tumor growth in MCF-7 xenografts
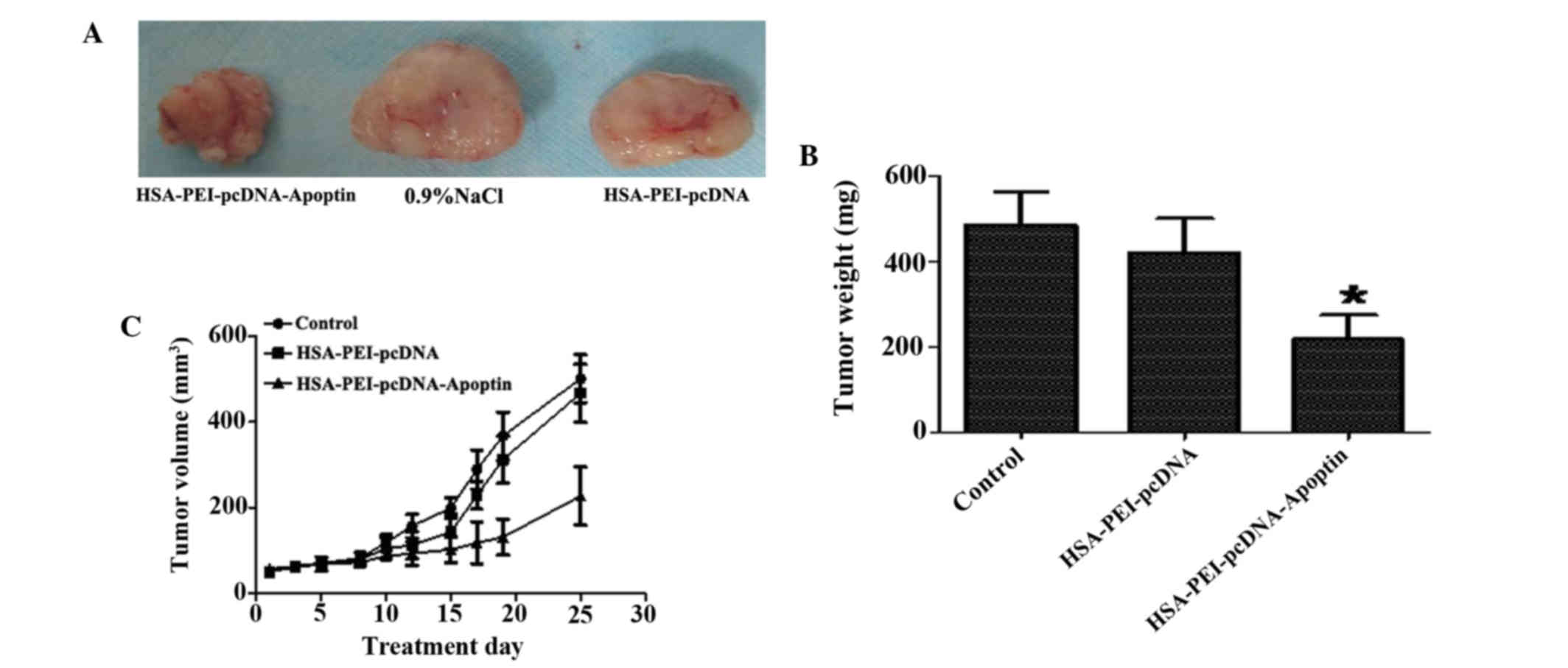
To evaluate the therapeutic effects of
HSA-PEI-pcDNA-Apoptin in vivo, HSA-PEI-pcDNA-Apoptin,
HSA-PEI-pcDNA or saline were injected into the tail veins of BALB/c
(nu-nu) athymic nude mice bearing MCF-7 cells once per day for 25
days, and the tumor volumes were measured every 3 days. As is shown
in Fig. 4, HSA-PEI-pcDNA-Apoptin
markedly inhibited tumor growth, including tumor weight and volume,
whereas there were no significant changes in the control group and
HSA-PEI-pcDNA group. The tumor weights following sacrifice were
significantly reduced in the mice injected with
HSA-PEI-pcDNA-Apoptin (P=0.0011 vs. HSA-PEI-pcDNA).
