Introduction
Mitochondria, which are multifunctional organelles,
are important in cellular proliferation and physiology, including
cellular energy (adenosine triphosphate) production via oxidative
phosphorylation and apoptosis via cytochrome c release
(1). Mitochondrial dysfunction is
linked to several human diseases, including premature aging,
diabetes mellitus and cancer (2).
Differences in the number, morphology and function of mitochondria
have been detected between normal cells and cancer cells (3). Previous studies have revealed that
mitochondrial dysfunction contributes to the development and
progression of cancer (4).
The epithelial-mesenchymal transition (EMT) is a
process by which cells undergo a morphological switch from the
epithelial polarized phenotype to the mesenchymal fibroblastoid
phenotype. During EMT, the function and expression of the
epithelial cell-cell adhesion molecule E-cadherin is lost, whereas
the expression of the mesenchymal cell-cell adhesion molecule
N-cadherin is induced (5). Numerous
studies have previously demonstrated that EMT is often activated
during cancer cell migration, invasion, metastatic dissemination
and chemoresistance (6). However,
changes in mitochondrial function during EMT are largely unknown.
In the present study, the mitochondrial function was examined
during EMT induced by transforming growth factor (TGF)β-1 exposure
in pancreatic cancer. Mitochondrial DNA (mtDNA), mitochondrial mass
and reactive oxygen species (ROS) increased, while the
mitochondrial membrane potential (ΔΨm) decreased, following TGFβ-1
treatment in pancreatic cancer.
Materials and methods
Cell culture and EMT induction
The pancreatic cell line Panc-1 was purchased from
the Type Culture Collection of Chinese Academy of Sciences
(Shanghai, China), and was cultured in RPMI 1640 medium (Jinuo,
Hangzhou, China) containing 10% fetal bovine serum (HyClone; GE
Healthcare Life Sciences, Logan, UT, USA) 1 µg/ml streptomycin
(Jinuo), 100 IU/ml penicillin (Jinuo), 2 mM glutamine (Jinuo) and 1
mM sodium pyruvate (Jinuo) at 37°C in a 5% CO2
atmosphere. EMT induction by TGF-β1 (PeproTech, Inc., Rocky Hill,
NJ, USA) was performed according to a previous protocol (7). Briefly, when cells grew to ~80%
confluence, Panc-1 cells were trypsinized (Jinuo) and seeded into
6-well plates in duplicate (4×105 cells/well). Following
24–48 h, EMT-inducing medium (serum free, containing 1 ng/ml TGF-β1
and 50 ng/ml epidermal growth factor) was used to replace the
common medium of Panc-1 cells, and the cells were incubated for
additional 24–48 h. Next, the optical density was determined.
Western blot analysis
EMT-induced Panc-1 cells were washed twice using
ice-cold PBS and lysed on ice using radioimmunoprecipitation assay
buffer (20 mM Tris-HCl, 150 mM NaCl, 1% NP-40, 0.1% SDS, 0.5%
sodium deoxycholate, 2 mM sodium fluoride, 2 mM
Na3VO4·2H2O, 1 mM EDTA and
protease inhibitor phenylmethane sulfonyl fluoride) (Beyotime
Institute of Biotechnology, Haimen, China). The protein
concentration of the lysates was determined using a BCA Protein
Assay kit (P0009; Beyotime Institute of Biotechnology), and then
marker proteins of EMT were detected using 10% SDS-PAGE and western
blotting. Cell lysates were heated at 100°C for 10 min and
electrophoresed though 10% SDS-PAGE. Then, proteins were
transferred to a polyvinylidene difluoride membrane (EMD Millipore,
Billerica, MA, USA), which were next incubated with 5% bovine serum
albumin (BSA) (ST023; Beyotime Institute of Biotechnology) for 1 h
at room temperature. Subsequently, membranes were incubated with
the corresponding antibodies in a 5% BSA solution at 4°C overnight.
Antibodies against N-cadherin (14215; 1:1,000 dilution), E-cadherin
(5296; 1:1,000 dilution) and vimentin (3390; 1:1,000 dilution)
(8) were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Upon washing three
times with PBS containing Tween 20 [0.1% (v/v) Tween 20, 137 mM
NaCl, 2.7 mM KCl, 10 mM Na2HPO4 and 1.8 mM
KH2PO4), the membranes were incubated with
horseradish peroxidase-conjugated rabbit anti-mouse immunoglobulin
G (58802; Cell Signaling Technology, Inc.; 1:1,000 dilution) for 2
h at room temperature. Finally, protein bands were detected with an
enhanced chemiluminescence detection kit (P0018A; Beyotime
Institute of Biotechnology).
