Introduction
Pelvic organ prolapse (POP) is a major health
concern for women in the reproductive and menopausal years. POP
involves the decent of pelvic organ and the pelvic floor, caused by
insufficiency of the fibrous connective tissue and striated muscles
that form the pelvic floor (1).
Fibroblasts and their product collagen are the main components
within the connective tissue. The change in the number of
fibroblasts in pelvic floor connective tissue may cause changes in
the collagen content (2,3), and the functionality of fibroblasts
may also be related to a deficiency of collagen, which has been
observed in patients with POP (4). The type, amount and degree of
cross-linking of collagen contribute to the tension of the
connective tissue. Collagen I specifically contributes to the
tensile force of the connective tissue forming the pelvic floor,
and is degraded by a family of enzymes called the matrix
metalloproteinases (MMPs), whose expression levels are modulated by
locally produced tissue inhibitors of metalloproteinases (TIMPs)
(5). General speaking, changes in
both the qualitative and quantitative properties of collagen have
been linked to patients with POP (6,7).
In 1996, Jackson et al (7)
demonstrated that genitourinary prolapse is associated with a
reduction in total collagen content supporting the findings of
another study (8). Kerkhof et
al found that pyridinoline collagen cross-links which reflect
the level of mature collagen in the prolapse site increased
significantly, compared to the non-prolapse group (9). Vulic et al found there was
increased expression of MMP-1 and decreased expression of collagen
I in uterosacral ligaments of women with POP compared with non-POP
women (10). Dviri et al
concluded that the expression of MMP-1 and MMP-9 appears to be
increased in tissues from women with POP (11). Wang et al demonstrated that
TIMP-1 expression levels in a POP patient group were significantly
lower than those in the control group (12). Thus, it is hypothesized that
changes in the metabolism of collagen I are regulated by MMP-1 and
TIMP-1, and other matrix metalloproteinases and its tissue
inhibitors, are related to the physiopathology of POP.
Moreover, it has been confirmed that the metabolism
of collagen can be impacted by advanced glycation end products
(AGEs) (13). AGEs, the products
of nonenzymatic glycation and oxidation of proteins and lipids,
accumulate in diverse biological settings including: diabetes,
inflammation, renal failure and aging. AGEs adjust the metabolism
of target proteins through the receptor of advanced glycation end
products (RAGE) (14), and
activate an array of signal transduction cascades, such as MAPK,
ROS, p38, NO and nuclear factor-κB (NF-κB). Together these pathways
are involved in numerous biological functions including, but not
limited to: skin aging, cardiovascular injury and remodeling,
diabetes, inflammation and gingival hyperplasia (15,16). In the context of skin aging, AGEs
promote fibroblast apoptosis, inhibit the synthesis of collagen,
and accelerate the degradation of collagen through the balance of
MMP and TIMP (17), which may be
similar to the metabolic change in collagen in connective tissue of
the pelvic floor in POP.
Concerning the actual role of AGEs in the
pathological physiology of POP, Jackson et al also found
that both intermediate intermolecular cross-links and advanced
glycation cross-links were increased in prolapsed tissue (7). Moreover, our previous study
indicated that collagen I levels were decreased in prolapse tissue
while the expression of AGEs in prolapse tissue was concomitantly
increased. RAGE expression, however, was found to remain stable in
pelvic tissue of prolapsed patients (18). Thus, we speculated that AGEs
impact the metabolism of collagen in the pelvis through RAGE on the
surface of fibroblasts and downstream pathways; however, the
related mechanism remains to be elucidated, and there is no
information concerning the role of AGEs and its receptor in POP. In
the present study, we describe the metabolism of collagen I
activated by AGEs through MMP-1, TIMP, and changes in p38 and NF-κB
following AGE-RAGE interactions.
Materials and methods
The present study was approved by the Ethics
Committee of the Obstetrics and Gynecology Hospital of Fudan
University, Shanghai, China. This study included two parts: i) the
impact of AGEs on the metabolism of collagen I in human vaginal
fibroblasts (HVFs) obtained from patients with POP. Six primary
cultured HVF samples from 3 cases of POP (51, 71 and 65 years of
age, respectively), and 3 cases of non-POP (55, 57 and 70 years of
age, respectively), were collected. The protein expression of
collagen I, MMP-1, TIMP-1 and RAGE were chosen for study; ii) the
mechanism involved in the impact of AGEs on the metabolism of
collagen I in primary cultured HVFs; the molecules, RAGE, p38 MAPK
and NF-κB were selected for study.
Reagents
Anti-collagen I (sc-136154), anti-AGE (ab23722)
antibody, anti-RAGE monoclonal antibody (sc-365154) and
anti-vimentin monoclonal antibody were purchased from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA). AGE protein (ab51995),
anti-MMP-1 (ab119922) and anti-TIMP-1 (ab28261) and were purchased
from Abcam (Cambridge, MA, USA). Anti-RAGE siRNA were purchased
from GenePharma Co., Ltd. (Shanghai, China). SB203580 (inhibitor of
p38 MAPK) was purchased from Selleck Chemicals (Houston, TX, USA)
and PDTC (inhibitor of NF-κB) was purchased from Beyotime
Biotechnology (Shanghai, China). Transfection reagent
Lipofectamine® 2000 was purchased from Invitrogen™ Life
Technologies (Carlsbad, CA, USA). Anti-p-p38 MAPK antibody (#9211),
anti-p38 MAPK antibody (#9212), anti p-p65 NF-κB antibody (#3033)
and anti p65 NF-κB antibody (#3034), were all purchased from Cell
Signaling Technology (Danvers, MA, USA).
Culture and identification of the primary
fibroblasts
Human fibroblasts derived from the vaginal wall were
obtained from patients suffering from POP or other diseases who
required hysterectomy at the Obstetrics and Gynecology Hospital of
Fudan University. All subjects provided informed consent which was
signed accompanied by an Operation Consent Form prior to surgery.
Briefly, fresh vaginal wall tissue specimens from the surgical
margin of the free womb were washed in phosphate-buffered saline
(PBS) (containing 1% penicillin, streptomycin, amphotericcin B) at
4°C for 5 min for 3 times, and digested at 37°C for 30 min with PBS
containing 2% collagenase. Following isolation, the cells were
cultured in Dulbecco's modified Eagle's medium (DMEM) (containing
10% fetal bovine serum, 1% penicillin, streptomycin, amphotericcin
B) in 5% CO2 at 37.5°C, with replacement of the mediun
every 2–3 days. HVFs were identified by anti-vimentin antibody
staining and subsequently stored in liquid nitrogen for further
study (19,20).
Cell counting assay
HVFs were thawed and allowed to recover for 72 h,
and the cell counting per dish was affirmed by an automatic cell
counting apparatus (Bio-Rad TC10TM; Bio-Rad Laboratories, Hercules,
CA, USA) after pre-stage test. Then the cells were fixed using 10%
trichloroacetic acid at 60 min at 4°C. After discarding the
supernatant, the plates were washed with deionized water 5 times
and dried at room temperature. Fifty microliters of sulforhodamine
B solution (0.4% sulforhodamine B dissolved in 0.1% acetic acid)
was added to the cells followed by 30 min of incubation. Unbound
sulforhodamine B was washed with 1% acetic acid, and the plates
were air-dried. One-hundred fifty microliters of 10 mM Tris-base
(pH 10.5) was added to each well, and the plates were shaken gently
for 20 min on a plate shaker. The absorbance of each well was
determined using a microplate reader (Tecan, Männedorf,
Switzerland) at 560 nm (21,22).
Western blot analysis
Collagen I, RAGE, MMP-1, TIMP-1, p38, p-p38, p65 and
p-p65 were detected in the vaginal tissues (50 mg) which were cut
into small fragments and were ground by hand in a glass homogenizer
on ice. Ten microliters of phenylmethanesulfonyl fluoride was added
followed by 1000 μl RIPA lysis buffer [50 mM Tris-HCl, 150
mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 2 mM NaF, 2 mM
ethylenediaminetetraacetic acid (EDTA), 0.1% sodium dodecyl sulfate
(SDS) and a protease inhibitor cocktail tablet] a few minutes
later, and the proteins were separated by SDS-polyacrylamide gel
and transferred to PVDF membranes by electrophoresis. Membranes
were blocked overnight at 4°C in protein blocker (Bio-Rad
Laboratories). The membranes were exposed to the primary antibody
at a dilution of 1:500 for 12 h at 4°C. After three washes with
PBST, the membranes were hybridized with HRP-conjugated anti-mouse
or ant-rabbit IgG antibody. Protein bands were visualized by
enhanced chemiluminescence (ECL; Amersham Pharmacia, Piscataway,
NJ, USA), and relative intensities of the protein bands were
analyzed using ImageJ software [National Institutes of Health
(NIH), Bethesda, MD, USA].
Measurement of mRNA expression by
quantitative (real-time) PCR (qPCR)
Total RNA was isolated from human vaginal wall
tissues using the Total RNA Extraction Miniprep System (Promega,
Madison, WI, USA) according to the manufacturer's instructions. The
PCR were established according to the instructions provided by
Applied Biosystems (Foster City, CA, USA). In brief, first-strand
cDNA was synthesized using 1 μg of total RNA and the
First-Strand cDNA synthesis kit (Invitrogen). For qPCR, 12
μl of cDNA solution was mixed with 0.5 μmol/l
primers, 5 mmol/l magnesium chloride and 2 μl of Master
SYBR-Green in nuclease-free water with a final volume of 20
μl. The primers used for PCR were collagen I, forward,
5′-GTGCGATGACGTGATCTG TGA-3′ and reverse,
5′-CGGTGGTTTCTTGGTCGGT-3′; MMP-1, forward,
5′-GGGGCTTTGATGTACCCTAGC-3′ and reverse, 5′-TGT
CACACGCTTTTGGGGTTT-3′; TIMP-1, forward, 5′-CTT CTGCAATTCCGACCTCGT-3
′ and reverse, 5′-ACGCTGGTATAAGGTGGTCTG-3′; RAGE, forward,
5′-GTGTCCTTCCCAACGGCTC-3′ and reverse, 5′-ATTG CCTGGCACCGGAAAA-3′.
PCR amplification was performed using the following cycling
conditions: 95°C for 30 sec, then 40 cycles at 95°C for 5 sec
followed by elongation at 60°C for 20 sec. Glyceraldehyde
3-phosphate dehydrogenase (GAPDH) was used as an endogenous control
against which the different template values were normalized. All
PCR reactions were performed in duplicate. The threshold cycle (Ct)
method was used for quantification. Relative quantification of the
genes was performed by using the ΔΔCt approach.
Silencing of AGE, RAGE, MAPK and NF-κB
cell signaling pathways using siRNA interference and
inhibitors
HVFs were cultured in DMEM for 2 days post-recovery
from deep freeze. The cells were treated with a RAGE-targeting
siRNA expression system, which included four sequences siRNA-1,
5-GAGUAUCUGUGAAGGAACAtt-3; siRNA-2, 5-UGU UCCUUCACAGAUACUCtt-3;
siRNA-3, 5-AUCUAC AAUUUCUGGCUUCtt-3; and control, 5-GUUCUCCG
AACGUGUCACGUtt-3. These siRNAs were chemically synthesized,
purified and annealed by GenePharma Biotechnology (Shanghai,
China), which were designed to target the coding sequence of RAGE,
as previously described (23–25). Both the control and vector
containing siRNAs were transfected into fibroblasts using
Lipofectamine 2000. The efficiency of siRNA delivery was determined
by western blotting as previously described (26). SB203580 was used to inhibit MAPK
at 10 μM (27), while 10
μM PDTC was used to inhibit NF-κB (28).