Expression of apoptin in breast tumor
and normal tissues
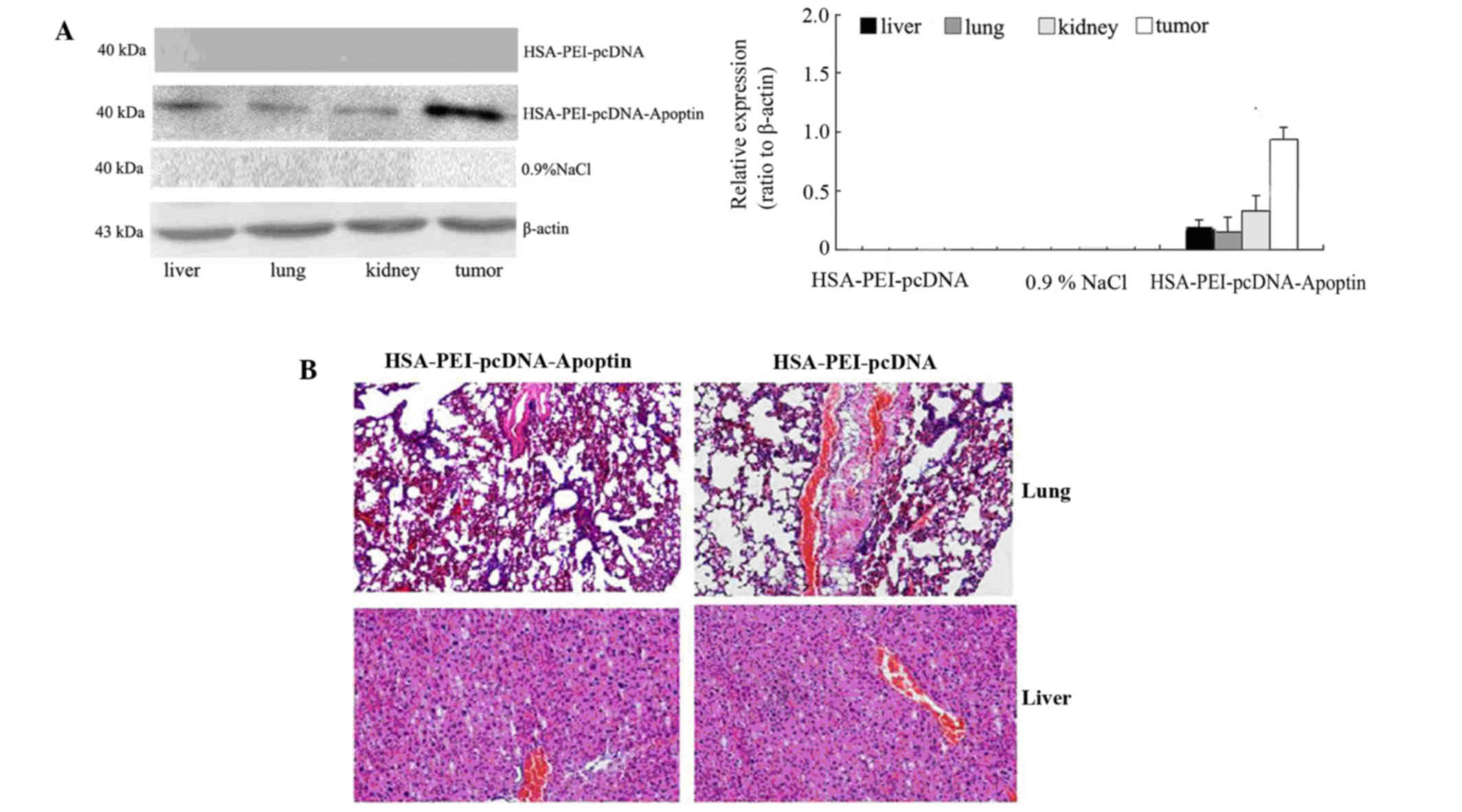
Western blotting was performed to detect whether
apoptin was expressed in the tumor tissue and normal tissues of
mice injected with HSA-PEI-pcDNA-Apoptin. As is shown in Fig. 5A, the mice injected with
HSA-PEI-pcDNA-Apoptin showed positive apoptin protein expression in
the normal tissues (liver, lungs and kidneys) and tumor tissues,
although the expression of apoptin was higher in the tumor tissue
compared with the normal tissues. Apoptin expression was rarely
detected in the mice injected with HSA-PEI-pcDNA or saline
(Fig. 5A). In addition, the
paraffin-embedded sections of the mice lung and liver tissues were
stained with HE, and the tissue morphology was observed under a
microscope. As is shown in Fig. 5B,
there were no pathological changes in the lung and liver tissues of
the mice injected with HSA-PEI-pcDNA-Apoptin or HSA-PEI-pcDNA,
suggesting that apoptin causes no damage to normal tissues.