Mitochondrial density of EMT-Panc-1
cells determined by MitoTracker Green FM staining
Panc-1 cells were seeded in 6-well plates and
cultured in EMT-inducing medium. After 24–48 h, Panc-1 cells in
plates were stained with MitoTracker Green FM kit (Beyotime
Institute of Biotechnology) according to the manufacturer's
protocol to determine mitochondria density (9). Briefly, cells were firstly washed using
PBS twice and then incubated at 37°C with 50 nM MitoTracker Green
FM probe for 30 min. Next, the staining buffer was removed and
replaced with fresh complete medium. Fluorescence was detected with
a fluorescent microscope (Olympus Corporation, Tokyo, Japan), and
the cells were detached for analysis using flow cytometry at 490
nm.
Genome DNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis of mtDNA
The mtDNA copy number of EMT-Panc-1 cells was
determined by a standard protocol (10). Total genomic DNA was isolated with the
Gentra Puregene Cell kit (Qiagen GmbH, Hilden, Germany) according
to manufacturer's protocol. The mtDNA content was determined in
cells utilizing RT-qPCR via SYBR Green assay (RR820A; Takara
Biotechnology Co., Ltd., Dalian, China). The method for mtDNA copy
number detection based on qPCR used a 107-bp sized amplicon of
mtDNA transfer RNA (tRNA)Leu (UUR) (forward primer,
5′-CACCCAAGAACAGGGTTTGT-3′ and reverse primer,
5′-TGGCCATGGGTATGTTGTTA-3′) to determine the mtDNA copy number, and
an 86-bp sized amplicon of β2-microglobulin (forward primer,
5′-TGCTGTCTCCATGTTTGATGTATCT-3′ and reverse primer,
5′-TCTCTGCTCCCCACCTCTAAGT-3′) to determine nuclear DNA (nDNA) as an
internal control of the experiment (10). The PCR process consisted of 1 cycle at
95°C for 10 min followed by 40 cycles (95°C for 15 sec and 62°C for
30 sec) and melting curve (50–95°C with a 0.5°C interval). qPCR was
conducted in a StepOnePlus Real-Time PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc., Waltham, MA, USA). PCR
assays were performed in triplicate for each DNA sample. The
expression of mtDNA copy number relative to that of nDNA was
determined using the formula 2×2ΔCq, where ΔCq is the
difference of the Cq values between the β2-microglobulin gene and
the tRNALeu (UUR) gene.
Detection of ROS by flow cytometry and
fluorescence spectroscopy
Panc-1 cells were pretreated with EMT-inducing
buffer for 24–48 h and stained with the ROS detection probe
2′,7′-dichlorofluorescin diacetate at a final concentration of 10
µM for 20–30 min at 37°C. The level of ROS was detected using
fluorescence spectroscopy. Subsequently, the cells were harvested
and washed twice with PBS, resuspended in PBS, and analyzed by flow
cytometry.
ΔΨm determination
ΔΨm determination of EMT-Panc-1 cells was conducted
using the lipophilic cationic fluorescent dye
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolcarbocyanine
iodide (JC-1) (Beyotime Institute of Biotechnology). The cells were
plated and treated according to the manufacturer's protocol, and 1
µM JC-1 was added 30 min prior to harvesting the cells.
JC-1-stained cells were firstly counted using fluorescence
spectroscopy (Leica Microsystems GmbH, Wetzlar, Germany), and then
the cells were collected by trypsinization and washed with PBS. The
red (aggregated JC-1; R2 region) and green (monomeric JC-1; R1
region) fluorescence signals were analyzed by flow cytometry.
Statistical analysis
Data were analyzed with SPSS 19.0 software (IBM
SPSS, Armonk, NY, USA) and expressed as means ± standard deviation.
The statistical significance of differences was evaluated using an
unpaired, non-parametric Student's t test. P<0.05 was considered
to indicate a statistically significant difference.