Statistical analysis
Data are expressed as means ± SE. One-way ANOVA was
used to determine significant differences between the POP group and
non-POP group; p<0.05 was assigned as criterion for significance
for western blotting and PCR assays, using SPSS 16.0.
Results
Part 1 results
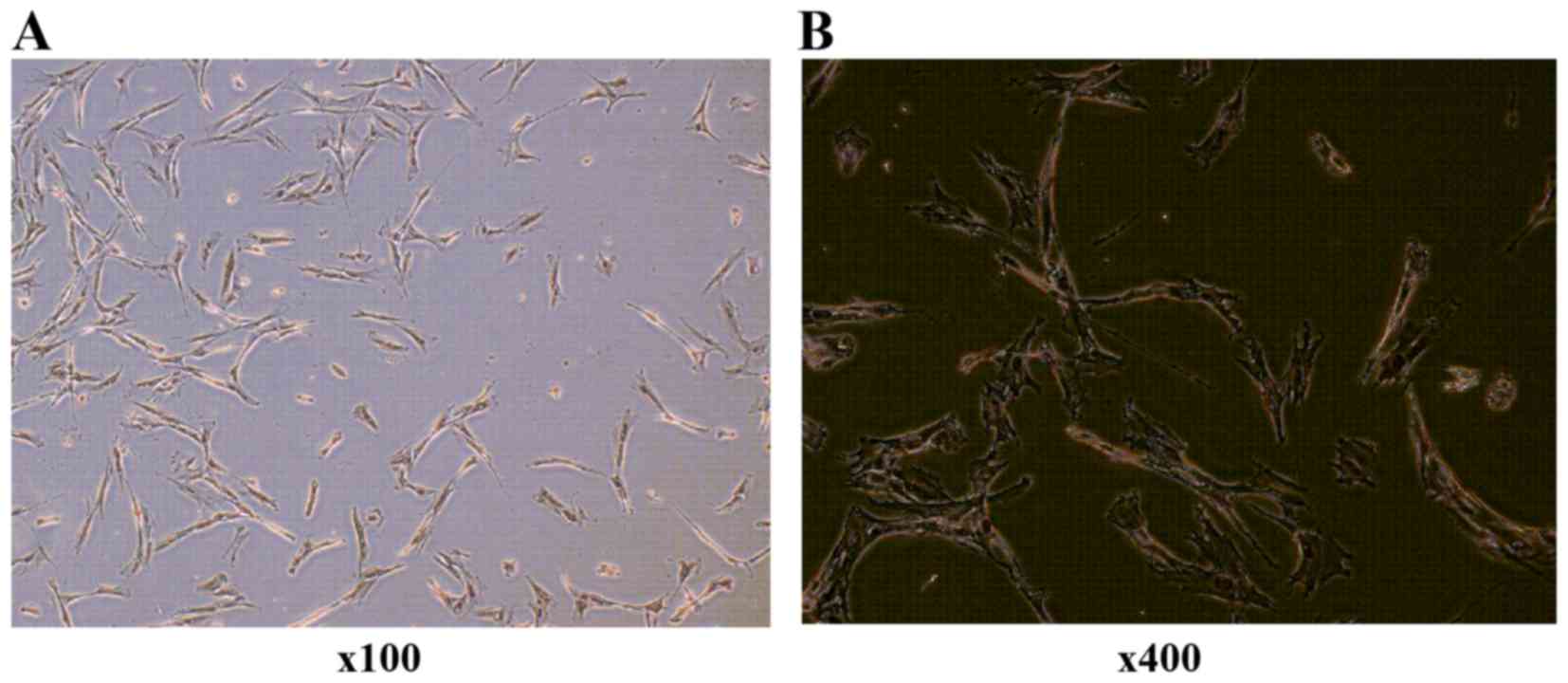
Identification of HVF cultures
Six primary cultured HVF samples from 3 cases of POP
(51, 71 and 65 years of age, respectively), and 3 cases of non-POP
(55, 57 and 70 years of age, respectively), were collected. Primary
cultured fibroblasts were identified by anti-vimentin antibody
(Fig. 1).
Impact of AGEs on HVFs
The cell counting assay demonstrated that the
proliferation of the primary cultured fibroblasts was inhibited by
AGE protein at various concentrations (0, 25, 50, 75, 100 and 150
mg/ml; 48 h) (Fig. 2A and B) and
incubation times (0, 12, 24, 48, 72 and 96 h; AGEs, 50 mg/l)
(Fig. 2C and D). Proliferation of
HVFs in the POP groups (25 mg/l) was more readily inhibited than
that noted in the non-POP groups (25 mg/l). At the same various
treatment time (0, 12, 24, 48, 72 and 96 h; AGEs, 50 mg/l), the
effect of AGEs on the proliferation of HVFs was analogous between
the POP and non-POP groups.
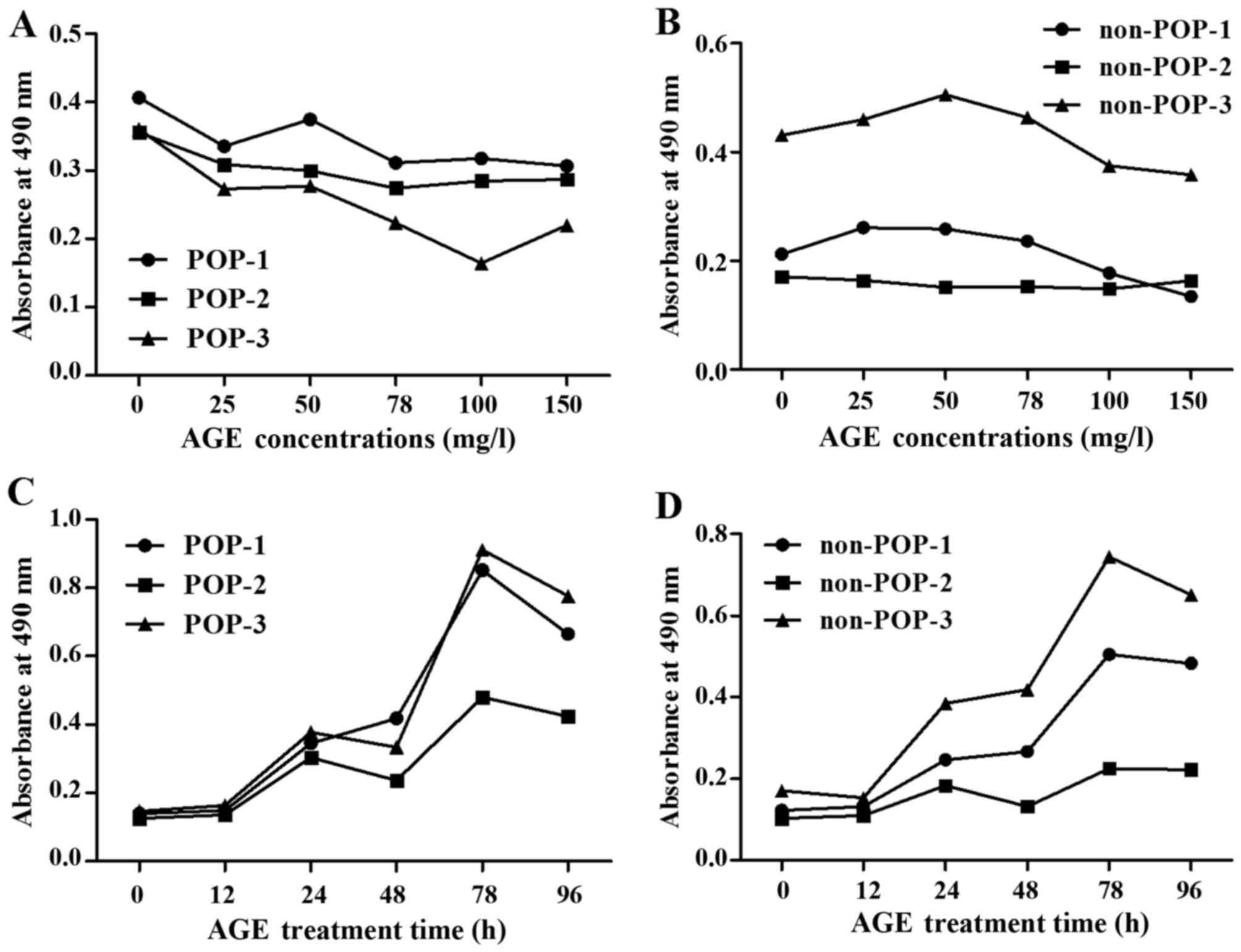 | Figure 2Comparison of the effects of advanced
glycation end products (AGEs) on the proliferation of human vaginal
fibroblasts (HVFs). The effects were detected by sulforhodamine B.
(A and B) Proliferation curves of HVFs of pelvic organ prolapse
(POP) and the control group cultured under different concentrations
of AGEs (at doses of 0, 25, 50, 75, 100 and 150 mg/l, 48 h). (C and
D) Proliferation curves of HVFs of POP and the control group
cultured for various time periods (treatment time, 0, 12, 24, 48,
72 and 96 h, at 50 mg/l). 'POP-1, 2, 3' belong to the POP groups'
'non-POP-1, 2, 3' belong to the control groups. Proliferation of
the HVFs of the POP groups was more easily inhibited than that of
the control groups. |
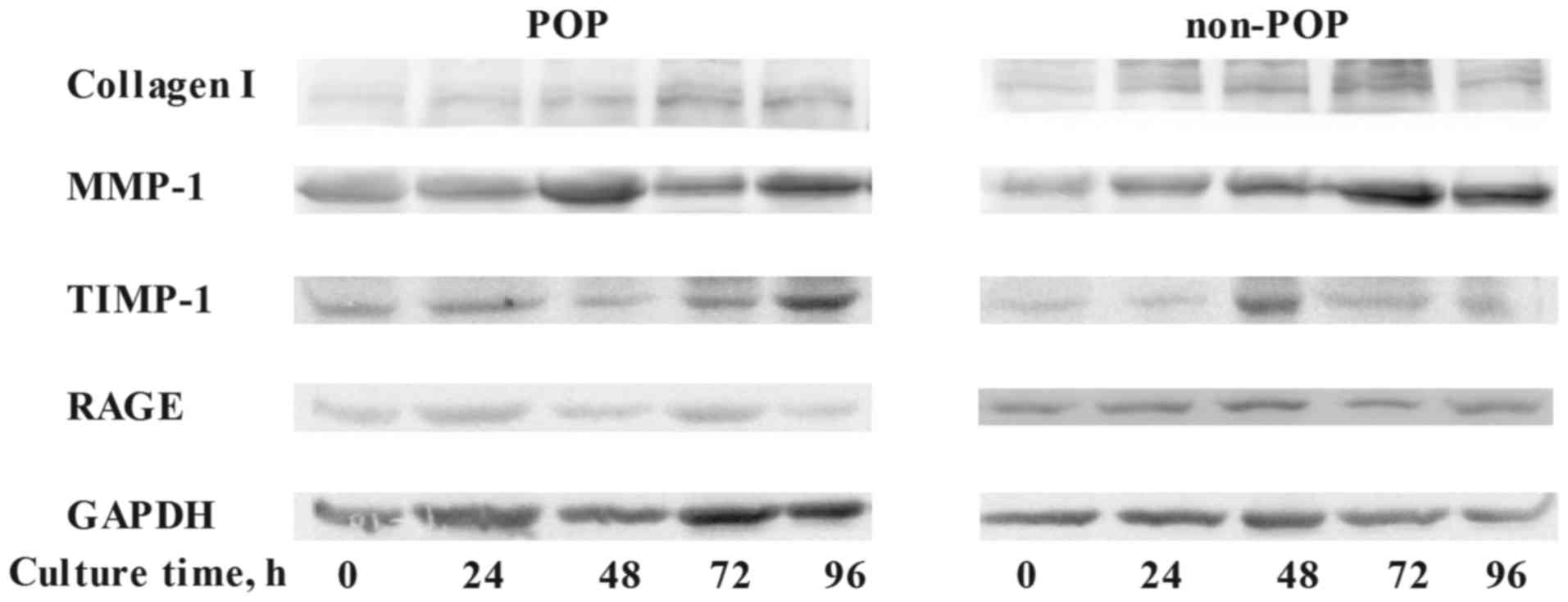
Effect of AGEs on the metabolism of
collagen I in HVFs by western blotting
To assess the effect of AGEs on the metabolism of
collagen I in HVFs, the expression levels of collagen I, MMP-1,
TIMP-1 and RAGE were evaluated by western blotting (Fig. 3). Following treatment with various
concentrations of AGEs (0, 25, 50, 75, 100 and 150 mg/ml; 48 h),
the expression of collagen I in the POP group was decreased
(inversely correlating with the concentration of AGEs) as compared
to control levels (p<0.05) (Fig.
