Introduction
Tissue engineering is an interdisciplinary field
that focuses on developing biological substitutes that restore,
maintain or improve natural tissue function (1). However, tissues are sophisticated
structures formed by multiple cells types acting in concert under
regulated conditions. In this context, one of the major
difficulties in tissue engineering is acquiring sufficient numbers
of cells. For this reason, researchers have focused on improving
the differentiation potential of stem cells to accelerate organ
regeneration.
Mechanical stimulation affects stem cell
differentiation. Exposing cells to real or simulated microgravity
(SMG) is a suitable technique for tissue engineering, as
microgravity triggers cells to assemble three-dimensionally in
vitro as they do in vivo (2–4).
Human mesenchymal stem cells (hMSCs) cultured in SMG possess the
proliferative characteristics of stem cells and retain their
ability to differentiate into hyaline cartilage following
transplantation (5). Huang et
al (6) reported that
microgravity induced MSCs to differentiate into adipocytes, which
are force-insensitive cells. Chen et al (7) demonstrated that MSCs more readily
differentiated into neuronal cells under SMG. However, modeled
microgravity inhibited the differentiation of MSCs into osteoblasts
(8). The functional mechanisms
underlying the effect of SMG on MSC differentiation remains
unclear.
Cytoskeletal tension and ras homolog family member A
(RhoA) have previously been demonstrated to mediate the lineage
decision of hMSCs (9). Increased
actomyosin contractility promotes osteogenesis, while cytoskeleton
disruption enhances adipogenesis (10). During spaceflight and under SMG
conditions, cytoskeletal alterations occur in several cell types
including lymphocytes, glial cells and stem cells (11–13).
In addition, previous research demonstrated that MSCs are sensitive
to gravity changes (6). MSC
microtubules and α-actin undergo remodeling following SMG
stimulation (7). The Rho subfamily
of Ras-related small GTPases is characterized predominantly by its
regulation of the actin cytoskeleton. RhoA primarily mediates
stress fiber formation (14).
Thus, the present study examined whether SMG remodels the
cytoskeleton by regulating the activity of RhoA. The purpose of the
present study was to measure the effect of time-dependent changes
in the cytoskeleton of MSCs on MSC differentiation following
different conditions of modeled microgravity.
Materials and methods
Preparation and culture of adult rat
(r)MSCs
A total of 10 male Wistar rats (two weeks old) were
purchased from the animal research center of the Fourth Military
Medical University (Xi'an, China). The rats were housed in isolated
and ventilated cages, and the animal protocol was approved by the
Fourth Military Medical University Medical Ethics Committee (Xi'an,
Shaanxi, China; approval no. XJYYLL-2015508). The animals were
acclimated to the laboratory environment for 5–7 days before use.
While in their home cage environment, the animals were allowed
access to a standard animal diet and tap water ad libitum. The room
was maintained at 20–23°C with a 12 h:12 h light:dark cycle. The
animals were deeply anesthetized using inhaled isoflurane (1–3% or
as required) before all surgery. All efforts were made to minimize
animal suffering and to reduce the number of animals used. rMSCs
were collected as previously described by Azizi et al
(15), with slight modifications.
The Wistar rats were euthanized and bilateral femurs and tibias
were removed. Mesenchymal stem cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin G
and 100 mg/ml streptomycin (Gibco; Thermo Fisher Scientific, Inc.)
at 2.5×105/cm2. Fluorescence phase-contrast
microscopy (Axiocam MR R3; Carl Zeiss AG, Oberkochen, Germany) was
used to observe rMSCs every 2–3 days. Fluorescence activated cell
sorting (FACS) analysis of CD44, CD34, CD90 and CD45 was performed
in rMSCs.
Clinorotation to modeled
microgravity