Results
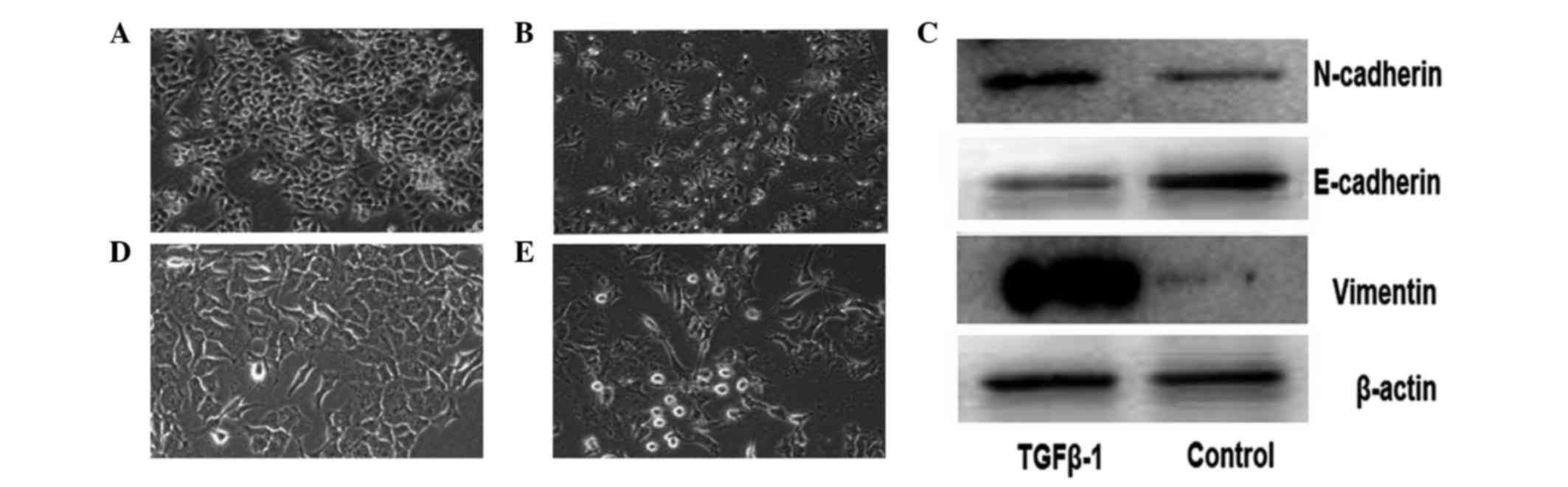
TGFβ-1 treatment induces morphological
changes and a shift from epithelial to mesenchymal phenotype in
pancreatic cancer
For EMT induction in cancer cells, TGFβ-1, which is
a major factor during EMT (11), was
used. As demonstrated by Ikenaga et al (11), pancreatic cancer cells treated with
TGFβ-1 exhibited a spindle-shaped fibroblastic morphology and cell
scattering compared with untreated cancer cells (Fig. 1). Lower E-cadherin expression, and
higher N-cadherin and vimentin expression, were detected in Panc-1
cells following treatment with TGFβ-1 by western blotting (Fig. 1C). These results indicate that the
pancreatic cancer Panc-1 cells acquired a mesenchymal phenotype,
suggesting that EMT was induced by TGFβ-1 in these cells.
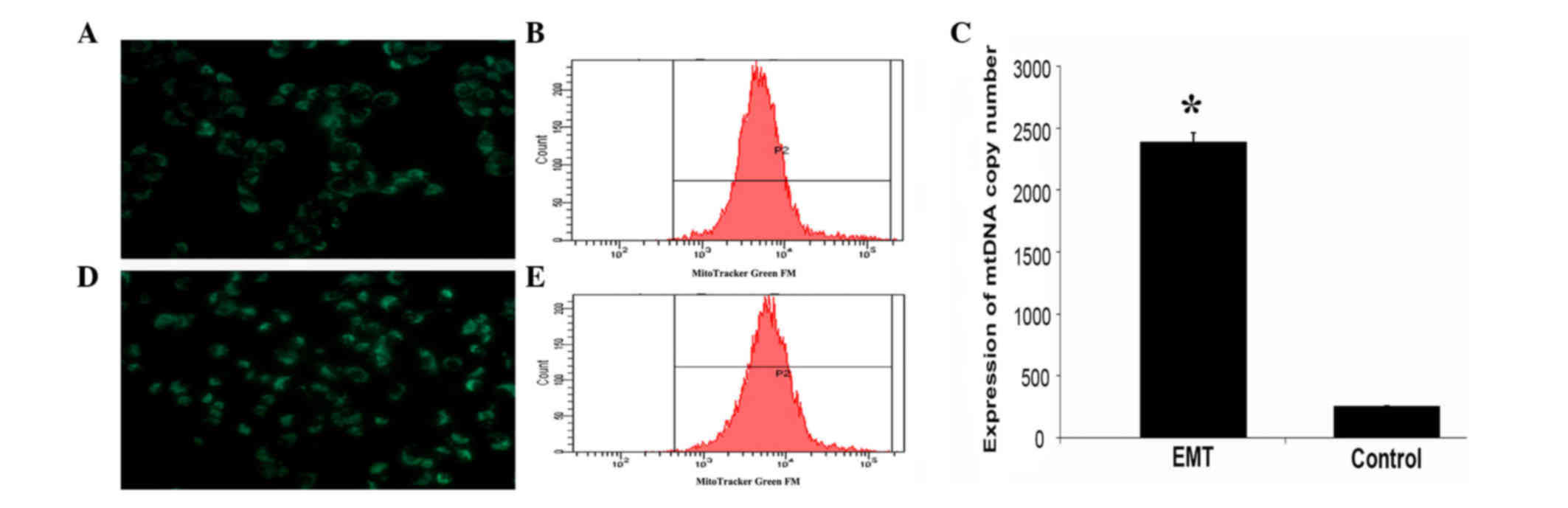
Increased mitochondrial mass following
TGFβ-1 treatment in pancreatic cancer
To examine the mitochondrial mass upon TGFβ-1