3A). In contrast to these findings, expression of MMP-1 in the
POP group was increased relative to the control group (p<0.05),
with increasing concentrations of AGEs (Fig. 3B). The expression of TIMP-1 in POP
was lower than that in the non-POP group, and no significant
changes occurred in both groups with increasing concentrations of
AGEs (Fig. 3C). No significant
change in the expression of RAGE was observed among the groups
(Fig. 3D). Across various
time-points (0, 24, 48, 72 and 96 h; AGEs, 50 mg/l), there was no
significant difference in the expression tendency of collagen I,
MMP-1, TIMP-1 and RAGE between the POP and non-POP groups
(P>0.05) (Fig. 4).
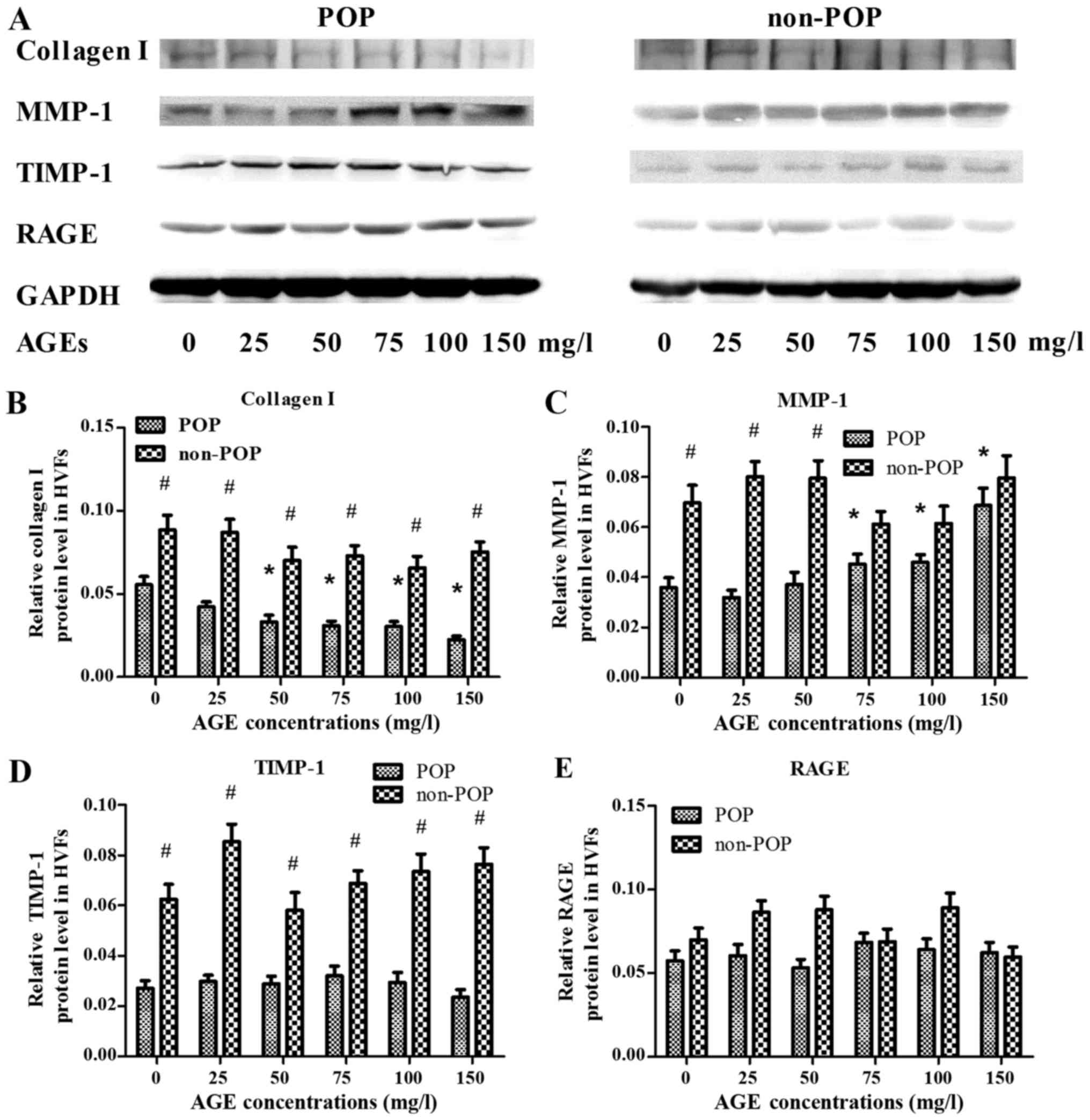 | Figure 3Treatment with advanced glycation end
products (AGEs) was shown to affect the expression levels of
collagen I, matrix metalloproteinases-1 (MMP-1), tissue inhibitor
of metalloproteinase-1 (TIMP-1) and receptor of advanced glycation
end products (RAGE) in human vaginal fibroblasts (HVFs) by western
blot analysis. (A) Representative western blots; (B–E) Protein
levels of collagen I, MMP-1, TIMP-1 and RAGE, respectively. HVFs of
pelvic organ prolapse (POP) and control groups were cultured under
different concentrations of AGEs (at the dose of 0, 25, 50, 75, 100
and 150 mg/l, respectively, 48 h). Data are presented as the means
± SEM. *p<0.05 vs. control (0 mg/l).
#P<0.05 as compared with POP group. |
Effect of AGEs on the metabolism of
collagen I in HVFs by qPCR
To examine the effect of AGEs on the metabolism of
collagen, HVFs were harvested at various time-points (0, 1, 2, 4,
16, 24 and 48 h). There was a peak in RNA transcription (including
collagen I, MMP-1, TIMP-1 and RAGE) detected at 4 h after treatment
with the AGEs (50 mg/l) in the POP group, but also at 16 h in the
control group (Fig. 5). Our
results demonstrated that HVFs in the POP group were more sensitive
to AGEs than the control group. We compared the difference in
metabolism of collagen I under various AGE concentrations between
two groups at 4 and 16 h of treatment, respectively. At 4 h, the
mRNA expression of collagen I and TIMP-1 in the POP group was
significantly decreased as compared to the mRNA expression in the
control group (blank, AGEs, 0 mg/l) with the increasing
concentration of AGEs (p<0.05) (Fig. 6A and C). However, the expression
of TIMP-1 decreased more significantly in the POP than in the
non-POP group. MMP-1 mRNA expression increased up to a
concentration of 50 mg/l AGEs in the POP group, and then decreased
(Fig. 6B). No significant changes
in RAGE mRNA expression were observed between the 2 groups
(Fig. 6D). At 16 h, the level of
collagen I in both the POP and non-POP groups increased firstly and
then decreased, respectively with the increasing concentrations of
AGEs; the change tendency in the level of TIMP-1 and RAGE in the 2
groups, respectively was analogous to those changes observed at 4
h. Similarly, the mRNA expression of MMP-1 in the POP group was
also similar to those results observed at 4 h (Fig. 7).
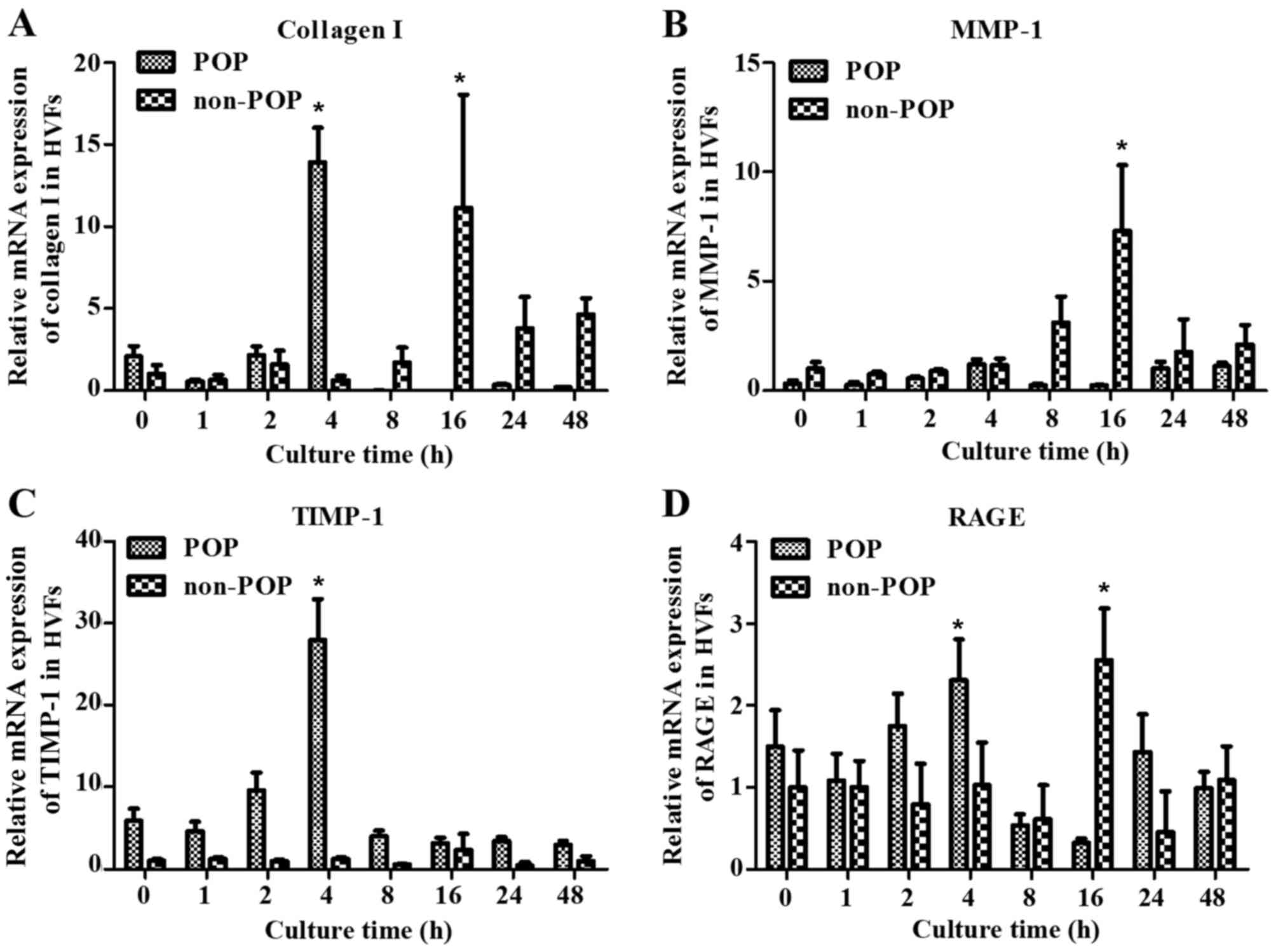 | Figure 5Treatment with advanced glycation end
products (AGEs) was shown to impact the mRNA expression of (A)
collagen I, (B) matrix metalloproteinase-1 (MMP-1), (C) tissue
inhibitor of metalloproteinase-1 (TIMP-1) and (D) receptor of
advanced glycation end products (RAGE) by real-time PCR in human
vaginal fibroblasts (HVFs). mRNA expression of collagen I, MMP-1,
TIMP-1 and RAGE cultured under AGEs (50 mg/l) for different culture
times in HVFs of pelvic organ prolapse (POP) and non-POP groups.