Discussion
Efficacy and specificity is an important requirement
for successful cancer therapy (12).
The present study demonstrated the in vitro and in
vivo effects of an HSA-carrying apoptin construct on MCF-7
human breast cancer cells. Notably, this recombinant
HSA-PEI-pcDNA-Apoptin construct was able to selectively induce the
apoptosis of breast tumor cells.
Apoptin is a protein of 13.6 kDa that was derived
from the CAV and is encoded by the VP3 gene (13,14).
Apoptin has been shown to selectively induce the apoptosis of a
variety of tumor cells, including hepatomas, lymphomas,
cholangiocarcinomas, melanomas, and breast, lung, oral and colon
carcinomas, while not affecting the normal non-transformed human
cells, including primary fibroblasts, smooth muscle cells, T-cells,
hepatocytes, hematopoietic stem cells, keratinocytes and
endothelial cells (13,15–17). In
addition, numerous human tumor cell lines were shown to be
susceptible to apoptin (12,18–20), which
makes it a promising new tool for cancer gene therapy. However,
efficient systems are required to deliver apoptin to cancer cells
or to promote the expression of apoptin within these cells. The
apoptin gene is suitable for inclusion in technologies such as
conditionally replicative viruses or nonviral transduction methods
because of its potency and small size; however these novel delivery
strategies are yet to be validated (21). HSA may be a good choice for a vector
because of its characteristics: HSA is a major protein component of
blood plasma that has an important function in regulating colloidal
osmotic pressure and transporting numerous endogenous compounds,
including fatty acids, hormones, toxic metabolites and bile acids
(22). Furthermore, HSA selectively
binds to various drug molecules, and in doing so alters their
pharmacokinetic properties and biological activity (23,24). In a
previous study, an antitumor effect was reported in human breast
cancer cell lines transfected with a λ phage nanobioparticle
expressing the apoptin gene (12).
This platform allowed the present study to successfully construct
an apoptin expression vector that effectively transported apoptin
into breast cancer cells. The results presented in this study
provide suggested that HSA-mediated apoptin infection was able to
reduce the viability and induce the apoptosis of MCF-7 cells.
Furthermore, intravenous delivery of the HSA-PEI-pcDNA-Apoptin
construct decreased tumor growth in vivo, and resulted in no
damage to normal tissues.
At present, cancer therapy includes surgery,
chemotherapy, radiotherapy, targeted therapy and immunotherapy.
However, these conventional therapeutic methods have unavoidable
side effects, and resistance to cancer therapies has been reported
for the majority of tumors (25–32). Novel
therapeutic approaches that facilitate the selective targeting of
cancer cells have emerged in recent years. Furthermore, gene
therapy has become a research hot-spot. Gene therapy involving the
delivery of apoptin into cancer cells offers unique advantages over
current approaches for cancer therapy. Guan et al (18) developed a construct involving the
Salmonella typhimurium-mediated delivery of apoptin into
human laryngeal cancer cells, and demonstrated that the recombinant
was able to induce apoptosis in these cells in vitro and
in vivo, without causing significant side effects on normal
tissues. van der Eb et al (19) used HepG2 tumor-bearing mice to
investigate the in vivo effects of an adenovirus-expressing
apoptin, and demonstrated that treatment with apoptin significantly
improved the long-term survival of the mice. Other experiments also
reported that apoptin is a promising and useful gene for cancer
therapy (33–35).
In conclusion, the present study generated a
recombinant HSA-PEI-pcDNA construct expressing apoptin, and
investigated its antitumor effects in vitro and in
vivo. In vitro analyses demonstrated that the
HSA-PEI-pcDNA-Apoptin construct resulted in the successful
expression of apoptin in MCF-7 cells, and induced the apoptosis of
these cells. In vivo experiments showed that the injection
of HSA-PEI-pcDNA-Apoptin in nude mice bearing MCF-7 cells
significantly suppressed tumor growth, without causing damage to
normal tissues. These results indicated that HSA-PEI-pcDNA-Apoptin
may be considered a potential therapeutic agent for the treatment
of solid tumors, as apoptin was specifically active in malignant
cells and its toxicity in normal cells was negligible. However, the
mechanism of apoptin-induced cell death has yet to be fully
elucidated and should be investigated in further experiments. In
addition, further studies are required using breast tumor models to
determine the tumor-specific apoptotic effect of the recombinant
complex in its actual target environment, as well as its most
appropriate application in gene therapy.