treatment, pancreatic cancer cells were stained with MitoTracker
Green FM dye. As shown in Fig. 2,
there were significant differences in mitochondrial mass between
treated and control (untreated) cells (P=0.0004). Cells that
underwent EMT had more mitochondria than control cells. To
determine the mitochondrial mass more precisely, the fluorescence
intensity of the cells was quantified immediately by flow
cytometric scanning. The results of flow cytometry revealed that
the mitochondrial mass of the treated group was higher than that of
the control cells (Fig. 2B and E). In
addition, the mtDNA content was also significantly increased in
treated cells relative to that in control cells (P=0.0005)
(Fig. 2C).
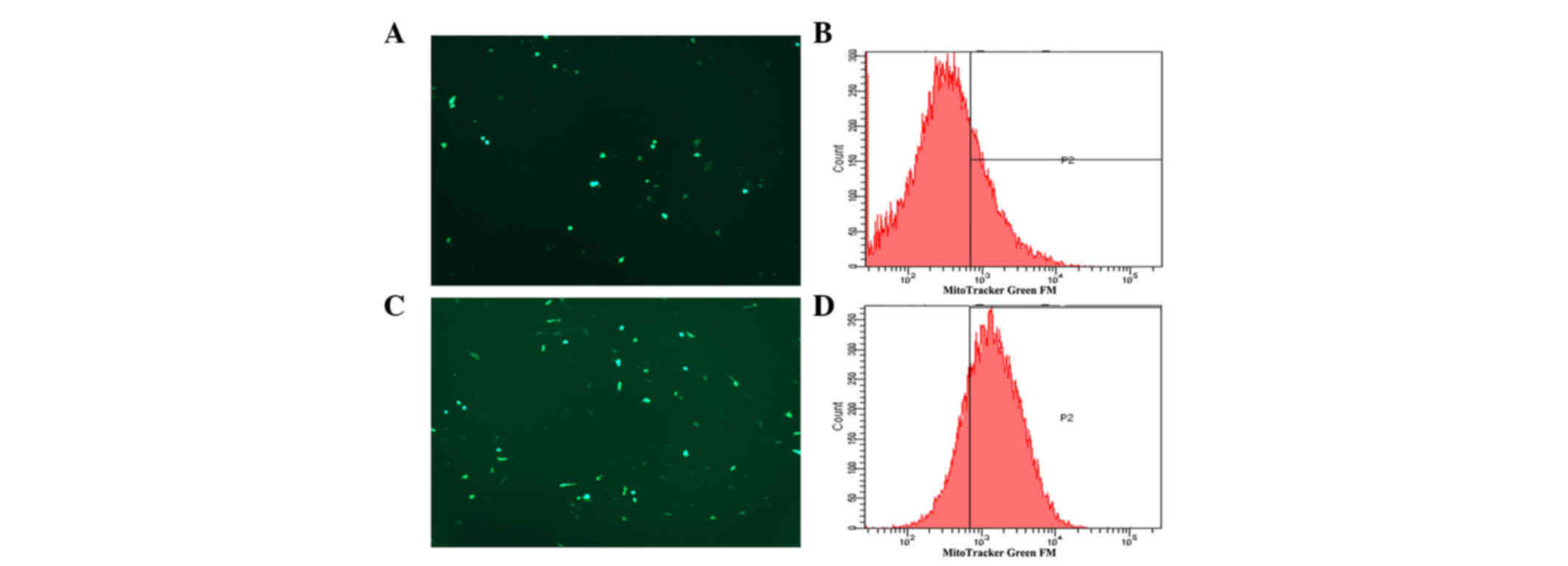
Effect of TGFβ-1 treatment on the
production of ROS in pancreatic cancer cells
Mitochondria are considered the main source of ROS,
and ROS production is generally associated with impairments of the
respiratory chain and mitochondrial function (12,13).