The weightless environment from a clinostat, or
rotating vessel, is regarded as ‘modeled microgravity’ (16). The conditions prevent the cell from
being affected by gravity. The method is based on the hypothesis
that sensing no gravity has similar effects to being weightless.
The clinostat model system (clinorotation) used in the present
study provides a vector-averaged reduction in the apparent gravity
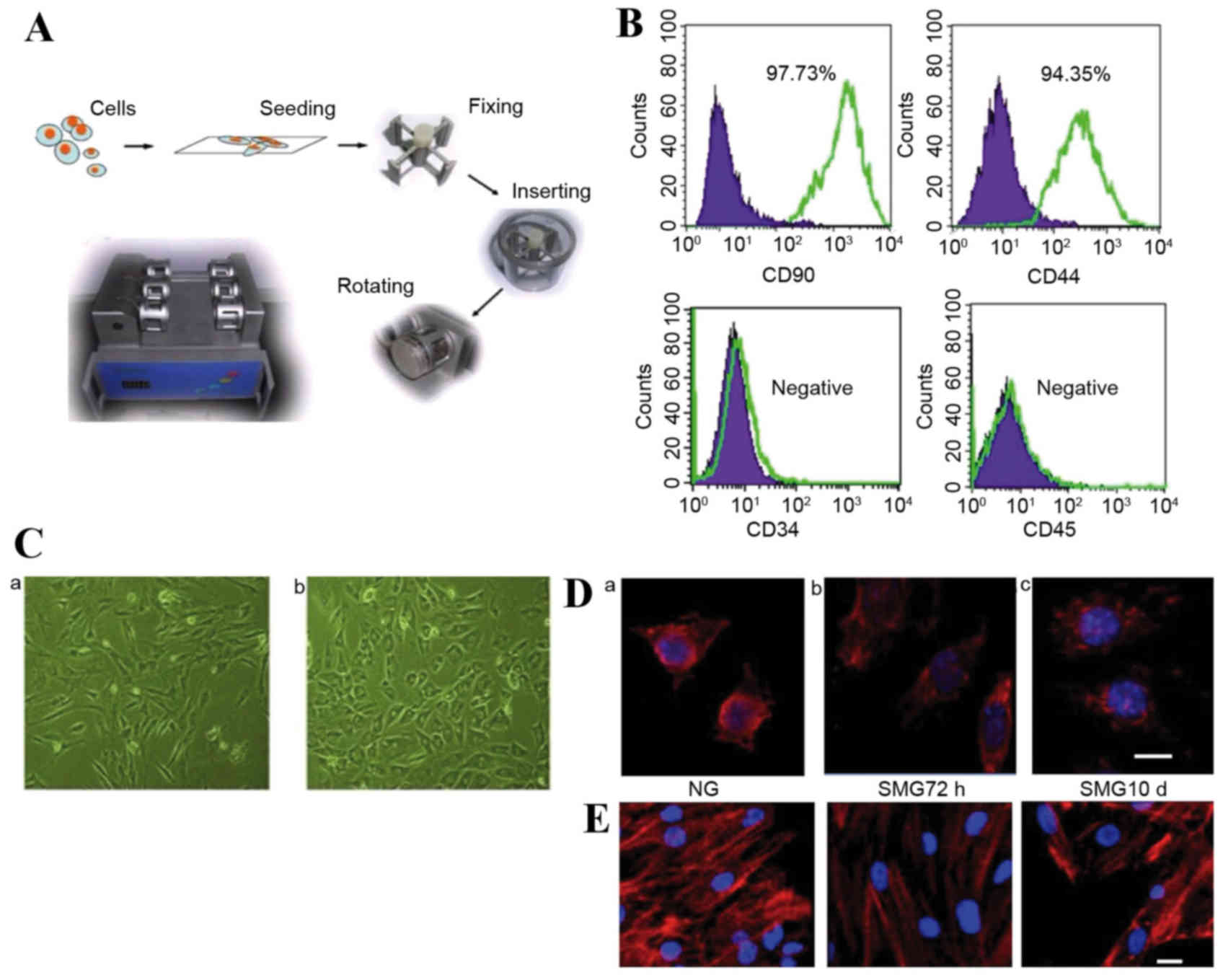
on cell culture (Fig. 1A). Stem
cells were seeded at a density of 1×105 cells on 2.5×2.5
cm coverslips. The coverslips were inserted into the fixture of the
chambers, filled completely with DMEM + 10% FBS and aspirated to
eliminate air bubbles. The chambers were divided into two groups
randomly: Normal gravity (NG) controls without rotation for 72 h
and SMG groups with clinorotation (30 × g) for 72 h or 10 days.
Neuronal differentiation
Following SMG stimulation, rMSCs were induced to
differentiate as previously described, with modifications (17). Cells were maintained in
pre-induction media consisting of DMEM, 0.1 mM/l 2-mercaptoethanol
(Gibco; Thermo Fisher Scientific, Inc.) and 2% dimethylsulfoxide
(WAK-Chemie Medical GmbH, Steinbach, Germany) for 5 h.
Pre-induction media were removed, cells were transferred to
neuronal induction media composed of DMEM + 10% FBS, 10 µg/l basic
fibroblast growth factor (R&D Systems, Inc., Minneapolis, MN,
USA), 10 µg/l human epidermal growth factor (R&D Systems,
Inc.), 1 mM dibutyrylcyclicn AMP (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) and 0.5 mM 3-isobutyl-1-methyl-xanthine (IBMX;
Sigma-Aldrich; Merck KGaA) for 10 days.
Osteogenic differentiation
Induced osteogenic differentiation of MSCs was
performed according to the accepted protocol of culturing cells in
DMEM-advanced medium containing 10−8 M dexamethasone, 10
mM beta-glycerol 2-phosphate and 0.2 mM 2-phospho-L-ascorbic acid
(Sigma-Aldrich; Merck KGaA) (18).
Endothelial differentiation
Endothelial differentiation was performed as
described previously with modifications (19). Cells were incubated for up to 10
days in endothelial differentiation medium containing low glucose
DMEM supplemented with 100 ng/ml vascular endothelial growth factor
(Invitrogen; Thermo Fisher Scientific, Inc.), 50 ng/ml epidermal
growth factor (Invitrogen; Thermo Fisher Scientific, Inc.), 1 µg/ml
hydrocortisone (Sigma-Aldrich; Merck KGaA), 5% FBS (Invitrogen;
Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and 100 mg/ml
streptomycin.
Adipogenic differentiation of
MSCs
Following culture under simulated microgravity for
72 h or 10 days, cells were cultured in adipogenic medium with 1 µM
dexamethasone, 10 µg/ml insulin, 500 µM IBMX and 200 µM
indomethacin (20).
Fluorescence immunocytochemistry
Following SMG stimulation, the cytoskeletons of MSCs
were observed. Cultured cells were fixed with 4% paraformaldehyde
for 20 min, immersed in PBS for 10 min then exposed to 0.01% Triton
X-100 at room temperature for 10 min. Following rotation for 72 h
or 10 days, rMSCs were blocked with 1% FBS at room temperature for
30 min then incubated with Texas Red-labeled phalloidin (T7471;
1:200; Invitrogen; Thermo Fisher Scientific, Inc.) and β-tubulin
(SC398937; 1:200; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
antibodies at 37°C for 1 h. MSCs were stained with
4′6′-diamidino-2-phenylindole (1:500; Sigma-Aldrich; Merck KGaA)
for 15 min for nuclear staining. To further examine the involvement
of RhoA activation in the differentiation potential of rMSCs
cultured in SMG, Y27632 (040012; Stemgent, Cambridge, MA, USA) was
used to specifically block RhoA/rock signaling. Cells were
pretreated with 20 µml inhibitor and stimulated with SMG for 10
days. ALP (LS-C44264; 1:200; LifeSpan BioSciences, Inc., Seattle,
WA, USA) antibody was incubated as above. Cells were observed on a
laser scanning confocal microscope (Leica Microsystems GmbH,
Wetzlar, Germany) and imaged using a SPOT II camera (Diagnostic
Instruments, Inc., Sterling Heights, MI, USA).