The culture times were 0, 1, 2, 4, 8, 16, 24 and 48 h,
respectively. Peak mRNA levels occurred at 4 h after AGE treatment
in the POP groups, and 16 h in the control groups respectively.
Data are presented as the means ± SEM (n=3), *p<0.06
compared to the control (blank, 0 mg/l) in their own group,
respectively. |
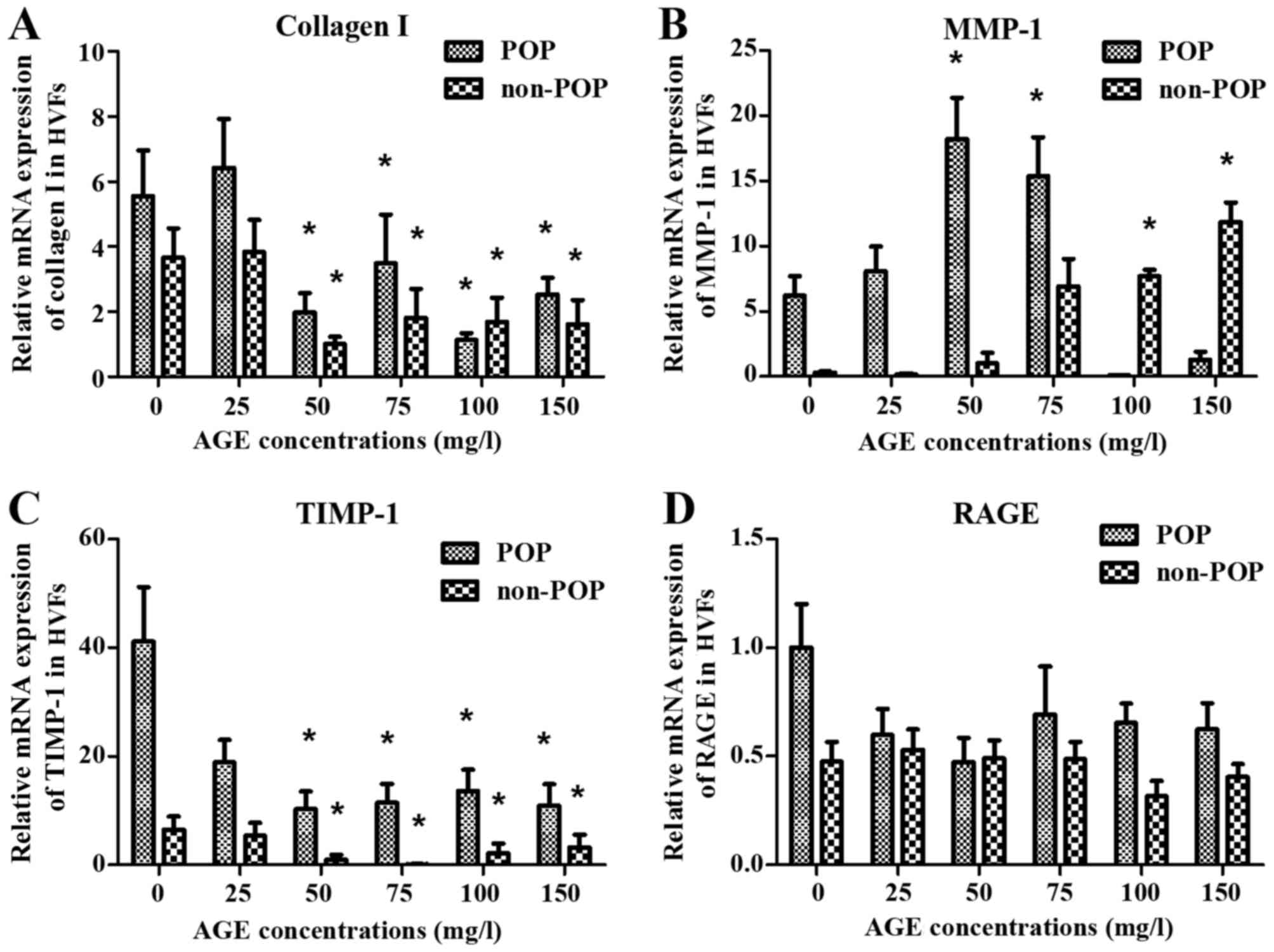 | Figure 6Effects on the mRNA expression of (A)
collagen I, (B) matrix metalloproteinase-1 (MMP-1), (C) tissue
inhibitor of metalloproteinase-1 (TIMP-1) and (D) receptor of
advanced glycation end products (RAGE) at 4 h after treatment with
advanced glycation end products (AGEs). mRNA expression of collagen
I, MMP-1, TIMP-1 and RAGE was detected by real-time PCR after human
vaginal fibroblasts (HVFs) of pelvic organ prolapse (POP) and
control group were cultured with AGEs at different concentrations
(0, 25, 50, 75, 100 and 150 mg/l, respectively). (A-D) Collagen I,
MMP-1, TIMP and RAGE groups, respectively. Data are presented as
the mean ± SEM (n=3), *p<0.05 compared to the control
(0 mg/l) in their own group, respectively. |
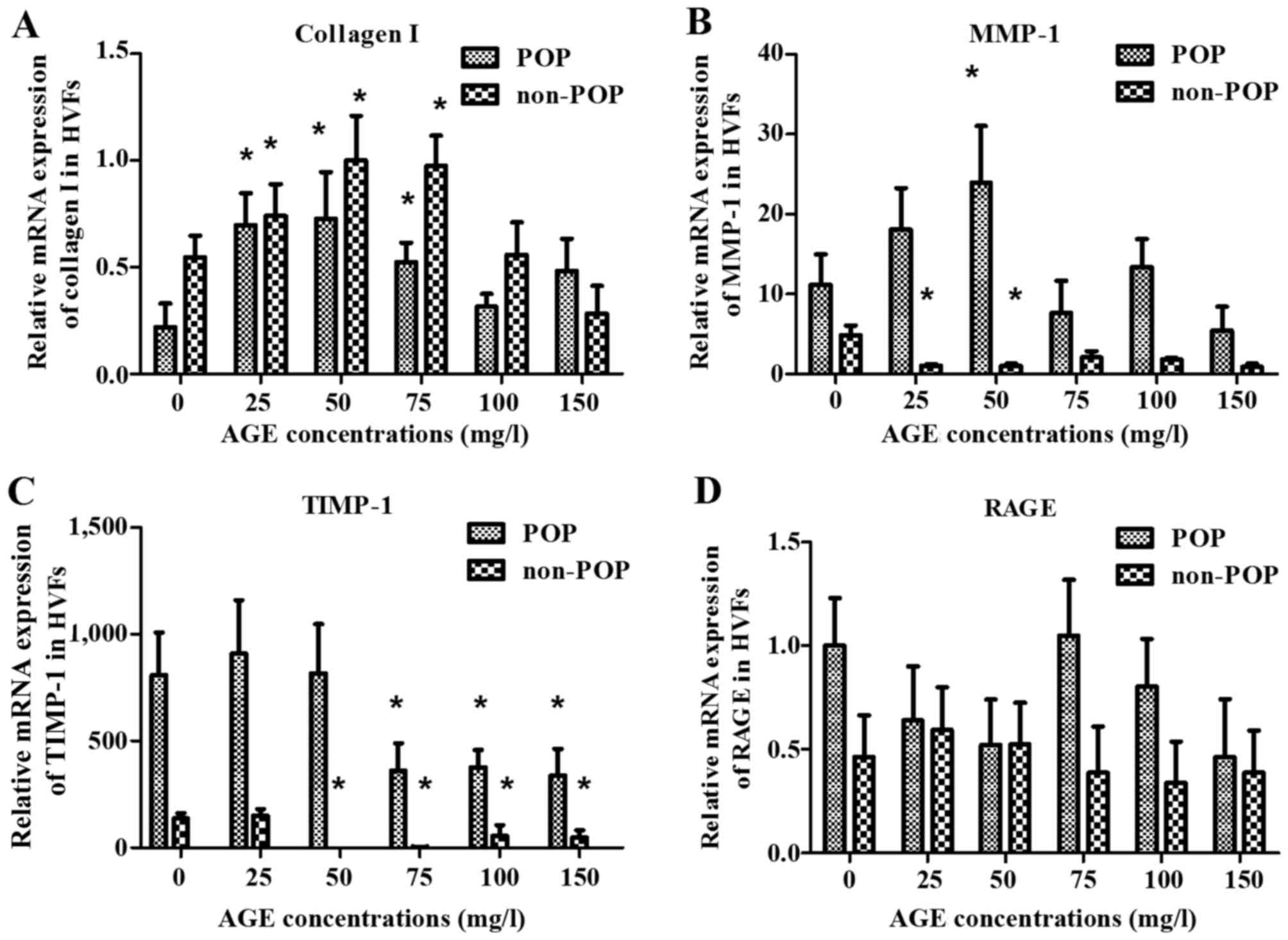 | Figure 7Effect on the mRNA expression of (A)
collagen I, (B) matrix metalloproteinase-1 (MMP-1), (C) tissue
inhibitor of metalloproteinase-1 (TIMP-1) and (D) receptor of
advanced glycation end products (RAGE) at 16 h after treatment with
advanced glycation end products (AGEs) is shown. mRNA expression of
collagen I, MMP-1, TIMP-1 and RAGE was detected by real-time PCR
after human vaginal fibroblasts (HVFs) of pelvic organ prolapse
(POP) and control group were cultured under AGEs at different
concentrations (0, 25, 50, 75, 100 and 150 mg/l, respectively).