Therefore, in order to ascertain whether alterations in the
generation of ROS also occur in association with EMT, the
intracellular ROS levels were measured in pancreatic cancer cells
with or without TGFβ-1 treatment. The results are shown in Fig. 3. The production of ROS in
TGFβ-1-treated tumor cells was significantly increased compared
with that in control cells (P=0.004).
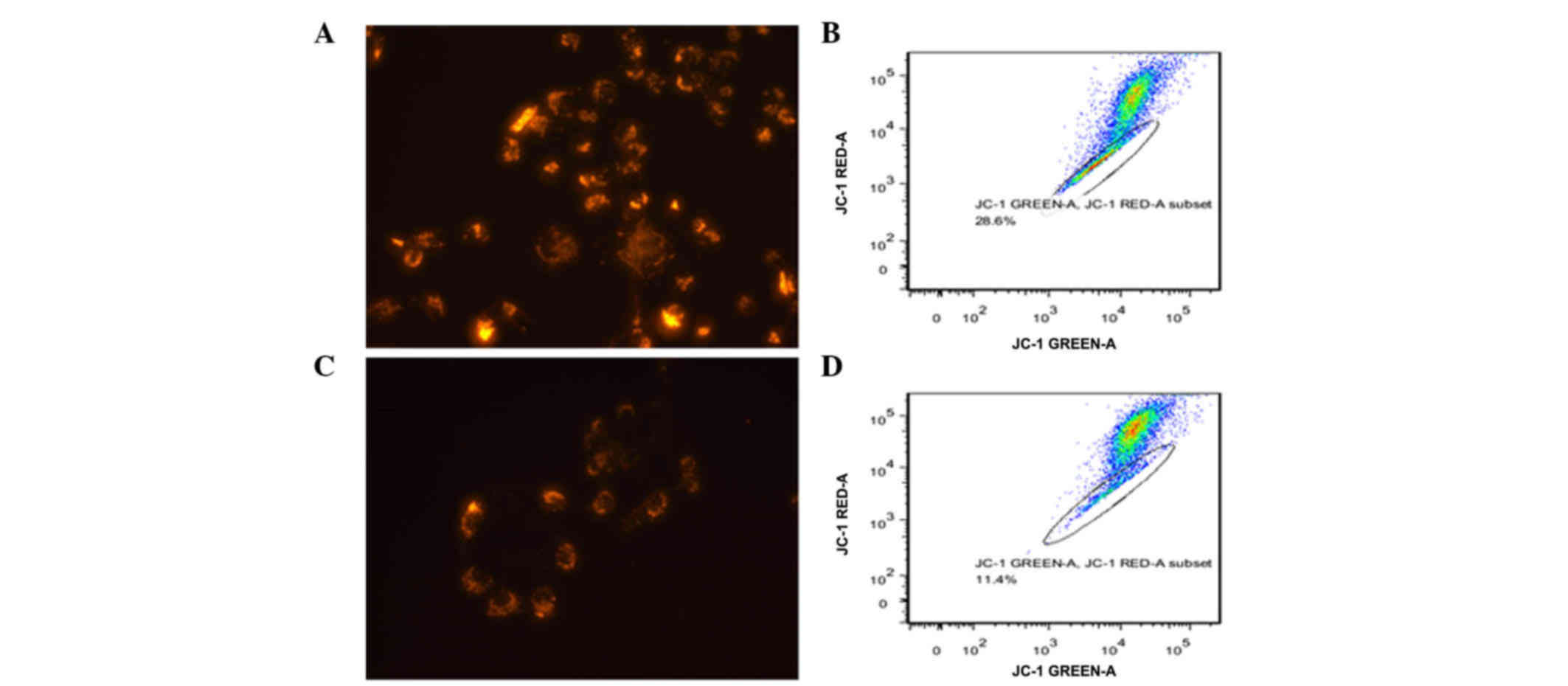
Decreased ΔΨm upon TGFβ-1 treatment in
pancreatic cancer
The present study further investigated whether
TGFβ-1 treatment affected the ΔΨm, since this is important for
mitochondrial functions (14). A
fluorescent probe, JC-1, was used to stain polarized mitochondria.