Reverse transcription polymerase chain
reaction (RT-PCR) analysis
Total RNA was extracted from cultured cells in
induction media using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
primer sequences used for microtubule-associated protein 2 (MAP-2),
von Willebrand factor (vWF), alkaline phosphatase (ALP), peroxisome
proliferator-activated receptor γ2 (PPARγ-2) and GAPDH are
presented in Table I and were
synthesized by Beijing SBS Genetech Co., Ltd. (Beijing, China).
RT-PCR using the Access RT-PCR reagent (Promega Corporation,
Madison, WI, USA) was performed for 35 cycles, with each cycle
consisting of 94°C for 30 sec, 55°C for 30 sec and 68°C for 1 min.
Samples (10 µl) of each PCR product were size-fractionated by 1.5%
agarose gel electrophoresis and the bands were visualized with
ImageMaster VDS (Pharmacia Biotech; GE Healthcare Life Sciences,
Chalfont, UK). The densitometry values were normalized to GAPDH and
performed by Total Lab computer program version 1.11 (GE Healthcare
Life Sciences, Chalfont, UK).
 | Table I.Primers used in the semi-quantitative
reverse transcription-polymerase chain reaction. |
Table I.
Primers used in the semi-quantitative
reverse transcription-polymerase chain reaction.
| Primer | Gene sequence
(5′-3′) | Length (bp) |
|---|
| MAP-2 | Forward
TTGAAGGTTAAAATGCATCTGA | 152 |
|
| Reverse
GGCATTTCAAGGAAAAACTCA |
|
| vWF | Forward
CACCCAAGTTCCACATCAGC | 120 |
|
| Reverse
TCCACCATCGTCTGCTTCAT |
|
| ALP | Forward
GGAAGGGTCAGTCAGGTT | 366 |
|
| Reverse
GTGGGCCGCTCTAGGCACCAA |
|
| PPARγ2 | Forward
5′-TTGATTTCTCCAGCATTTC-3′ | 360 |
|
| Reverse
GCTCTACTTTGATCGCACT |
|
| GAPDH | Forward
5′-CCACAGTCCATGCCATCAC-3′ | 452 |
|
| Reverse
TCCACCACCCTGTTGCTGTA |
|
Oil red o staining
Oil red O kit (ab150678; Abcam, Cambridge, UK) was
used to investigate the efficiency of adipogenic induction. Cells
were fixed in 10% formalin for 10 min, rinsed 3 times in 1X PBS,
then placed in oil red O solution for 10 min, rinsed in tap water
and counterstained with hematoxylin for 1 min. An inverted
phase-contrast microscope (Eclipse TE 300; Nikon Co., Tokyo, Japan)
was used for examination.
FACS
Following induction for 10 days, the differentiated
cells were analyzed using FACS. The harvested cells washed in
Dulbecco's PBS and incubated at 4°C for 30 min with antibodies for
MAP-2 (13–1500; 1:200; Invitrogen; Thermo Fisher Scientific, Inc.),
vWF (MA5-14029; 1:200; Invitrogen; Thermo Fisher Scientific, Inc.),
ALP (LS-C44264; 1:200; LifeSpan BioSciences, Inc.) and PPARγ-2
(PA3-821A; 1:200; Invitrogen; Thermo Fisher Scientific, Inc.)
following blocking with normal goat serum (10%) at 4°C for 30 min.
Cells were resuspended in DMEM/PBS containing a working dilution
(1:100) of phycoerythrin-labeled goat anti-mouse immunoglobulin G
(43R-1613; Fitzgerald, Acton, MA, USA) and incubated at 4°C for 30
min. Cells were analyzed using a fluorescence activated cell sorter
(FACScan) cytometer (Cytoflex S; Beckman Coulter Inc., Brea, CA,
USA) with FlowJo software version 7.6 (Tree Star Inc, Ashland, OR,
USA).
RhoA activity assay
The RhoA activity assay was performed according to
the manufacturer's protocol (RhoA Pulldown Activation Assay kit;
BK036; Cytoskeleton, Inc., Denver, CO, USA). Activated (GTP-bound)
RhoA was pulled-down from whole cell lysates (300 µg
protein/sample) with rhotekin-conjugated agarose beads for 1 h at
4°C. The beads were collected by centrifugation for 1 min at 5,000
× g at 4°C and washed with lysis buffer, followed by wash buffer
(1X lysis buffer). Activated RhoA was detached from the beads by
boiling for 5 min in Laemmli reducing buffer (1X) immediately prior
to separation by 12.5% SDS-PAGE, following which it was transferred
to a polyvinylidene membrane. Following blocking for 1 h at room
temperature in TBST containing 5% non-fat dry milk, the membranes
were incubated with a primary antibody directed against RhoA
(21009; 1:200; Cytoskeleton, Inc.) overnight at 4°C. Signals were
detected using a horseradish peroxidase-conjugated goat anti-mouse
immunoglobulin G1 secary antibody incubated for 1 h at room
temperature (R-05071-500; 1:1,000; Advansta, Menlo Park, CA, USA)
and an enhanced chemiluminescence detection system (GE Healthcare
Life Sciences). The optical densities were analyzed by using
ImageMaster 2D Platinum version 5.0 (GE Healthcare Life
Sciences).
Statistical analysis
Statistical analysis was carried out using SPSS
version 10.0 for Window software (SPSS, Chicago, IL, USA).
Statistical analysis was performed using the Student's t-test on
pixel intensity data generated from scanning autoradiographs.
P<0.05 was considered to indicate a statistically significant
difference. All experiments were repeated at least three times.
Data were expressed as the mean ± standard error of the mean.
Results
Characterization of cultured
rMSCs
Flow cytometry analysis was performed on cells at
passage 2. Undifferentiated rMSCs expressed CD90 and CD44, with a
positive ratio of 97.37 and 94.35%, respectively. These cells were
negative for CD45 and CD34 (Fig.
1B). Following 2 passages, adherent cells exhibited spindle or
large flattened morphology, and following SMG treatment the shape
of the cells visibly shifted from spindle to round (Fig. 1C). This morphological change may
reflect cytoskeletal reorganization; therefore, the cytoskeletal
change following SMG intervention was analyzed at different time
points.