(A-D) Collagen I, MMP-1, TIMP and RAGE, respectively. Data are
presented as the mean ± SEM (n=3), *p<0.06 compared
to the control (0 mg/l, respectively). |
Part 2 results
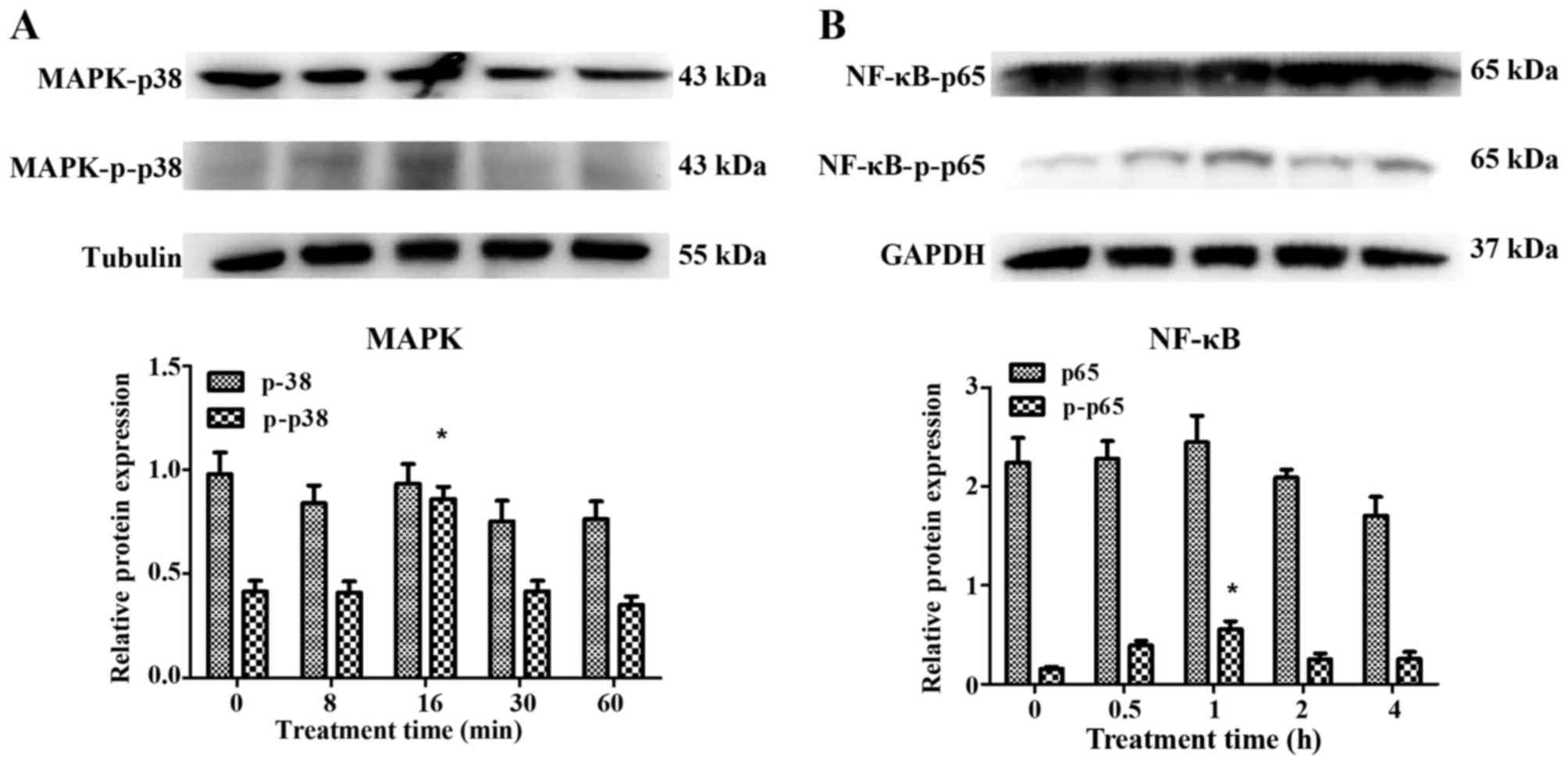
Identification and inhibition of
signaling molecules of p38 MAPK, NF-κB-p65 and RAGE
Following treatment with AGEs (50 mg/l), no
significant changes were observed in the expression of p38 MAPK and
NF-κB-p65, as detected by western blot analysis in the HVFs.
Notably, phosphorylation products increased in the fibroblasts from
patients with POP. p-p38 increased to maximum levels at 16 min
post-treatment and decreased soon afterwards, while the levels of
p-p65 peaked at 60 min (Fig. 8).
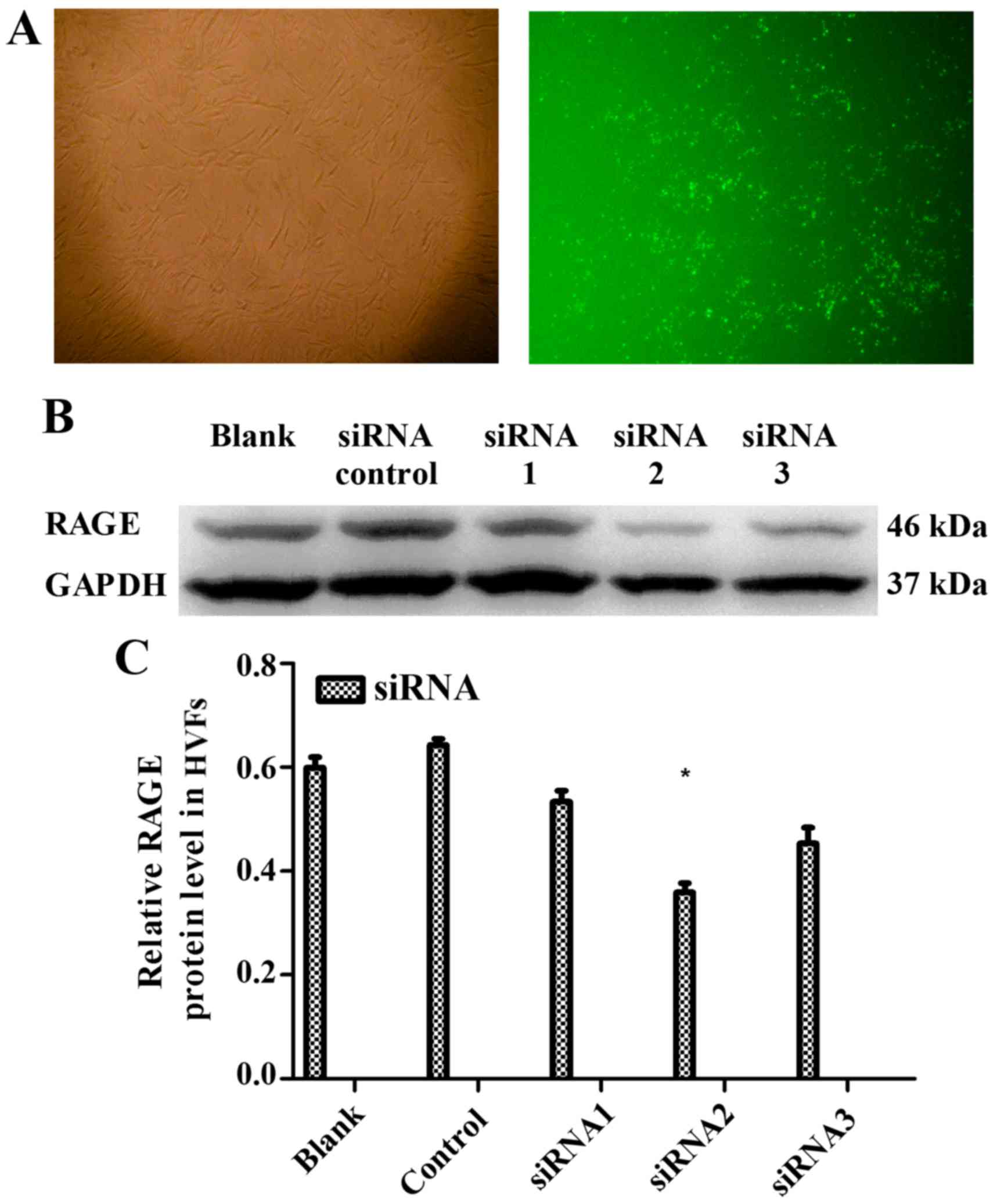
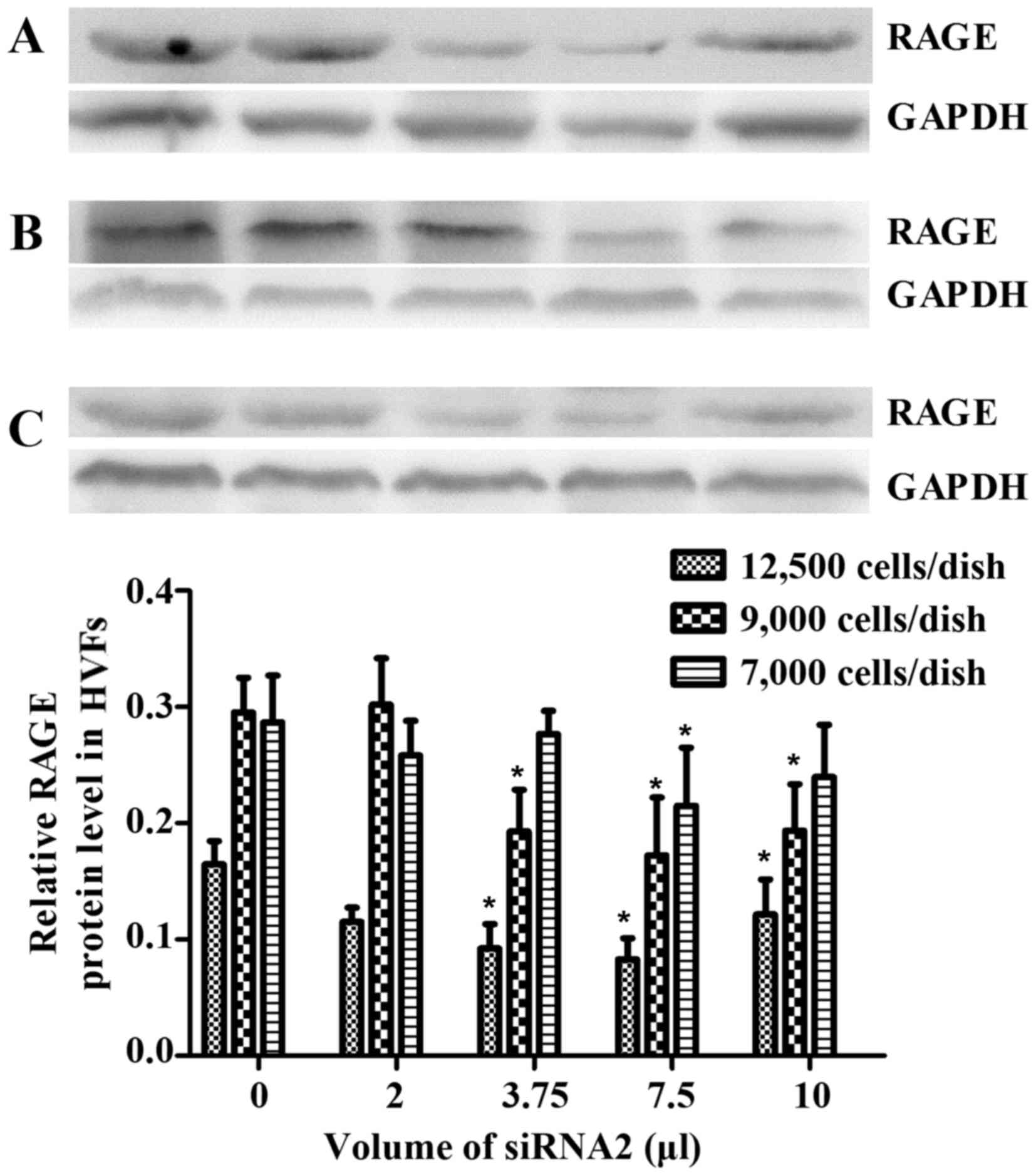
We selected 3 siRNAs (1, 2 and 3) to silence RAGE. The blocking
efficiency of siRNA2 was higher than that of the other siRNAs, and
was subsequently used for all downstream applications (Fig. 9). The fibroblasts were cultured at
12,500, 9,000 and 7,000 cells/dish using a 6-well plate with 2 ml
DMEM, and a series of volumes of Lipofectamine 2000 and siRNA2; the
blocking efficiency of siRNA2 for RAGE was evident (p<0.05) when
using a 6-well plate with 2 ml DMEM, 7.5 μl Lipofectamine
2000 and 7.5 μl siRNA2 (Fig.