As represented in Fig. 4A and C,
incubation with TGFβ-1 decreased the number of pancreatic cancer
cells displaying a high ΔΨm. Flow cytometry revealed the same
results than fluorescence microscopy (Fig. 4B and D).
Discussion
The EMT is a basic physiological process in which
epithelial cells lose their polarity and undergo a transition to a
mesenchymal phenotype (6). Hallmarks
of EMT include loss of cell-cell adhesion, re-organization of
cytoskeletal actin and acquisition of migratory characteristics
(15).
Emerging evidence suggests that EMT is essential in
promoting tumor invasion, metastasis, recurrence and drug
resistance in various types of cancer, including pancreatic cancer
(16,17). TGFβ-1 promotes EMT by transcriptional
and post-transcriptional regulation of a group of transcription
factors that suppress epithelial features and enhance mesenchymal
features (18,19). In agreement with previous reports
(20,21), the present study demonstrated that
TGFβ-1 induces EMT in pancreatic cancer cells by acquisition of
mesenchymal morphology, increased expression of the mesenchymal
markers vimentin and N-cadherin, and decreased expression of the
epithelial marker E-cadherin.
Mitochondria are the primary energy producers of the
cell, and regulate intracellular energy metabolism, cell death and
free radical (ROS) production (22).
In this context, the aim of the present study is to demonstrate the
change in mitochondrial function during EMT induced by TGFβ-1 in
pancreatic cancer. First, mitochondrial mass and mtDNA were
investigated. Human mitochondria contain a small quantity of their
own DNA (mtDNA), which is essential for normal mitochondrial
function (23). Changes in mtDNA
content have been described in a wide variety of cancers, with both
increases and decreases being reported in either tumor tissue,
circulating cells or metastatic cancer cells (24,25).
However, in the present study, the mitochondrial mass and mtDNA
content were significantly increased in TGFβ-1-treated pancreatic
cancer cells relative to those detected in the control cells.
Furthermore, a recent study revealed that high mtDNA content is
associated with tumor invasion and EMT characteristics in
esophageal squamous cell carcinoma (ESCC) cells, and suggested that
a relatively high mtDNA copy number may confer an advantage for
tumor invasion in ESCC (26). Other
studies have associated mitochondrial dysfunction with mtDNA copy
number, and have proposed that the mtDNA content may be a potential
biomarker of mitochondrial dysfunction (23,27).
Therefore, the increase in mitochondrial mass and mtDNA in
pancreatic cancer cells with EMT phenotype observed in the present
study suggests that mitochondrial dysfunction happened during the
process of EMT, and may promote pancreatic cancer cell
migration.
Mitochondrial dysfunction has been regarded as a
hallmark of malignancy in human gastric cancer (28). Several studies have implied that
mitochondrial dysfunction is important in cancer metastasis, in
which induction of ROS was a key element. Thus, increased
generation of ROS has an association with mitochondrial damage and
dysfunction (27–29). Additionally, accumulating evidence
suggests that cancer cells exhibit increased intrinsic ROS stress
partly due to mitochondrial malfunction (30). In turn, excessive production of ROS in
cancer cells may contribute to mitochondrial dysfunction and
further lead to the stimulation of cellular proliferation, cell
migration and invasion, thus contributing to carcinogenesis
(31). Therefore, the current study
next examined whether EMT induced by TGFβ-1 resulted in excessive
production of ROS in pancreatic cancer cells (32). As expected, an increase in ROS was
observed in TGFβ-1-treated cancer cells. This is consistent with
the results of Zhou et al (33), who noticed that hypoxia-induced EMT
requires the generation of mitochondrial ROS, which participate in
hypoxia-induced TGF-β1 production and results in EMT. Furthermore,
a previous study reported that EMT-promoted mitochondrial
dysfunction was accompanied by increased ROS generation and
decreased ΔΨm (27). Thus,
determination of ΔΨm following TGF-β1 treatment was conducted in
the present study. As previously reported, the ΔΨm was markedly
increased in the present study, suggesting that mitochondrial
dysfunction occurred during the process of EMT (27).