Cytoskeletal reorganization following
SMG stimulation
In the present study, MSC microtubules were highly
sensitive to weightlessness. Modified microtubules were observed in
some MSC clusters under SMG, and few fibers were distinguishable
against the background under SMG for 72 h (Fig. 1D). However, following 10 days of
SMG culture, cells appeared to have reestablished more microtubules
(Fig. 1D). Compared with the
static control, α-actin filaments underwent remodeling following
SMG stimulation, appearing diffuse following 72 h rotation
(Fig. 1E). However, following
rotation for 10 days, re-concentrated actin was observed (Fig. 1E).
Neuronal differentiation
MSCs were cultured using a clinostat to simulate
microgravity conditions for 0 h (NG), 72 h or 10 days. Following
SMG, cells were incubated for up to 10 days in differentiation
medium. At day 10, cells were lysed with TRIzol for total RNA
extraction. The expression of the neuron-specific marker MAP-2 was
analyzed by semi-quantitative RT-PCR. The results revealed that
expression of MAP-2 increased significantly following 72 h SMG
compared with the NG group (P<0.05; Fig. 2A). In contrast, expression
significantly decreased following 10 days SMG stimulation compared
with the 72 h group (P<0.05; Fig.
2A).
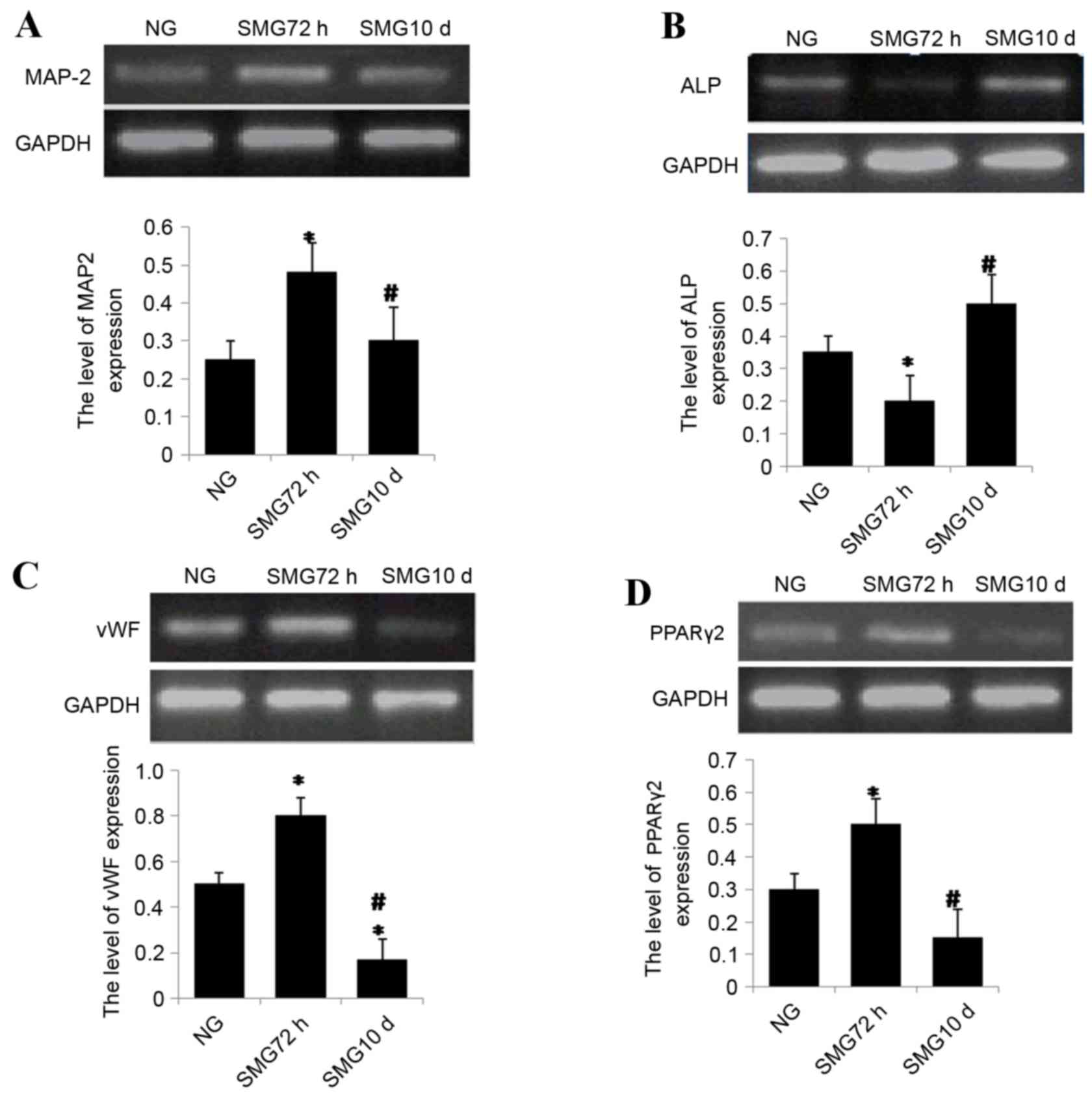 | Figure 2.(A) Semi quantitative reverse
transcription polymerase chain reaction analysis of (A) MAP-2
following neuronal differentiation, (B) ALP following osteogenic
differentiation, (C) vWF following endothelial differentiation and
(D) PPARγ2 expression following adipogenic differentiation. Gene
expression was normalized to endogenous GAPDH. The results
represent three independent experiments. *P<0.05 vs. NG group,
#P<0.05 vs. 72 h. MAP-2, microtubule-associated
protein 2; ALP, alkaline phosphatase; vWF, von Willebrand factor;
PPARγ2, peroxisome proliferator-activated receptor γ2; GAPDH,
glyceraldehyde 3-phosphate dehydrogenase; NG, normal gravity; SMG,
simulated microgravity. |
Osteogenic differentiation
mRNA expression levels of ALP, a marker of
osteogenesis, were investigated. The results revealed that ALP
expression was reduced significantly following 72 h exposure to SMG
compared with the NG group (P<0.05; Fig. 2B). ALP mRNA expression levels
significantly increased following 10 day SMG exposure compared with
72 h SMG (P<0.05; Fig. 2B).