Acknowledgements
The authors would like to thank Professor Chen Huang
at Xi'an Jiaotong University for providing the platform to conduct
the experiments and for the expert advice. This study was supported
by the Natural Science Foundation of Shaanxi Province, China (grant
no. S2013JC10894).
References
|
1
|
Xue H, Ni P, Lin B, Xu H and Huang G:
X-ray repair cross-complementing group 1 (XRCC1) genetic
polymorphisms and gastric cancer risk: A HuGE review and
meta-analysis. Am J Epidemiol. 173:363–375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
DeSantis C, Siegel R, Bandi P and Jemal A:
Breast cancer statistics, 2011. CA Cancer J Clin. 61:409–418. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cao W, Wang X and Li JC: Hereditary breast
cancer in the Han Chinese population. J Epidemiol. 23:75–84. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen WQ, Zeng HM, Zheng RS, Zhang SW and
He J: Cancer incidence and mortality in china, 2007. Chin J Cancer
Res. 24:1–8. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gehrig S, Sami H and Ogris M: Gene therapy
and imaging in preclinical and clinical oncology: Recent
developments in therapy and theranostics. Ther Deliv. 5:1275–1296.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Noteborn MH, Verschueren CA, Zantema A,
Koch G and van der Eb AJ: Identification of the promotor region of
chicken anemia virus (CAV) containing a novel enhancer-like
element. Gene. 150:313–318. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Danen-Van Oorschot AA, Fischer DF,
Grimbergen JM, Klein B, Zhuang S, Falkenburg JH, Backendorf C, Quax
PH, Van der Eb AJ and Noteborn MH: Apoptin induces apoptosis in
human transformed and malignant cells but not in normal cells. Proc
Natl Acad Sci USA. 94:5843–5847. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
O'Connor TP and Crystal RG: Genetic
medicines: Treatment strategies for hereditary disorders. Nat Rev
Genet. 7:261–276. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kobayashi K: Summary of recombinant human
serum albumin development. Biologicals. 34:55–59. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rhaese S, von Briesen H, Rübsamen-Waigmann
H, Kreuter J and Langer K: Human serum albumin-polyethylenimine
nanoparticles for gene delivery. J Control Release. 92:199–208.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shoae-Hassani A, Keyhanvar P, Seifalian
AM, Mortazavi-Tabatabaei SA, Ghaderi N, Issazadeh K, Amirmozafari N
and Verdi J: λ Phage nanobioparticle expressing apoptin efficiently
suppress human breast carcinoma tumor growth in vivo. PLoS One.
8:e799072013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Maddika S, Mendoza FJ, Hauff K, Zamzow CR,
Paranjothy T and Los M: Cancer-selective therapy of the future:
Apoptin and its mechanism of action. Cancer Biol Ther. 5:10–19.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Noteborn MH, Todd D, Verschueren CA, de
Gauw HW, Curran WL, Veldkamp S, Douglas AJ, McNulty MS, van der Eb
AJ and Koch G: A single chicken anemia virus protein induces
apoptosis. J Virol. 68:346–351. 1994.PubMed/NCBI
|
|
15
|
Tavassoli M, Guelen L, Luxon BA and Gäken
J: Apoptin: Specific killer of tumor cells? Apoptosis. 10:717–724.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Backendorf C, Visser AE, de Boer AG,
Zimmerman R, Visser M, Voskamp P, Zhang YH and Noteborn M: Apoptin:
Therapeutic potential of an early sensor of carcinogenic
transformation. Annu Rev Pharmacol Toxicol. 48:143–169. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Los M, Panigrahi S, Rashedi I, Mandal S,
Stetefeld J, Essmann F and Schulze-Osthoff K: Apoptin, a
tumor-selective killer. Biochim Biophys Acta. 1793:1335–1342. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guan GF, Zhao M, Liu LM, Jin CS, Sun K,
Zhang DJ, Yu DJ, Cao HW, Lu YQ and Wen LJ: Salmonella typhimurium
mediated delivery of apoptin in human laryngeal cancer. Int J Med
Sci. 10:1639–1648. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
van der Eb MM, Pietersen AM, Speetjens FM,
Kuppen PJ, van de Velde CJ, Noteborn MH and Hoeben RC: Gene therapy
with apoptin induces regression of xenografted human hepatomas.