In conclusion, the present study investigated the
change in mitochondrial function that occurs during EMT induced by
TGFβ-1 exposure in pancreatic cancer. Our results indicate that the
phenomenon of EMT is associated with mitochondrial dysfunction.
Mitochondrial dysfunction may be a cause of EMT in pancreatic
cancer, which leads to heterogeneity in pancreatic cancer, and may
be a potential therapeutic target in the future.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (Beijing, China; grant
no. 81071960) and the New Teacher Foundation of Ministry of
Education (Beijing, China; grant no. 20100101120129).
References
|
1
|
Kim AJ, Jee HJ, Song N, Kim M, Jeong SY
and Yun J: p21 (WAF1/C1P1)
deficiency induces mitochondrial dysfunction in HCT116 colon cancer
cells. Biochem Biophys Res Commun. 430:653–658. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kang D and Hamasaki N: Alterations of
mitochondrial DNA in common diseases and disease states: Aging,
neurodegeneration, heart failure, diabetes, and cancer. Curr Med
Chem. 12:429–441. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jose C and Rossignol R: Rationale for
mitochondria-targeting strategies in cancer bioenergetic therapies.
Int J Biochem Cell Biol. 45:123–129. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gottfried E, Kreutz M and Mackensen A:
Tumor metabolism as modulator of immune response and tumor
progression. Semin Cancer Biol. 22:335–341. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yilmaz M and Christofori G: EMT, the
cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev.
28:15–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pirozzi G, Tirino V, Camerlingo R, Franco
R, La Rocca A, Liguori E, Martucci N, Paino F, Normanno N and Rocco
G: Epithelial to mesenchymal transition by TGFβ-1 induction
increases stemness characteristics in primary non small cell lung
cancer cell line. PLoS One. 6:e215482011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moreno-Bueno G, Peinado H, Molina P,
Olmeda D, Cubillo E, Santos V, Palacios J, Portillo F and Cano A:
The morphological and molecular features of the
epithelial-to-mesenchymal transition. Nat Protoc. 4:1591–1613.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fetisova EK, Avetisyan AV, Izyumov DS,
Korotetskaya MV, Chernyak BV and Skulachev VP:
Mitochondria-targeted antioxidant SkQR1 selectively protects MDR
(Pgp 170)-negative cells against oxidative stress. FEBS Lett.
584:562–566. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Venegas V, Wang J, Dimmock D and Wong LJ:
Real-time quantitative PCR analysis of mitochondrial DNA content.
Curr Protoc Hum Genet Chapter. 19:Unit 19.7. 2011. View Article : Google Scholar
|
|
11
|
Ikenaga N, Ohuchida K, Mizumoto K, Akagawa
S, Fujiwara K, Eguchi D, Kozono S, Ohtsuka T, Takahata S and Tanaka
M: Pancreatic cancer cells enhance the ability of collagen
internalization during epithelial-mesenchymal transition. PLoS One.
7:e404342012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rigoulet M, Yoboue ED and Devin A:
Mitochondrial ROS generation and its regulation: Mechanisms
involved in H(2)O(2) signaling. Antioxid Redox Signal. 14:459–468.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Figueira TR, Barros MH, Camargo AA,
Castilho RF, Ferreira JC, Kowaltowski AJ, Sluse FE, Souza-Pinto NC
and Vercesi AE: Mitochondria as a source of reactive oxygen and
nitrogen species: From molecular mechanisms to human health.
Antioxid Redox Signal. 18:2029–2074. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brand MD and Nicholls DG: Assessing
mitochondrial dysfunction in cells. Biochem J. 435:297–312. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bao B, Wang Z, Ali S, Kong D, Li Y, Ahmad
A, Banerjee S, Azmi AS, Miele L and Sarkar FH: Notch-1 induces
epithelial-mesenchymal transition consistent with cancer stem cell
phenotype in pancreatic cancer cells. Cancer Lett. 307:26–36. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Z, Li Y, Ahmad A, Azmi AS, Kong D,
Banerjee S and Sarkar FH: Targeting miRNAs involved in cancer stem
cell and EMT regulation: An emerging concept in overcoming drug
resistance. Drug Resist Updat. 13:109–118. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Heldin CH, Vanlandewijck M and Moustakas
A: Regulation of EMT by TGFβ in cancer. FEBS Lett. 586:1959–1970.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Horiguchi K, Sakamoto K, Koinuma D, Semba
K, Inoue A, Inoue S, Fujii H, Yamaguchi A, Miyazawa K, Miyazono K
and Saitoh M: TGF-β drives epithelial-mesenchymal transition
through δEF1-mediated downregulation of ESRP. Oncogene.