Endothelial differentiation
rBMSCs were cultured under SMG conditions for 72 h
or 10 days, then further cultured for 10 days in differential
medium. The expression level of the endothelium- specific marker
vWF was analyzed by semi-quantitative RT-PCR. vWF mRNA expression
levels were significantly higher in the 72 h group compared with
the NG group, and four-fold higher in the 72 h group compared with
10 day group (both P<0.05; Fig.
2C).
Adipogenic differentiation
PPARγ-2 is a critical transcription factor involved
in adipogenic differentiation in MSCs. To measure the potential of
differentiation into adipocytes, PPARγ-2 mRNA expression levels
were measured by semi-quantitative RT-PCR in MSCs under simulated
microgravity. The results revealed that mRNA expression levels of
PPARγ-2 increased significantly following 72 h SMG compared with
the NG group (P<0.05; Fig. 2D).
Notably, following long term stimulation for 10 days, PPARγ-2 mRNA
expression levels significantly decreased by ~60% compared with 72
h group (P<0.05; Fig. 2D).
Confirmation of differentiation by
FACS
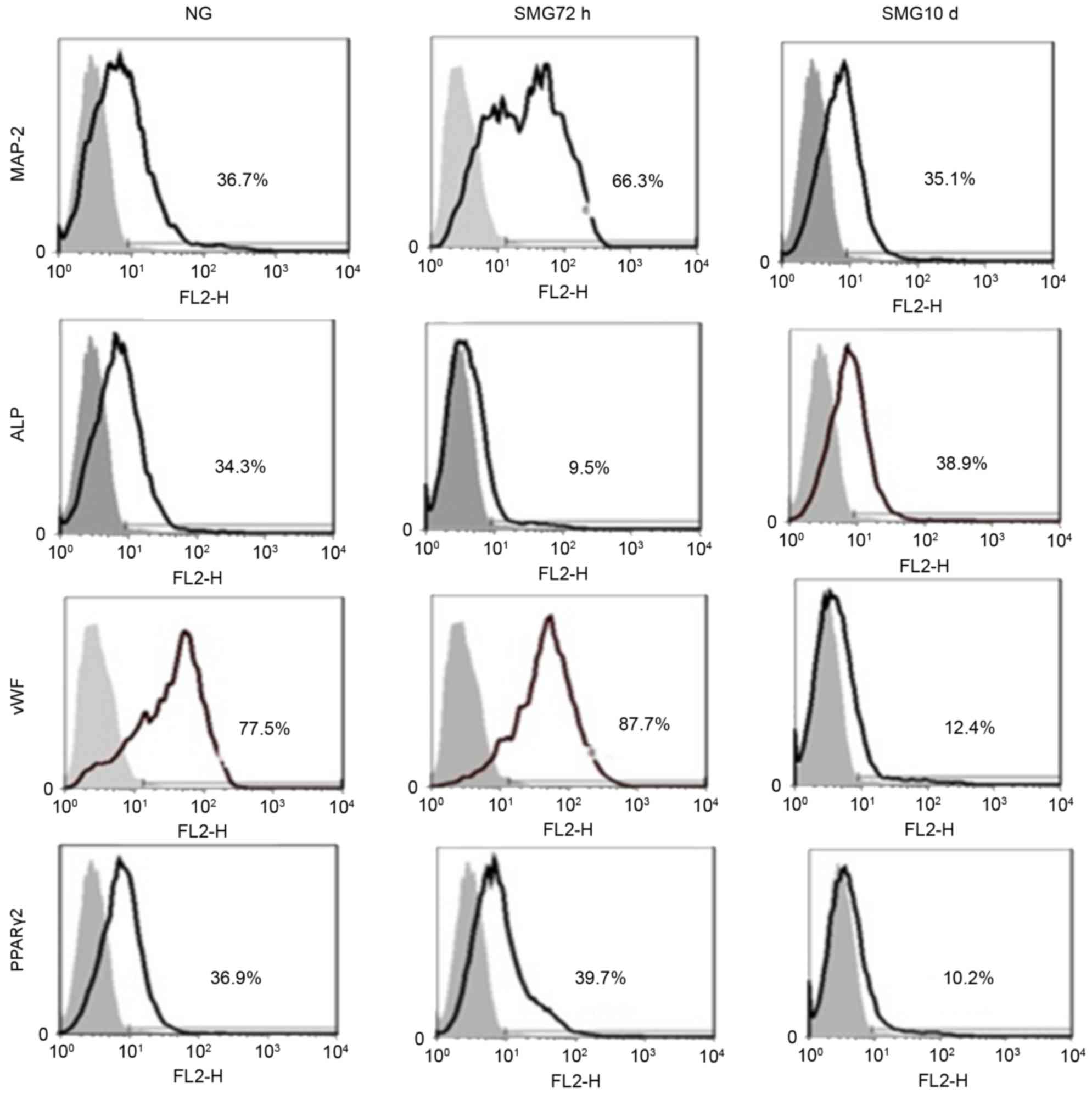
The FACS results revealed that MAP-2-positive cells
from the SMG 72 h group accounted for 66.3% of the cells, compared
with the NG group (36.7%) and SMG 10 day group (35.1%; Fig. 3), confirming that a shorter period
of SMG induces MSCs to differentiate into neuronal cells.
ALP-positive cells in the SMG 10 day group accounted for 38.9±2.03%
of cells, compared with the NG group (34.3±1.28%) and SMG 72 h
group (9.5±1.12%; Fig. 3),
suggesting that an extended period of SMG induces MSCs to
differentiate into osteogenic cells. vWF-positive cells in the SMG
72 h group accounted for 87.7±2.34% of cells, compared with the NG
group (77.5±1.74%) and SMG 10 day group (12.4±1.16%; Fig. 3), suggesting that a short period of
SMG induces MSCs to differentiate into endothelial cells.