10). Due to cell death by Lipofectamine 2000, which can affect
sequence treatment, 9,000 cells/dish were employed
appropriately.
Investigation of the RAGE, MAPK and NF-κB
signaling pathways in the HVFs by western blot analysis
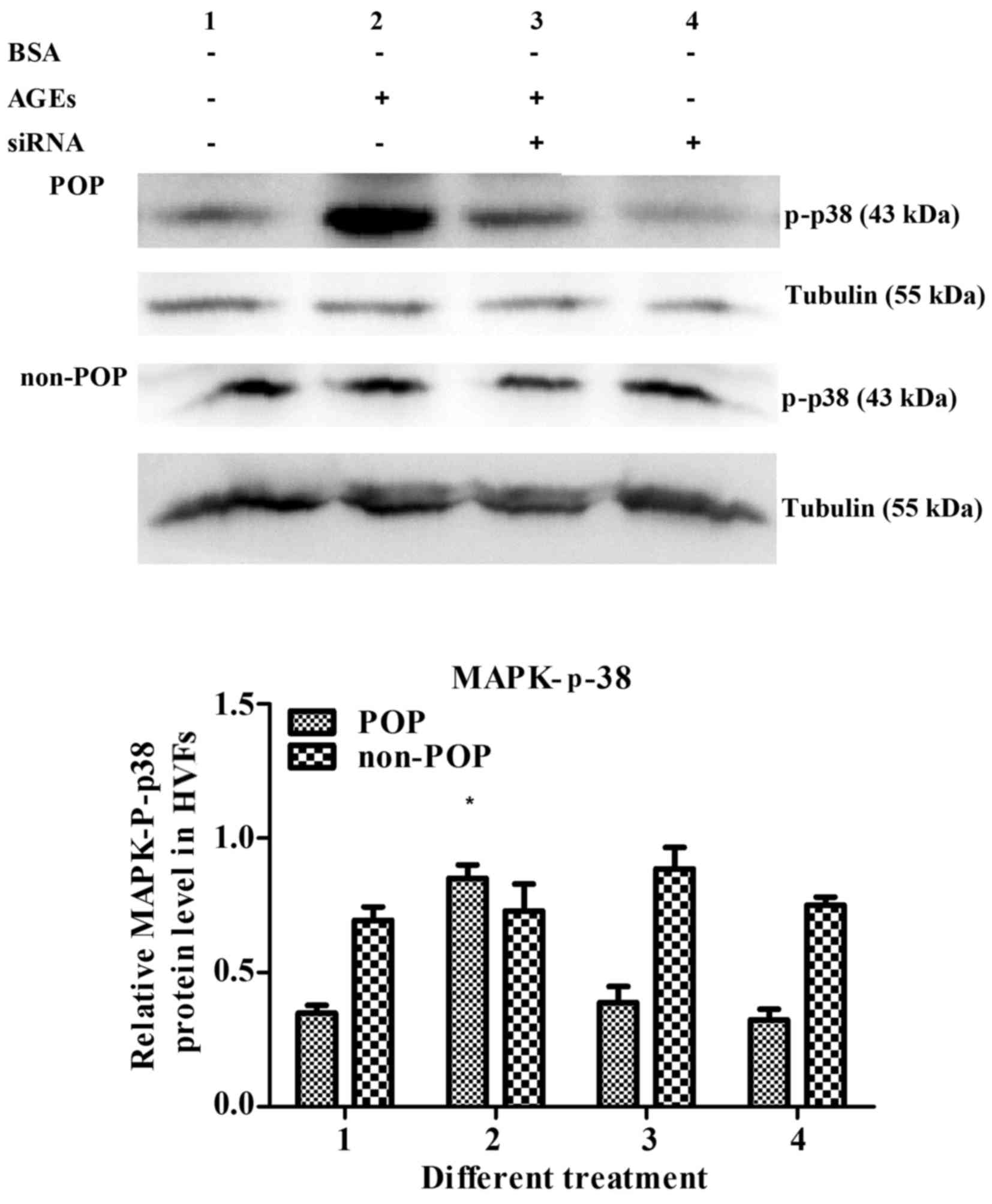
Detection of p38 MAPK
In the HVFs from patients with POP, the levels of
-p-p38 MAPK increased following AGE treatment (50 mg/l); however,
this effect was reversed following the inhibition of RAGE by siRNA.
These results suggest that while AGEs can activate p-p38 MAPK, this
process is RAGE-dependent. In HVFs from non-POP patients, the
levels of p-p38 MAPK were not significantly affected by AGE
treatment, which highlights a POP-specific p-p38 MAPK and AGE
connection (Fig. 11).
Detection of NF-κB-p-p65
In the HVFs from the POP and non-POP groups alike,
the levels of NF-κB-p-p65 were increased following treatment with
AGEs, and these decreased following siRNA-mediated RAGE-inhibition.
These results suggest that AGE-RAGE interactions affect target
proteins through NF-κB-p-p65. Notably, the p-p65 levels were not
affected by the inhibition of p-p38, suggesting that NF-κB
activation is not a result of a linear activation of AGE/RAGE/MAPK
signaling (Fig. 12).
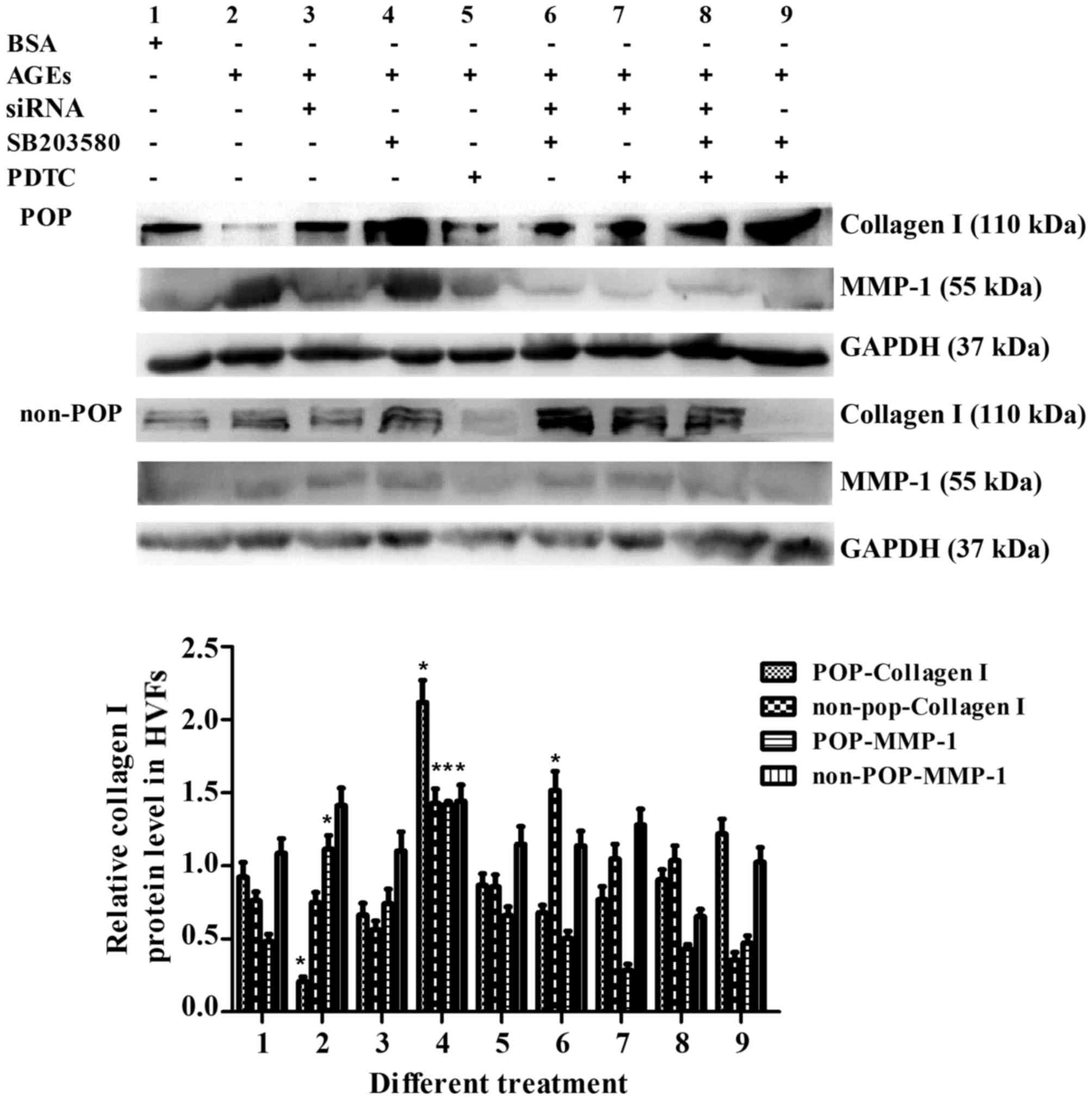
Detection of collagen I and MMP-1
Collagen I and MMP-1 were detected when RAGE, MAPK
and NF-κB were blocked in various combinations. In the HVFs from
patients with POP, the protein expression of collagen I decreased
following AGE treatment (50 mg/l) (Fig. 13), but increased to varying
degrees after the molecular or pharmacological inhibition of RAGE,
MAPK or NF-κB. In the HVFs from the non-POP group, there were no
significant changes observed following the same treatment modality.