31:3190–3201. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Boreddy SR and Srivastava SK: Deguelin
suppresses pancreatic tumor growth and metastasis by inhibiting
epithelial-to-mesenchymal transition in an orthotopic model.
Oncogene. 32:3980–3991. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lou C, Zhang F, Yang M, Zhao J, Zeng W,
Fang X, Zhang Y, Zhang C and Liang W: Naringenin decreases
invasiveness and metastasis by inhibiting TGF-β-induced epithelial
to mesenchymal transition in pancreatic cancer cells. PLoS One.
7:e509562012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen EI: Mitochondrial dysfunction and
cancer metastasis. J Bioenerg Biomembr. 44:619–622. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Malik AN and Czajka A: Is mitochondrial
DNA content a potential biomarker of mitochondrial dysfunction?
Mitochondrion. 13:481–492. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu M: Generation, function and diagnostic
value of mitochondrial DNA copy number alterations in human
cancers. Life Sci. 89:65–71. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sotgia F, Whitaker-Menezes D,
Martinez-Outschoorn UE, Flomenberg N, Birbe RC, Witkiewicz AK,
Howell A, Philp NJ, Pestell RG and Lisanti MP: Mitochondrial
metabolism in cancer metastasis: Visualizing tumor cell
mitochondria and the ‘reverse Warburg effect’ in positive lymph
node tissue. Cell Cycle. 11:1445–1454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin CS, Lee HT, Lee SY, Shen YA, Wang LS,
Chen YJ and Wei YH: High mitochondrial DNA copy number and
bioenergetic function are associated with tumor invasion of
esophageal squamous cell carcinoma cell lines. Int J Mol Sci.
13:11228–11246. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yuan Y, Chen Y, Zhang P, Huang S, Zhu C,
Ding G, Liu B, Yang T and Zhang A: Mitochondrial dysfunction
accounts for aldosterone-induced epithelial-to-mesenchymal
transition of renal proximal tubular epithelial cells. Free Radic
Biol Med. 53:30–43. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hung WY, Huang KH, Wu CW, Chi CW, Kao HL,
Li AF, Yin PH and Lee HC: Mitochondrial dysfunction promotes cell
migration via reactive oxygen species-enhanced β5-integrin
expression in human gastric cancer SC-M1 cells. Biochim Biophys
Acta. 1820:1102–1110. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ma J, Zhang Q, Chen S, Fang B, Yang Q,
Chen C, Miele L, Sarkar FH, Xia J and Wang Z: Mitochondrial
dysfunction promotes breast cancer cell migration and invasion
through HIF1α accumulation via increased production of reactive
oxygen species. PLoS One. 8:e694852013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Moreno-Sánchez R, Rodríguez-Enríquez S,
Marín-Hernández A and Saavedra E: Energy metabolism in tumor cells.
FEBS J. 274:1393–1418. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ishikawa K, Takenaga K, Akimoto M,
Koshikawa N, Yamaguchi A, Imanishi H, Nakada K, Honma Y and Hayashi
J: ROS-generating mitochondrial DNA mutations can regulate tumor
cell metastasis. Science. 320:661–664. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bonnard C, Durand A, Peyrol S, Chanseaume
E, Chauvin MA, Morio B, Vidal H and Rieusset J: Mitochondrial
dysfunction results from oxidative stress in the skeletal muscle of
diet-induced insulin-resistant mice. J Clin Invest. 118:789–800.
2008.PubMed/NCBI
|
|
33
|
Zhou G, Dada LA, Wu M, Kelly A, Trejo H,
Zhou Q, Varga J and Sznajder JI: Hypoxia-induced alveolar
epithelial-mesenchymal transition requires mitochondrial ROS and
hypoxia-inducible factor 1. Am J Physiol Lung Cell Mol Physiol.
297:L1120–L1130. 2009. View Article : Google Scholar : PubMed/NCBI
|