PPARγ-2-positive cells in the SMG 72 h group accounted for
39.7±2.01% of cells compared to the NG group (36.9±1.18%) and SMG
10 day group (10.2±1.11%; Fig. 3).
These results indicated shorter period of SMG resulted in the
induction of MSCs to differentiate into adipogenic cells.
Effects of microgravity on RhoA
activity
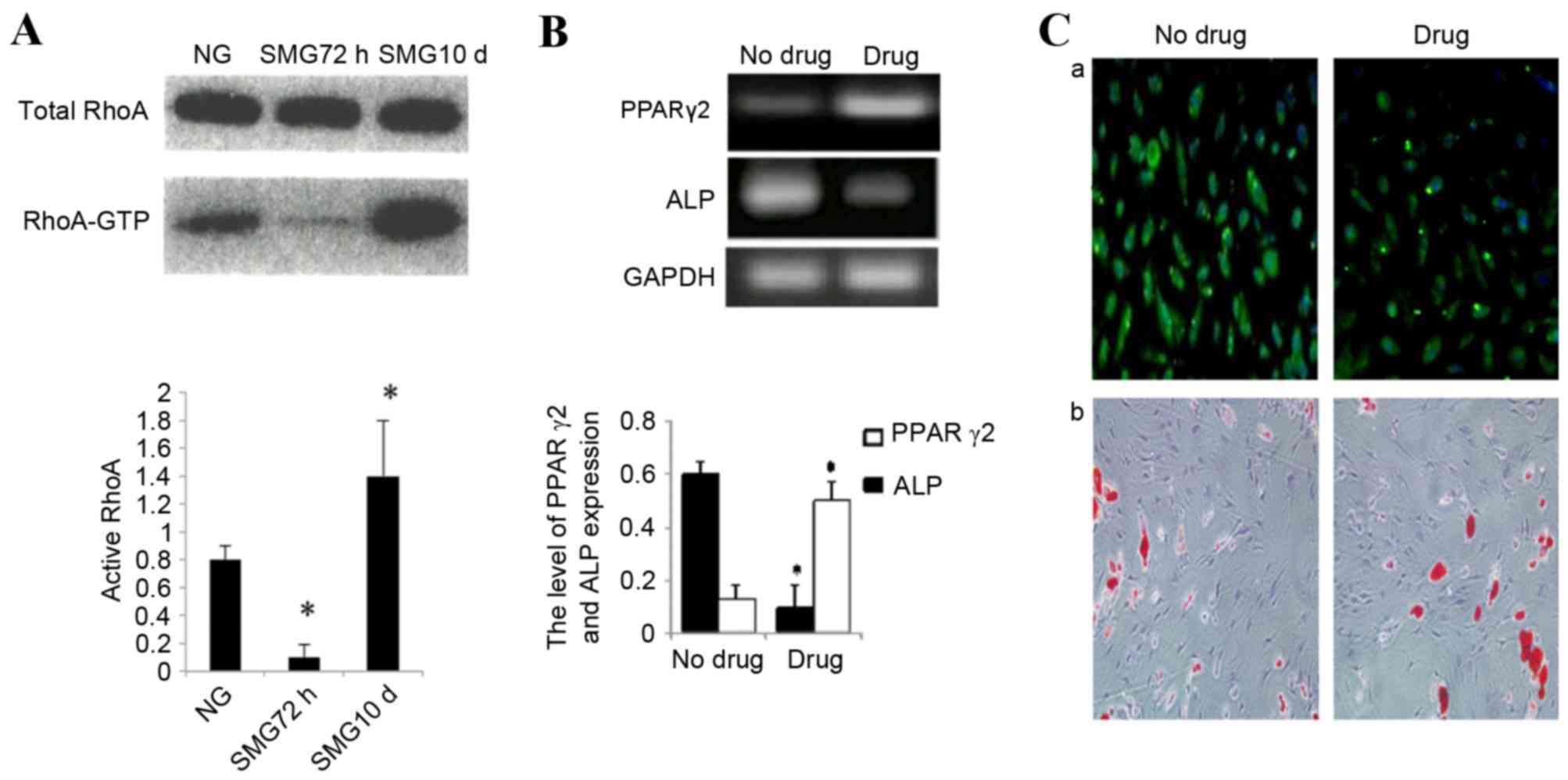
Total RhoA protein expression was confirmed by
western blot analysis. No significant change was observed in total
RhoA protein expression between control cells cultured in NG or
SMG. Activation of the small GTPase, RhoA, is required for both
stress fiber formation and integrin-mediated signaling (14). In addition, RhoA has been
implicated in the lineage decision of hMSCs (21). Therefore, the activation status of
RhoA in cells cultured in SMG was examined. In the 72 h group, an
83±3% reduction in the activated GTP-bound form of RhoA was
observed compared with the NG control group (P<0.05; Fig. 4A). This was consistent with reduced
stress fiber formation. As MSCs were exposed to SMG for 10 days,
the level of activated RhoA increased nine-fold compared with the
72 h group (P<0.05; Fig.
4A).
Taken together, these data indicated that a high
level of active RhoA is associated with osteogenic conditions and a
low level with adipogenic, neuronal and endothelial conditions. To
further examine the involvement of RhoA activation in the
differentiation potential of rMSCs cultured in SMG, Y27632 (040012;
Stemgent) was used to specifically block RhoA/rock signaling. Cells
were pretreated with inhibitor and stimulated with SMG for 10 days.
MSCs were subsequently cultured in adipogenic medium or osteogenic
medium separately. The inhibitor group was demonstrated to abrogate
osteogenic media-induced osteogenesis, and redirected cells to
differentiate into adipogenic cells. The expression of ALP
decreased significantly by 80% in inhibited cells compared with the
untreated control (P<0.05; Fig. 4B
and C). Finally, PPARγ-2 expression increased ~3 times in the
inhibited group compared with the untreated control group
(P<0.05; Fig. 4B and C).