For MMP-1 metabolism, the opposite trend as compared to collagen I
was observed (Fig. 13).
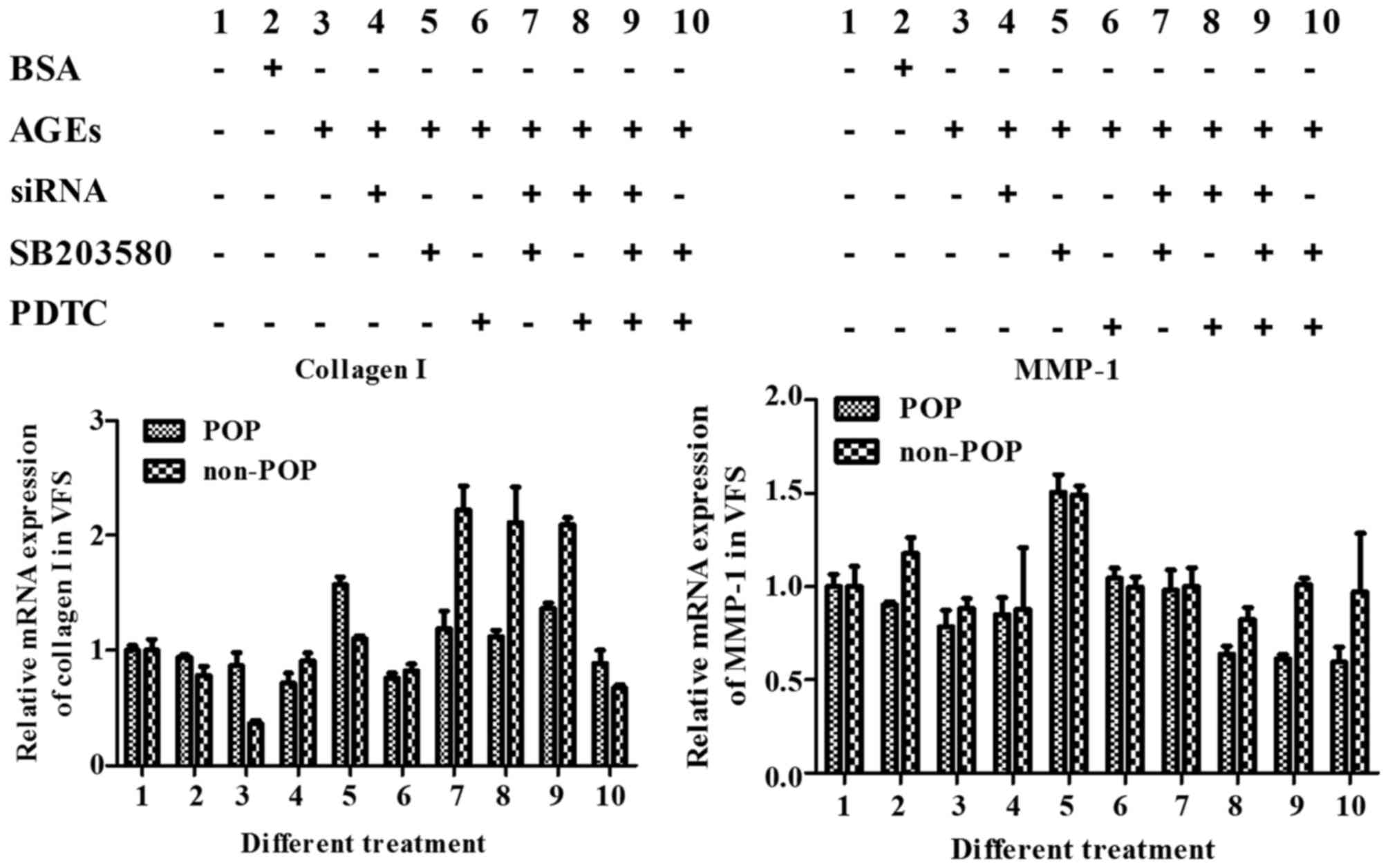
Ivestigation of the RAGE, MAPK and
NF-κB signaling pathways in HVFs by qPCR
To identify changes in the RAGE/MAPK/NF-κB pathway
when it was affected by AGEs, the mRNA levels of target collagen I
and MMP-1 were evaluated. In both the treated and control groups,
collagen I mRNA levels decreased after the signaling pathways were
activated by AGEs (50 mg/l) and increased following pathway (RAGE,
MAPK and NF-κB) inhibition. The effect was more pronounced using
combinations of inhibitors rather than a single inhibitor. For
MMP-1, the mRNA expression was increased following treatment with
AGEs, and decreased following treatment with the inhibitors
(Fig. 14).
Discussion
HVFs play an important role in the pathophysiology
of pelvic organ prolapse (POP), which controls the integrity of
collagen, and thereby impacts the mechanical properties of the
pelvic floor (8). Primary culture
of HVFs is commonly used to evaluate the connective tissue of POP.
In the present study, fibroblasts were successfully cultured from
vaginal tissue, and then identified by anti-vimentin antibody.
Considering vaginal fungi, amphotericin B was added to the DMEM
culture media. This method was simple and highly efficient and the
cells remained stable following recovery from long-term freeze.
Previous research has described the impacts of AGEs
on fibroblast proliferation. Research has reported that AGEs
promote the proliferation of fibroblasts (29), but others demonstrated that AGEs
induced the apoptosis of fibroblasts or inhibited proliferation
(30). In the present study, the
cell counting of HVFs treated by AGEs was detected by
sulforhodamine B. With increasing concentrations of AGEs,
fibroblast proliferation from the POP patient group was
significantly inhibited, suggesting that fibroblasts in POP were
more likely to be inhibited. These results explain why the number
of fibroblasts in the pelvic floor of POP is reduced (31).
In skin aging and gingival hyperplasia, AGEs can
regulate the metabolism of collagen by the AGE-RAGE pathway. In
this manner, synthesis of collagen I can be inhibited, degradation
can be promoted, and apoptosis of fibroblasts can be induced
(32,33). It still remains unclear how AGEs
impact collagen metabolism in the pelvis of POP. In this study,
secretion of collagen I, MMP-1 and TIMP-1 was increased gradually
in both control and treated groups as time increased in accordance
with a previously study (18).
Notably, the content of collagen I was decreased
more gradually with increasing concentrations of AGEs in the
patient group than that in the control group, which suggests that
new collagen I in POP is inhibited weakening pelvic connective
tissue repair. In keeping with these findings, other groups have
shown that MMP-1 expression, which can accelerate collagen I
degradation, is increased in pelvic tissue from POP (12,18). In this study, MMP-1 expression was
more pronounced in the POP group than that in the control group. We
also found that TIMP-1 levels were unaffected in both groups. This
suggested that TIMP-1 could not exert a protective effect in the
face of elevated MMP-1 expression. While the expression and
structure of RAGE have been shown to be an important contributor to
many diseases, including diabetic nephropathies (34), no changes were observed in POP
vaginal tissues (18). This
result suggests that AGEs regulate the metabolism of collagen
through RAGE-binding. These results were further confirmed by qPCR
data. Firstly, the levels of mRNA of collagen I, MMP-1 and TIMP-1
under various incubation times were tested, and peak mRNA levels
were observed at 4 h in the POP group, but 16 h in the control
group. This suggests that POP HVFs are more sensitive to AGEs than
non-POP cells. Secondly, the level of mRNA under various
concentration gradients was tested at 4 and 16 h, respectively.
Irrespective of the time-point, the change in mRNA in the POP group
was more readily detected than in the control. It was interesting
to note that the change in TIMP-1 mRNA was similar (in trend) to
that of collagen I, gradually decreasing in the face of increasing
levels of AGEs. Notably, in spite of these trends, TIMP-1 protein
expression remained stable suggesting a possible mechanism
dependent on post-translational modifications. Similarly, RAGE mRNA
and protein levels were unaffected suggesting that AGEs function
through an AGEs/RAGE-dependent pathway.
Characterization of AGE/RAGE pathway activation and
downstream signaling was an important objective in this study.
Previous studies have suggested that AGE/RAGE signaling involves
numerous signaling pathways including: eNOS, NAD (P) H-ROS,
p21RAS-MAPK, p38 MAPK, Cdc42-Rac and NF-κB. We chose to
specifically evaluate p38 MAPK and NF-κB-p65 signaling pathways
(35,39). Phosphorylation targets MAPK p39
and NF-κB-p65 increased substantially following treatment with
AGEs, but the non-phosphorylated products remained stable,
indicating that phosphorylated products participate in the
metabolism of collagen I.
To characterize downstream cell signaling pathways,
we inhibited specific cell signaling molecules alone or in
combination. We inhibited RAGE using an siRNA-based strategy,
validating our results using western blotting. Of the three siRNAs
evaluated, siRNA2 was most effective. Furthermore, we optimized the
transfection conditions. Specifically we addressed a number of
concerns including the number of cells and the amount of
transfection reagent both of which greatly impact the transfection
efficiency and cell viability, respectively. In contrast to this
siRNA-based approach we pharmacologically inhibited MAPK (SB203580)
and NF-κB (PDTC) and validating the effectiveness of this approach
by western blotting. In the POP group but not the control cells,
p-p38 was activated by AGEs but this effect was reversed by siRNA.
p-p65 could not be activated after RAGE was inhibited, but was
activated after p-p38 was inhibited in the two groups. These
results clarify that AGEs can activate NF-κB through RAGE and
downstream cell molecules besides p-p38 in POP-derived cells. We
also found that both collagen I and MMP-1 expression could be
modulated by activation and inhibition of these pathways. When
treatment with AGEs was utilized, the expression of collagen I was
inhibited, while MMP-1 was activated; when RAGE was silenced by
siRNA and p-p38 was blocked by SB203580, the suppression of
collagen I was recovered, but MMP-1 was inhibited; and the control
group remained stable.
In the last part of this study, the mRNA synthesis
of target proteins of MMP-1 and collagen I was detected after three
signaling molecules were blocked in various combinations. General
speaking, the mRNA expression trends matched those observations
made at the protein level by western blotting. On the one hand, the
result of PCR highlighted the impact of AGEs on the AGE, RAGE, MAPK
and NF-κB signaling pathways which are involved in the synthesis
and degradation metabolism of collagen, and may explain the reason
why collagen I was decreased in the vaginal tissue of POP. On the
other hand, there are multiple factors which can affect the
complicated process from binding of receptor to protein secretion,
and therefore innumerable additional factors may have
confounded/impacted the interpretation of these results.
In conclusion, AGEs can affect the metabolism of
collagen through RAGE, but not directly through changes in
expression or structure. AGEs activate p-p38 MAPK and NF-κB-p-p65
pathways, thereby regulating collagen metabolism, although other
pathways may also participate. Taken together, our study provides
enhanced understanding of the mechanism through which AGEs
contribute to collagen metabolism in pelvic tissue of POP and the
pathophysiology of POP.
Acknowledgments
This study was supported by grants from the Science
and Technology Commission of Shanghai Municipality (no.