Cancer Gene Ther. 9:53–61. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhuang SM, Shvarts A, van Ormondt H,
Jochemsen AG, van der Eb AJ and Noteborn MH: Apoptin, a protein
derived from chicken anemia virus, induces p53-independent
apoptosis in human osteosarcoma cells. Cancer Res. 55:486–489.
1995.PubMed/NCBI
|
|
21
|
Backendorf C, Visser AE, de Boer AG,
Zimmerman R, Visser M, Voskamp P, Zhang YH and Nobetorn M: Apoptin:
Therapeutic potential of an early sensor of carcinogenic
transformation. Annual review of pharmacology and toxicology.
48:143–69. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Peters T Jr: All About Albumin:
Biochemistry, Genetics, and Medical Applications. 1st. Academic
Press; San Diego, CA: 1996
|
|
23
|
Zhao XN, Liu Y, Niu LY and Zhao CP:
Spectroscopic studies on the interaction of bovine serum albumin
with surfactants and apigenin. Spectrochim Acta A Mol Biomol
Spectrosc. 94:357–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Abdi K, Nafisi SH, Manouchehri F, Bonsaii
M and Khalaj A: Interaction of 5-Fluorouracil and its derivatives
with bovine serum albumin. J Photochem Photobiol B. 107:20–26.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aleman BM, Moser EC, Nuver J, Suter TM,
Maraldo MV, Specht L, Vrieling C and Darby SC: Cardiovascular
disease after cancer therapy. EJC Suppl. 12:18–28. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Visovsky C: Treatment considerations for
the management of patients with hormone receptor-positive
metastatic breast cancer. J Adv Pract Oncol. 5:321–330.
2014.PubMed/NCBI
|
|
27
|
Wang MJ: Professor Hope S. Rugo: Reversing
resistance and hormone therapy for metastatic breast cancer. Chin
Clin Oncol. 3:512014.PubMed/NCBI
|
|
28
|
Jerusalem G, Rorive A and Collignon J: The
drug of the month: Everolimus (Afinitor) for the treatment of
metastatic breast cancer. Rev Med Liege. 69:510–517. 2014.(In
French). PubMed/NCBI
|
|
29
|
Bergen E, Berghoff AS, Rudas M, Dubsky P,
De Vries C, Sattlberger C, Mader RM, Zagouri F, Sparber C, Fitzal
F, et al: Taxanes plus trastuzumab compared to oral vinorelbine
plus trastuzumab in HER2-overexpressing metastatic breast cancer.
Breast Care (Basel). 9:344–348. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Domschke C and Schuetz F: Side effects of
bone-targeted therapies in advanced breast cancer. Breast Care
(Basel). 9:332–336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Greenlee H, Balneaves LG, Carlson LE,
Cohen M, Deng G, Hershman D, Mumber M, Perlmutter J, Seely D, Sen
A, et al: Society for Integrative Oncology: Clinical practice
guidelines on the use of integrative therapies as supportive care
in patients treated for breast cancer. J Natl Cancer Inst Monogr.
2014:346–358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yoo C, Kim SB, Ahn JH, Kim JE, Jung KH,
Gong GY, Son BH, Ahn SH, Ahn SD, Kim HH, et al: A randomized phase
II trial of capecitabine plus vinorelbine followed by docetaxel
versus Adriamycin plus cyclophosphamide followed by docetaxel as
neoadjuvant chemotherapy for breast cancer. Cancer Res Treat.
47:406–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
An S, Nam K, Choi S, Bai CZ, Lee Y and
Park JS: Nonviral gene therapy in vivo with PAM-RG4/apoptin as a
potential brain tumor therapeutic. Int J Nanomedicine. 8:821–834.
2013.PubMed/NCBI
|
|
34
|
Liu L, Wu W, Zhu G, Liu L, Guan G, Li X,
Jin N and Chi B: Therapeutic efficacy of an hTERT promoter-driven
oncolytic adenovirus that expresses apoptin in gastric carcinoma.
Int J Mol Med. 30:747–754. 2012.PubMed/NCBI
|
|
35
|
Pan Y, Fang L, Fan H, Luo R, Zhao Q, Chen
H and Xiao S: Antitumor effects of a recombinant pseudotype
baculovirus expressing Apoptin in vitro and in vivo. Int J Cancer.
126:2741–2751. 2010.PubMed/NCBI
|