Discussion
Bone marrow mesenchymal stem cells have demonstrated
potential in tissue engineering and regenerative medicine (22). However, multiple issues still
remain to be solved, including stem cell differentiation
efficiency, acquiring sufficient seed cells and generating
sufficient numbers of transplantation stem cells. Over the past 40
years, space exploration has revealed that microgravity has a major
impact on biology. Previous studies have demonstrated that
microgravity suppresses bone marrow mesenchymal stem cell
differentiation toward osteoblasts, but promotes adipogenic
differentiation (8,23). Contrary to those findings,
Buravkova et al (24)
demonstrated that culture of hMSCs in microgravity for 10 days
promoted osteogenic differentiation. Therefore, the effect of
microgravity on stem cells is a controversial topic that requires
further research attention.
The present study revealed that different durations
of SMG had different effects on the cytoskeleton. Short term 72 h
SMG decreased cytoskeletal tension and resulted in microtubule
disintegration. In comparison, longer term SMG for 10 days
increased cytoskeletal tension and tubulin maintained
polymerization activity. Given that RhoA is involved in the
regulation of the cytoskeleton, RhoA expression and activity were
analyzed. Short term SMG decreased RhoA activity significantly,
however, prolonged microgravity for 10 days significantly increased
RhoA activity.
The effect of microgravity on bone marrow
mesenchymal stem cell differentiation was also examined. A shorter
period of SMG promoted MSCs to differentiate into endothelial,
neuronal and adipogenic cells. In comparison, a longer period of
SMG promoted MSCs to differentiate into osteoblasts. When RhoA
activity was inhibited, the results revealed that the effect of
longer SMG exposure was reversed. These findings highlight a clear
distinction of variable microgravity duration driving bone marrow
mesenchymal stem cell differentiation to different fates; one to
stress-sensitive identities (including osteoblastic cells and
myocardial cells), and the other to cells not sensitive to stress
(including endothelial, neuronal and adipogenic cells). It has
previously been demonstrated that microgravity inhibits the
differentiation of osteoblasts from bone marrow mesenchymal stem
cells (8). However, this is
contradictory to the results of the present study, which revealed
that a shorter period of SMG inhibits osteoblast differentiation,
and a longer period of SMG supports osteoblast differentiation.
Spiegelman and Ginty (21) reported that changes in cell shape
modulated the differentiation degree and direction. McBeath et
al (25) demonstrated that
cytoskeletal tension regulated stem cell differentiation fate. RhoA
is an important molecule in cytoskeletal regulation, and has been
reported to promote smooth and skeletal muscle cell division
(26). Sordella et al
(27) demonstrated that, following
elimination of RhoA by inactivating the locus, the differentiation
ability of embryonic fibroblast cells into osteoblasts was
enhanced, but adipogenic differentiation was weakened. These
findings are consistent with the results of the present study.
RhoA is involved in cell stress fiber assembly. The
most critical downstream effectors are Rho-associated,
coiled-coil-containing protein kinase (ROCK) and mammalian
diaphanous related protein (mDia), which are involved in the
process of stress assembly (28).
ROCK phosphorylates LIM domain kinase (LIMK) (29). LIMK, in turn, phosphorylates
cofilin, a small actin-binding protein that promotes F-actin
depolymerization. Upon phosphorylation, the actin-binding ability
of cofilin is reduced, and actin polymerization is stabilized
(29). mDia supports parallel
stress fibers to help accumulate actin. It is also involved in the
mediation of microtubule assembly and dynamic balance (30). Therefore, the effect of
microgravity on cytoskeletal tension was speculated to be via the
RhoA-associated pathway. A short period of SMG, which downregulates
cytoskeletal tension, may inhibit RhoA to promote differentiation
into cells not sensitive to stress. In comparison, a longer period
of SMG upregulates cytoskeletal tension and activates RhoA to
enhance stress-sensitive cell differentiation. Therefore, different
periods of SMG stimulation may be an important factor that drives
differentiation fates of MSCs.
In conclusion, microgravity is an important factor
that affects the differentiation of rBMSCs. The findings of the
present study underscore the previously reported potential of MSCs
for regenerative medicine, tissue engineering and stem cell-based
therapy. Future in vivo studies are required to confirm and
clarify these findings, given that these results were obtained via
in vitro simulated experiments.
Acknowledgements
The authors would like to thank Dr Austin Cape at
ASJ Editors for careful reading and feedback. Confocal laser
scanning microscopy was performed at the Department of Anatomy and
K.K. Leung Brain Research Center, The Fourth Military Medical
University (Xi'an, China). The present study was funded by the
National Natural Science Foundation of China (grant no. 81402055),
Natural Science Foundation of Shaanxi Province (grant no.
2016JM8014).
Glossary
Abbreviations
Abbreviations:
|
SMG
|
simulated microgravity
|
|
NG
|
normal gravity
|
|
MSCs
|
mesenchymal stem cells
|
|
ROCK
|
Rho-associated, coiled-coil-containing
protein kinase
|
|
mDia
|
mammalian diaphanous related
protein
|
|
PPARγ2
|
peroxisome proliferator-activated
receptor γ2
|
|
MAP-2
|
microtubule-associated protein 2
|
|
Vwf
|
von Willebrand factor
|
|
ALP
|
alkaline phosphatase
|
References
|
1
|
Langer R and Vacanti JP: Tissue
engineering. Science. 260:920–926. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Grimm D, Bauer J, Kossmehl P, Shakibaei M,
Schöberger J, Pickenhahn H, Schulze-Tanzil G, Vetter R, Eilles C,
Paul M and Cogoli A: Simulated microgravity alters differentiation
and increases apoptosis in human follicular thyroid carcinoma
cells. FASEB J. 16:604–606. 2002.PubMed/NCBI
|
|
3
|
Hochleitner B, Hengster P, Duo L, Bucher
H, Klima G and Margreiter R: A novel bioartificial liver with
culture of porcine hepatocyte aggregates under simulated
microgravity. Artif Organs. 29:58–66. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marquette ML, Byerly D and Sognier M: A
novel in vitro three-dimensional skeletal muscle model. In Vitro
Cell Dev Biol Anim. 43:255–263. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yuge L, Kajiume T, Tahara H, Kawahara Y,
Umeda C, Yoshimoto R, Wu SL, Yamaoka K, Asashima M, Kataoka K and
Ide T: Microgravity potentiates stem cell proliferation while
sustaining the capability of differentiation. Stem Cells Dev.