124119a500); the General Program of National Natural Science
Foundation of China (no. 81671439).
References
|
1
|
Chow D and Rodríguez LV: Epidemiology and
prevalence of pelvic organ prolapse. Curr Opin Urol. 23:293–298.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sun B, Zhou L, Wen Y, Wang C, Baer TM,
Pera RR and Chen B: Proliferative behavior of vaginal fibroblasts
from women with pelvic organ prolapse. Eur J Obstet Gynecol Reprod
Biol. 183:1–4. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ruiz-Zapata AM, Kerkhof MH,
Zandieh-Doulabi B, Brölmann HA, Smit TH and Helder MN: Functional
characteristics of vaginal fibroblastic cells from premenopausal
women with pelvic organ prolapse. Mol Hum Reprod. 20:1135–1143.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen HY, Lu Y, Qi Y, Bai WP and Liao QP:
Relationship between the expressions of mitofusin-2 and procollagen
in uterosacral ligament fibroblasts of postmenopausal patients with
pelvic organ prolapse. Eur J Obstet Gynecol Reprod Biol.
174:141–145. 2014. View Article : Google Scholar
|
|
5
|
Kanta J: Collagen matrix as a tool in
studying fibroblastic cell behavior. Cell Adh Migr. 9:308–316.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lim VF, Khoo JK, Wong V and Moore KH:
Recent studies of genetic dysfunction in pelvic organ prolapse: the
role of collagen defects. Aust N Z J Obstet Gynaecol. 54:198–205.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jackson SR, Avery NC, Tarlton JF, Eckford
SD, Abrams P and Bailey AJ: Changes in metabolism of collagen in
genitourinary prolapsed. Lancet. 347:1658–1661. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Han L, Wang L, Wang Q, Li H and Zang H:
Association between pelvic organ prolapsed and stress urinary
incontinence with collagen. Exp Ther Med. 7:1337–1341. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kerkhof MH, Ruiz-Zapata AM, Bril H,
Bleeker MC, Belien JA, Stoop R and Helder MN: Changes in tissue
composition of the vaginal wall of premenopausal women with
prolapse. Am J Obstet Gynecol. 210:168.e1–9. 2014. View Article : Google Scholar
|
|
10
|
Vulic M, Strinic T, Tomic S, Capkun V,
Jakus IA and Ivica S: Difference in expression of collagen type I
and matrix metalloproteinase-1 in uterosacral ligaments of women
with and without pelvic organ prolapse. Eur J Obstet Gynecol Reprod
Biol. 155:225–228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dviri M, Leron E, Dreiher J, Mazor M and
Shaco-Levy R: Increased matrix metalloproteinases-1,-9 in the
uterosacral ligaments and vaginal tissue from women with pelvic
organ prolapse. Eur J Obstet Gynecol Reprod Biol. 156:113–117.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang X, Li Y, Chen J, Guo X, Guan H and Li
C: Differential expression profiling of matrix metalloproteinases
and tissue inhibitors of metalloproteinases in females with or
without pelvic organ prolapse. Mol Med Rep. 10:2004–2008. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Willett TL, Pasquale J and Grynpas MD:
Collagen modifications in postmenopausal osteoporosis: Advanced
glycation endproducts may affect bone volume, structure and
quality. Curr Osteoporos Rep. 12:329–337. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ajith TA and Vinodkumar P: Advanced
glycation end products: Association with the pathogenesis of
diseases and the current therapeutic advances. Curr Clin Pharmacol.
11:118–127. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yamagishi S, Fukami K and Matsui T:
Evaluation of tissue accumulation levels of advanced glycation end
products by skin autofluorescence: A novel marker of vascular
complications in high-risk patients for cardiovascular disease. Int
J Cardiol. 185:263–268. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu H, Jiang H, Ren H, Hu X, Wang X and Han
C: AGEs and chronic subclinical inflammation in diabetes: Disorders
of immune system. Diabetes Metab Res Rev. 31:127–137. 2015.
View Article : Google Scholar
|
|
17
|
Gkogkolou P and Böhm M: Advanced glycation
end products: Key players in skin aging? Dermatoendocrinol.
4:259–270. 2012. View Article : Google Scholar
|
|
18
|
Chen Y, Huang J, Hu C and Hua K:
Relationship of advanced glycation end products and their receptor
to pelvic organ prolapse. Int J Clin Exp Pathol. 8:2288–2299.
2015.PubMed/NCBI
|
|
19
|
El Mourabit H, Loeuillard E, Lemoinne S,
Cadoret A and Housset C: Culture model of rat portal
myofibroblasts. Front Physiol. 7:1202016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Olszewski S, Olszewska E, Popko J,
Poskrobko E, Sierakowski S and Zwierz K: Fibroblast-like synovial
cells in rheumatoid arthritis - the impact of infliximab on
hexosaminidase activity. Adv Clin Exp Med. 24:807–813. 2015.
View Article : Google Scholar
|
|
21
|
Yong WK, Ho YF and Malek SN: Xanthohumol
induces apoptosis and S phase cell cycle arrest in A549 non-small
cell lung cancer cells. Pharmacogn Mag. 11(Suppl 2): S275–S283.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nájera-Martínez M, García-Latorre EA and
Reyes-Maldonado E: Halomethane-induced cytotoxicity and cell
proliferation in human lung MRC-5 fibroblasts and NL20-TA
epithelial cells. Inhal Toxicol. 24:762–773. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Radia AM, Yaser AM, Ma X, Zhang J, Yang C,
Dong Q, Rong P, Ye B, Liu S and Wang W: Specific siRNA targeting
receptor for advanced glycation end products (RAGE) decreases
proliferation in human breast cancer cell lines. Int J Mol Sci.
14:7959–7978. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Monden M, Koyama H, Otsuka Y, Morioka T,
Mori K, Shoji T, Mima Y, Motoyama K, Fukumoto S, Shioi A, et al:
Receptor for advanced glycation end products regulates adipocyte
hypertrophy and insulin sensitivity in mice: Involvement of
Toll-like receptor 2. Diabetes. 62:478–489. 2013. View Article : Google Scholar :
|
|
25
|
Volz HC, Laohachewin D, Seidel C,
Lasitschka F, Keilbach K, Wienbrandt AR, Andrassy J, Bierhaus A,
Kaya Z, Katus HA, et al: S100A8/A9 aggravates post-ischemic heart
failure through activation of RAGE-dependent NF-κB signaling. Basic
Res Cardiol. 107:2502012. View Article : Google Scholar
|
|
26
|
Angiolilli C, Kabala PA, Grabiec AM, Van
Baarsen IM, Ferguson BS, García S, Malvar Fernandez B, McKinsey TA,
Tak PP, Fossati G, et al: Histone deacetylase 3 regulates the
inflammatory gene expression programme of rheumatoid arthritis
fibroblast-like synoviocytes. Ann Rheum Dis. 76:277–285. 2017.
View Article : Google Scholar :
|
|
27
|
Liu H, Liu A, Shi C and Li B: Curcumin
suppresses transforming growth factor-β1-induced cardiac fibroblast
differentiation via inhibition of Smad-2 and p38 MAPK signaling
pathways. Exp Ther Med. 11:998–1004. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Su C, Chen M, Huang H and Lin J:
Testosterone enhances lipopolysaccharide-induced interleukin-6 and
macrophage chemotactic protein-1 expression by activating the
extracellular signal-regulated kinase 1/2/nuclear factor-κB
signalling pathways in 3T3-L1 adipocytes. Mol Med Rep. 12:696–704.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Marelli B, Le Nihouannen D, Hacking SA,
Tran S, Li J, Murshed M, Doillon CJ, Ghezzi CE, Zhang YL, Nazhat SN
and Barralet JE: Newly identified interfibrillar collagen
crosslinking suppresses cell proliferation and remodelling.
Biomaterials. 54:126–135. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li W, Xu Q, Deng Y, Yang Z, Xing S, Zhao
X, Zhu P, Wang X, He Z and Gao Y: High-mobility group box 1
accelerates lipopolysaccharide-induced lung fibroblast
proliferation in vitro: Involvement of the NF-κB signaling pathway.
Lab Invest. 95:635–647. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kökçü A, Yanik F, Cetinkaya M, Alper T,
Kandemir B and Malatyalioglu E: Histopathological evaluation of the
connective tissue of the vaginal fascia and the uterine ligaments
inwomen with and without pelvic relaxation. Arch Gynecol Obstet.
266:75–78. 2002. View Article : Google Scholar
|
|
32
|
Niu Y, Xie T, Ge K, Lin Y and Lu S:
Effects of extracellular matrix glycosylation on proliferation and
apoptosis of human dermal fibroblasts via the receptor for advanced
glycosylated end products. Am J Dermatopathol. 30:344–351. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Abbass MM, Korany NS, Salama AH, Dmytryk
JJ and Safiejko-Mroczka B: The relationship between receptor for
advanced glycation end products expression and the severity of
periodontal disease in the gingiva of diabetic and non diabetic
periodontitis patients. Arch Oral Biol. 57:1342–1354. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Matsui T, Nakashima S, Nishino Y, Ojima A,
Nakamura N, Arima K, Fukami K, Okuda S and Yamagishi S: Dipeptidyl
peptidase-4 deficiency protects against experimental diabetic
nephropathy partly by blocking the advanced glycation end
products-receptor axis. Lab Invest. 95:525–533. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Giridharan VV, Thandavarayan RA, Arumugam
S, Mizuno M, Nawa H, Suzuki K, Ko KM, Krishnamurthy P, Watanabe K
and Konishi T: Schisandrin B ameliorates ICV-infused amyloid β
induced oxidative stress and neuronal dysfunction through
inhibiting RAGE/NF-κB/MAPK and up-regulating HSP/Beclin expression.
PLoS One. 10:e01424832015. View Article : Google Scholar
|
|
36
|
Wu Q, Li S, Li X, Sui Y, Yang Y, Dong L,
Xie B and Sun Z: Inhibition of advanced glycation endproduct
formation by lotus seedpod oligomeric procyanidins through
RAGE-MAPK signaling and NF-κB activation in high-fat-diet rats. J
Agric Food Chem. 63:6989–6998. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Medeiros MC, Frasnelli SC, Bastos AS,
Orrico SR and Rossa C Jr: Modulation of cell proliferation,
survival and gene expression by RAGE and TLR signaling in cells of
the innate and adaptive immune response: Role of p38 MAPK and
NF-κB. J Appl Oral Sci. 22:185–193. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
He ZW, Qin YH, Wang ZW, Chen Y, Shen Q and
Dai SM: HMGB1 acts in synergy with lipopolysaccharide in activating
rheumatoid synovial fibroblasts via p38 MAPK and NF-κB signaling
pathways. Mediators Inflamm. 2013:5967162013. View Article : Google Scholar
|
|
39
|
de Bittencourt Pasquali MA, Gelain DP,
Zeidán-Chuliá F, Pires AS, Gasparotto J, Terra SR and Moreira JC:
Vitamin A (retinol) downregulates the receptor for advanced
glycation endproducts (RAGE) by oxidant-dependent activation of p38
MAPK and NF-κB in human lung cancer A549 cells. Cell Signal.
25:939–954. 2013. View Article : Google Scholar : PubMed/NCBI
|