15:921–929. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang Y, Dai ZQ, Ling SK, Zhang HY, Wan YM
and Li YH: Gravity, a regulation factor in the differentiation of
rat bone marrow mesenchymal stem cells. J Biomed Sci. 16:872009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen J, Liu R, Yang Y, Li J, Zhang X, Wang
Z and Ma J: The simulated microgravity enhances the differentiation
of mesenchymal stem cells into neurons. Neurosci Lett. 505:171–175.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dai ZQ, Wang R, Ling SK, Wan YM and Li YH:
Simulated microgravity inhibits the proliferation and osteogenesis
of rat bone marrow mesenchymal stem cells. Cell Prolif. 40:671–684.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
McBeath R, Pirone DM, Nelson CM,
Bhadriraju K and Chen CS: Cell shape, cytoskeletal tension, and
RhoA regulate stem cell lineage commitment. Dev Cell. 6:483–495.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kilian KA, Bugarija B, Lahn BT and Mrksich
M: Geometric cues for directing the differentiation of mesenchymal
stem cells. Proc Natl Acad Sci USA. 107:4872–4877. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hughes-Fulford M and Lewis ML: Effects of
microgravity on osteoblast growth activation. Exp Cell Res.
224:103–109. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schatten H, Lewis ML and Chakrabarti A:
Spaceflight and clinorotation cause cytoskeleton and mitochondria
changes and increases in apoptosis in cultured cells. Acta
Astronaut. 49:399–418. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Uva BM, Masini MA, Sturla M, Prato P,
Passalacqua M, Giuliani M, Tagliafierro G and Strollo F:
Clinorotation-induced weightlessness influences the cytoskeleton of
glial cells in culture. Brain Res. 934:132–139. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hall A: Rho GTPases and the actin
cytoskeleton. Science. 279:509–514. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Azizi SA, Stokes D, Augelli BJ, DiGirolamo
C and Prockop DJ: Engraftment and migration of human bone marrow
stromal cells implanted in the brains of albino rats-similarities
to astrocyte grafts. Proc Natl Acad Sci USA. 95:3908–3913. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang X, Ren Q, Fu S and Jiang P:
Development and application of clinostat for simulation of
microgravity biological effects. Acta Biophysica Sinica.
13:161–166. 1997.
|
|
17
|
Jiang J, Lv Z, Gu Y, Li J, Xu L, Xu W, Lu
J and Xu J: Adult rat mesenchymal stem cells differentiate into
neuronal-like phenotype and express a variety of neuro-regulatory
molecules in vitro. Neurosci Res. 66:46–52. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jaiswal N, Haynesworth SE, Caplan AI and
Bruder SP: Osteogenic differentiation of purified, culture-expanded
human mesenchymal stem cells in vitro. J Cell Biochem. 64:295–312.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Oswald J, Boxberger S, Jorgensen B,
Feldmann S, Ehninger G, Bornhäuser M and Werner C: Mesenchymal stem
cells can be differentiated into endothelial cells in vitro. Stem
Cells. 22:377–384. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xiang Y, Zheng Q, Jia B, Huang G, Xie C,
Pan J and Wang J: Ex vivo expansion, adipogenesis and neurogenesis
of cryopreserved human bone marrow mesenchymal stem cells. Cell
Biol Int. 31:444–450. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Spiegelman BM and Ginty CA: Fibronectin
modulation of cell shape and lipogenic gene expression in
3T3-adipocytes. Cell. 35:657–666. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Baksh D, Song L and Tuan RS: Adult
mesenchymal stem cells: Characterization, differentiation, and
application in cell and gene therapy. J Cell Mol Med. 8:301–316.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zayzafoon M, Gathings WE and McDonald JM:
Modeled microgravity inhibits osteogenic differentiation of human
mesenchymal stem cells and increases adipogenesis. Endocrinology.
145:2421–2432. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Buravkova L, Romanov Y, Rykova M,
Grigorieva O and Merzlikina N: Cell-to-cell interactions in changed
gravity: Ground-based and flight experiments. Acta Astronaut.
57:67–74. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
McBeath R, Pirone DM, Nelson CM,
Bhadriraju K and Chen CS: Cell shape, cytoskeletal tension, and
rhoa regulate stem cell lineage commitment. Dev Cell. 6:483–495.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Carnac G, Primig M, Kitzmann M, Chafey P,
Tuil D, Lamb N and Fernandez A: RhoA GTPase and serum response
factor control selectively the expression of MyoD without affecting
Myf5 in mouse myoblasts. Mol Biol Cell. 9:1891–1902. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sordella R, Jiang W, Chen GC, Curto M and
Settleman J: Modulation of Rho GTPase signaling regulates a switch
between adipogenesis and myogenesis. Cell. 113:147–158. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Young KG and Copeland JW: Formins in cell
signaling. Biochim Biophys Acta. 1803:183–190. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Maekawa M, Ishizaki T, Boku S, Watanabe N,
Fujita A, Iwamatsu A, Obinata T, Ohashi K, Mizuno K and Narumiya S:
Signaling from Rho to the actin cytoskeleton through protein
kinases ROCK and LIM-kinase. Science. 285:895–898. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hotulainen P and Lappalainen P: Stress
fibers are generated by two distinct actin assembly mechanisms in
motile cells. J Cell Biol. 173:383–394. 2006. View Article : Google Scholar : PubMed/NCBI
|